
小麦分蘖性状分子遗传研究进展
2022-05-20 08:15王同著郑跃婷杨雨露张香粉
麦类作物学报 2022年5期
陈 悦,王同著,郑跃婷,杨雨露,王 盈,张香粉,陈 锋,赵 磊
(河南农业大学农学院/省部共建小麦玉米作物学国家重点实验室/河南粮食作物协同创新中心,河南郑州 450046)
小麦是世界上种植面积最为广泛的粮食作物,为全球约40%的人口提供食物来源。小麦同时也是中国的第二大粮食作物,是中国人民最主要的口粮之一。小麦生产与国家粮食安全和社会稳定休戚相关,提高小麦产量一直都是科研人员关心的主要问题。在现有耕地面积逐年减少的情况下,培育具有更高单产潜力的小麦新品种就成为进一步提高小麦产量、解决粮食安全的必要举措之一。
株型一般是指植株地上部分的形态特征,是作物诸多重要农艺性状的综合,影响作物的适应性、产量以及收获指数等。合理的株型对作物产量的提高至关重要,株型改良育种在小麦高产育种中发挥着重要作用。20世纪60年代在全球范围内兴起的“绿色革命”正是通过利用矮秆基因实现矮化育种,最终极大地提高了小麦和水稻的产量,成功地解决了当时全球粮食危机。因此,对小麦株型相关性状的遗传机理进行研究,挖掘株型调控基因,并借助分子手段辅助选择育种,将有助于培育出株型合理的高产小麦新品种。
分蘖是小麦、水稻等谷类作物株型构成的主要因素,包括分蘖能力和分蘖角度两个方面,均是与产量相关的重要农艺性状。分蘖能力一般用分蘖数目来表示,是指植株近地面发生的分枝数目,其中能够抽穗并结实的地上部分蘖称为有效分蘖,其决定了作物的有效穗数和光合面积大小,直接影响产量;分蘖角度指分枝与竖直方向的夹角,体现了主茎与分蘖之间的集散程度,决定了植株的空间构型,并通过影响群体的种植密度以及光合效率间接影响作物群体产量。研究表明,通过调控分蘖的发育进程,禾谷类植物能够改变群体结构,最大限度的提高光能利用率,并提高对环境的适应性,进而影响产量。因此,对小麦分蘖相关性状分子机理进行研究,不仅有助于认识小麦株型的遗传特点,同时对株型改良育种也具有重要借鉴意义,有助于培育出优质高产的小麦新品种,为提高中国乃至世界粮食产量以及解决粮食危机提供出路。本文从分子水平对近年来小麦分蘖性状的遗传研究进展进行总结,以期为开展相关研究提供参考。
1 分蘖的形成与调控
分蘖(分枝)是植株地上部形态的主要表现之一。从幼苗期至成熟期,植株对环境变化进行响应并不断生长,由分生组织分裂产生新的细胞,进一步经过分化形成组织和器官。茎尖分生组织(shoot apical meristem,SAM)促进植株在垂直方向的生长;叶腋分生组织(axillary meristem,AM)则可以经过分化形成侧枝或分枝。AM形成于叶原基近轴侧的边界区中心,边界区将SAM与发育中的叶原基分开以维持分生组织和器官发育。分蘖的发育一般经历三个阶段:腋芽分生组织起始阶段、腋芽带叶发育阶段和腋芽长出阶段。虽然腋芽可发育形成许多分蘖,但并非所有分蘖最终都能成为有效穗,有许多为无效分蘖。因此,分蘖的发育和生长在很大程度上影响植物的整体生长状况及生产力。
植物分蘖(分枝)发育及其形成过程受多种植物激素相互作用共同调节。长期以来,一直认为植物分枝由生长素(indole acetic acid,IAA)和细胞分裂素(cytokinin,CK)两种植物激素参与调控。尤其是IAA,一直被视作为一种经典的、占主导地位的分枝调节激素。CK被认为是调节IAA作用的第二信使,其在根中合成,然后被转运至腋芽中,打破芽的休眠状态;同时IAA通过调节CK的浓度介导芽的生长。独脚金内酯(strigolactone,SL)是近些年新发现的第三类植物调控激素,也被认为参与调控植物的分枝发育,但IAA和SL如何相互作用控制腋芽生长仍存有争议。Brewer等研究认为,SL作用于IAA的下游,直接抑制腋芽的生长;而Crawford等研究认为,SL通过调节IAA极性运输来控制腋芽的生长。此外,Ferguson等研究报道,SL抑制芽的生长,而CK促进芽的生长,IAA则通过维持高SL和低CK的含量负调控芽的生长。另外,光照也会对分蘖产生一定的影响。光感受器光敏色素有Pr(非活性)和Pfr(活性)两种不同的构象,当吸收红光后,Pr转化为Pfr,Pfr通过与光敏色素相互作用因子(phytochrome interacting factor,PIF)或PIL(PIF3-like)相互作用,穿梭于细胞核并调控基因表达;低Pr/Pfr光照比可诱导()基因上调表达,从而抑制分枝发生。此外,在植株生长条件下,磷和氮的含量也对分枝起调控作用。研究表明,在磷酸盐饥饿条件下,促进了拟南芥、番茄、高粱和水稻中SL生物合成基因(如和)的表达;在低氮条件下,高粱和豌豆植株中SL的含量也明显增加。对拟南芥IAA响应突变体和SL生物合成突变体的分析表明,在低氮条件下,完整的IAA信号转导和SL生物合成途径可以增加茎尖生长素的供应,进而抑制侧枝生长。
2 分蘖性状相关基因/QTL的定位研究
分蘖是复杂的发育性状,其形成受基因型、植物激素、环境以及营养条件等诸多因素的影响,但更容易受到环境的影响,同时又具有较强的遗传可塑性,通常认为分蘖是由多基因共同调控的数量性状。以下分别围绕小麦分蘖数目和分蘖角度对现有主要QTL定位研究进行简要 综述。
2.1 分蘖数目相关基因/QTL的定位研究
对小麦分蘖数目的遗传定位研究最早主要是从分蘖抑制基因开始的。是最早在普通小麦中鉴定到的分蘖抑制基因,该基因为隐性基因,与颖壳稃毛和黑芒基因位点连锁,定位在1AS染色体上,与微卫星标记Xgwm136紧密连锁。位于2A染色体上的基因是一个显性基因,同样具有抑制分蘖的效应。Kuraparthy等利用栽培一粒小麦单分蘖突变体在3AL染色体上鉴定到由单基因隐性突变引起的分蘖抑制基因,与标记Xpsr1205紧密连锁,该基因的突变体植株只有一个主茎。Zhang等利用寡分蘖材料Pubing 3558与京4841杂交,在1AS染色体上鉴定到一个分蘖抑制基因,该基因与SSR标记Xcfa2153紧密连锁,并推测是一个不同于的新位点。Wang等利用一个寡分蘖材料H461与川农16构建的RIL群体,定位到三个与低分蘖数目相关的QTL,分别为、和,可分别解释达19.1%、14.6%和9.6 %的表型变异(表1)。
除鉴定到减少分蘖数目的QTL外,也鉴定到的增加分蘖数目的QTL。如Kato等利用染色体代换系中国春(Cappelle Desprez 5A)在5A染色体上定位到一个控制分蘖数目的QTL,该位点同春化基因相关联,可解释 37.0%的表型变异。Li等用普通小麦Opata85和人工合成的六倍体小麦W-7984构建的RIL群体为材料,分别在1Ds、2Ds和6As染色体上鉴定到三个与单株分蘖数目相关的QTL,可解释 11.0%~31.0%的表型变异。Campbell等和Dilbirligi等利用染色体重组代换系CNN(WI3A)在3A染色体上共鉴定到四个与单位面积穗数相关的QTL,可解释9.0%~17.0%的表型变异。Narasimhamoorthy等以硬红冬小麦品种卡尔92为轮回亲本,以TA4152-4为供体亲本,构建BCF回交群体,在3B染色体检测到一个与分蘖数目相关的QTL(),可解释6.2%的表型变异。Li等利用花培3和豫麦57构建的DH群体和永久F群体检测到多个与最大分蘖数和有效分蘖数相关的QTL,可解释4.5%~35.0%的表型变异。Zou等利用两个春小麦品种Attila和CDC Go构建的RIL群体为材料,在6A染色体上70 cM处检测到一个与每平方米分蘖数相关的QTL(),可解释11.2%的表型变异。Hu等利用川农18和T1208构建的RIL群体为材料,在4D染色体上定位到一个稳定的穗形成率主效QTL(),可解释18.2%~24.5%的表型变异。Wang等用UI Platinum和BSY Capstone构建的DH群体为材料,定位到两个与单位面积分蘖数相关的QTL(和),分别可解释19.0%和26.0%的表型变异。Ren等利用55K芯片对川农18和T1208构建的RIL群体进行分析,在多个环境下共检测到7个稳定的QTL,分布于2D、4A、4D、5A、5D和7D染色体上,单个QTL可解释 1.6%~21.2%的表型变异。Liu等以20828和SY95-71构建的RIL群体为材料,在1BL染色体上检测到一个控制有效分蘖数的主效遗传位点,可解释12.1%~55.7%的表型变异。此外,前人也分别在1B、3A、3D、4B、5B、6B、6D、7A等染色体上均定位到与分蘖数目相关的QTL。上述研究结果表明,几乎在所有染色体上都能定位到与分蘖数目相关的QTL(汇总结果见表1),而且不同研究之间的结果不一致,在QTL数量、染色体分布以及QTL效应均有不同,这也进一步表明分蘖性状遗传的复杂性。而且截至目前,上述QTL仍停留在初步定位阶段,没有一个被克隆出来,研究任务仍然任重而道远。
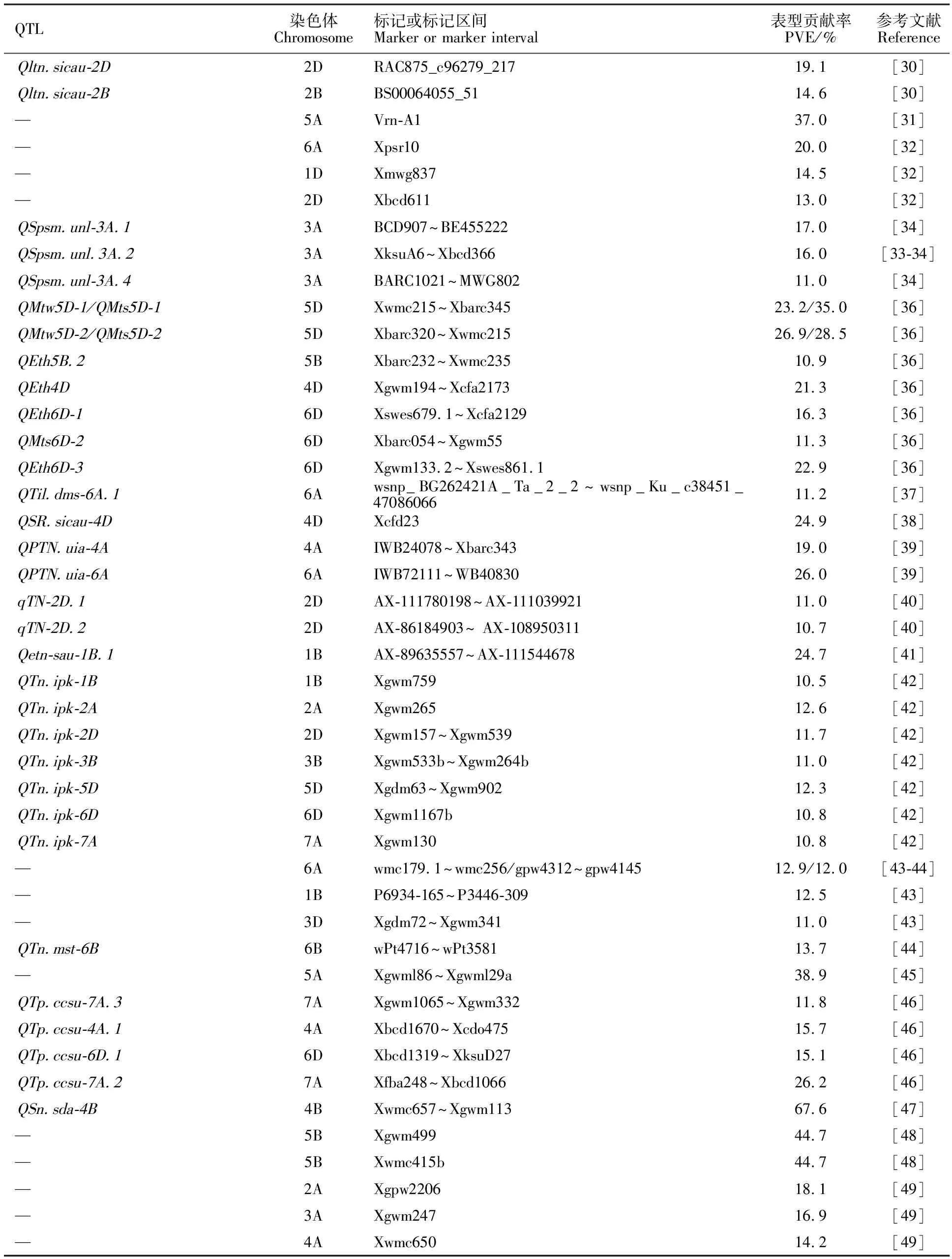
表1 平均表型贡献率大于10%的分蘖数目主效QTL
2.2 分蘖角度相关基因/QTL的定位研究
与分蘖数目相比,前人对小麦中分蘖角度的遗传研究较少。究其原因,可能由于该性状更易受环境影响,难以进行准确测量,且长期以来不受重视所致,近几年来相关研究逐渐增多。Marone等对184份硬粒小麦的“匍匐/直立”生长习性进行全基因组关联分析(genome wide association study,GWAS),共鉴定到12个调控分蘖角度的QTL,其中位于2B和5A染色体上的两个稳定位点均可在6个环境中检测到。Zhao等以中麦871和中麦895为亲本构建的RIL群体为材料,在1AL和5DL染色体上检测到两个与分蘖角度相关的稳定QTL,并进一步预测了5DL染色体位点上的候选基因,依据水稻中的同源基因将其命名为,并对其功能进行初步分析,但未进行转基因功能验证。Liu等以3个RIL群体和1个杂交F群体为材料,在2BL染色体上检测到一个调控分蘖角度的主效位点,可解释18.1%~51.1%的表型变异;该位点距离较近,但独立于,并通过分析预测了候选基因,但也未对候选基因功能进行验证。
3 小麦分蘖性状相关基因/QTL的功能研究
由于普通栽培小麦复杂的遗传背景,在2018年之前一直未获得完整的基因组序列,采用图位克隆的方法从六倍体小麦中分离基因非常困难;此外,分蘖性状易受环境因素的影响,因此,迄今为止,利用正向遗传手段克隆小麦分蘖性状相关基因仍罕有报道。
参考拟南芥、水稻等植物中的研究结果,在小麦中采用同源克隆的方法已克隆出数个与分蘖数目相关的基因。如基因是SPL(squamosa promoter binding protein-like)基因家族的成员之一,编码一个含SBP-box结构域的蛋白,受miR156的调控,参与调控理想株型,对小麦的分蘖、株高、茎粗和穗部发育均起着重要的调控作用;是植物GRAS家族调控AM起始和分枝的基因,其编码的蛋白具有保守功能域VHIID,与水稻中的MOC1、拟南芥中的LAS和番茄中的LS具有较高的同源性,参与小麦分蘖的形成和穗的发育;其启动子上存在一个顺式作用元件CAT-box,决定了TaMOC1的表达部位,并影响启动子的活性,但具体如何发挥作用调控启动子的活性尚不清楚;基因同水稻中的同源基因具有相似的功能,通过参与SL合成途径进而调控小麦分蘖的发生;基因可通过参与SL信号转导途径来调控小麦分蘖数目相关基因的表达,且基因与和基因互作,共同调控小麦分蘖的发生;硬粒小麦中基因除参与调控小麦分蘖的发生外,可能还参与根系的分枝发育。此外,Liu等研究显示,过表达可增加小麦分蘖数目,SL信号抑制因子TaD53可直接与miR156控制的TaSPL3/17的N端结构域相结合,通过多个基因相互作用,最终影响小麦分蘖的形成。研究还发现,AP2类多效基因和可能参与调控小麦分蘖数目,基因可能参与调控小麦分蘖的形成。在分蘖角度调控基因研究上,曹 鑫等利用同源克隆的方法分离获得小麦分蘖角度调控基因,通过分析认为该基因可调控小麦的分蘖角度、分蘖数、旗叶夹角等性状,同时推测该基因可能参与生长素极性运输。另外,Marone等利用GWAS的方法鉴定出多个小麦分蘖角度调控位点,并推测小麦春化()、矮秆()和光周期()相关基因可能也参与调控小麦分蘖角度的形成。此外,Cu等研究发现,调控分蘖数目的基因也可能参与调控小麦分蘖角度。
上述研究结果进一步表明,小麦分蘖相关性状是受多基因控制的数量性状。对小麦分蘖调控基因同源基因的研究也表明,该类基因在不同植物中的功能高度保守,对其他植物中相关重要基因的研究也可促进和加快对小麦分蘖分子遗传机制研究的进程。不足之处是目前对重要基因的研究还比较匮乏,且比较分散,根据现有研究结果还未能获得具体的、可信的调控分蘖相关性状的作用机制途径。
4 讨论与展望
高产是作物研究永恒不变的追求目标,而株型改良育种一直以来都是作物高产育种的主要方向。分蘖模式是植物株型建成的重要构成因素,在过去的一二十年中,通过对模式植物拟南芥以及水稻、玉米、番茄、大豆等农作物的研究,加深了我们对植物分蘖(分枝)遗传机制的理解。在大多数作物中,驯化品种的分蘖要比野生品种少,使其更适于密植,且能减少群体间对光和土壤养分的竞争,同时还可减轻因分蘖过多造成郁闭而使病虫害危害加重;然而分蘖过少会造成穗数不足,也会影响作物产量。分蘖作为重要的农艺性状,与穗数的形成直接相关,是决定作物最终产量的重要因素之一。因此,对分蘖性状遗传机理进行研究不仅有助于增加对作物理想株型建成机制的认识和理解,也可使通过株型改良培育高产作物品种成为可能。
小麦作为人类三大主要粮食作物之一,在保障全球粮食安全中发挥着不可替代的作用。加快小麦重要性状基因的克隆,利用分子生物学手段改良现有品种,对培育高产、优质、抗逆的小麦新品种具有十分重要的意义。小麦分蘖数目直接影响和决定单位面积穗数,分蘖角度也通过影响种植密度间接影响单位面积穗数,两者对小麦产量的形成均具有重要意义。通过对小麦分蘖性状遗传机制进行研究,发掘小麦分蘖相关基因,克隆在生产上有应用前景的优异功能基因,系统研究基因的功能,并借用分子手段加以利用,对拓宽小麦育种资源,改良小麦株型和产量,培育高产、稳产新品种,确保中国乃至全球粮食安全具有十分重要的意义。然而由于六倍体小麦基因组庞大且重复序列较多,采用传统定位方法从中分离基因非常困难;同时分蘖性状是多基因控制的数量性状,且易受环境因素影响,这些因素均增加了研究难度。目前,对于小麦分蘖性状的遗传研究虽然已开展多年,但一直未有突破性进展,大部分工作停留在定位阶段,相关基因的分离和克隆鲜有报道,遗传调控机制仍很不清晰,远远不能满足实际需要,目前还未有相关研究成果应用于育种工作的报道。随着小麦基因组测序工作的发展,小麦基因组数据库及基因注释信息日益完善,IWGSC已公布最新的中国春参考序列(IWGSC RefSeq v2.1),为小麦功能基因组学的研究奠定了良好基础。同时GWAS、集群分离分析(bulked sample/segregant analysis,BSA)、全外显子组测序(whole exome sequencing,BSA)等多种新兴技术的出现和发展,为鉴定复杂农艺性状遗传调控基因提供了快速有效的途径,相信该领域的研究会有日新月异的进展,未来会有更多小麦分蘖相关基因被分离和克隆,且小麦分蘖性状的分子遗传机制也终将会得到阐明。
猜你喜欢
小猕猴智力画刊(2021年6期)2021-08-05
农民致富之友(2019年9期)2019-05-22
农家科技(2018年2期)2018-05-05
新农业(2017年6期)2017-07-15
养生保健指南(2016年12期)2017-01-06
作文大王·低年级(2016年3期)2016-03-11
新农村(2015年10期)2015-09-28
南方农业·下旬(2014年10期)2014-12-20
湖北农业科学(2014年11期)2014-09-10
中学理科·综合版(2008年3期)2008-03-07
