
育成期高能饲喂下开产与未开产蛋鸡肝脏转录组差异分析
2022-05-18 09:12王星果窦套存胡玉萍李永峰王克华
西北农业学报 2022年1期
曲 亮,王星果,郭 军,窦套存,胡玉萍,马 猛,李永峰,王克华
(江苏省家禽科学研究所,江苏扬州 225125)
蛋鸡的生产性能直接与其养殖效益挂钩,一直是蛋鸡行业的重要研究课题[1],而能量水平对生产性能的影响是其中一个重要的研究方向,也越来越受到人们的重视[2-5],其中包括对育成期蛋鸡进行能量调控,检测其对生产性能的影响。研究表明育成期高能饲喂能明显提高蛋鸡的生产性能,如使用不同能量的饲料饲喂育成蛋鸡的研究中发现,高能饲喂能显著提高体质量、体尺、产蛋率和屠宰性能[6-10]。江苏省家禽科学研究所蛋鸡研究室(本研究室)前期研究了不同能量水平饲料对育成期蛋鸡的影响,发现能量水平越高,蛋鸡体质量越大,开产日龄越早[11]。
肝脏是重要的代谢器官,在鸡等家禽中更是物质代谢特别是脂质代谢的主要器官,对育成期的能量调控肯定起着关键作用[12]。同时,产蛋与肝脏也有十分密切的关系[13]。由于鸡蛋中卵黄的30%由脂质构成,而这些脂质均来源于肝脏。肝脏中合成的大部分脂质以脂蛋白的形式运输出肝脏,经血液循环至卵泡,被卵母细胞吸收储存,形成卵黄[14]。因此,肝脏脂质代谢的快慢影响鸡蛋卵黄形成的速度,从而影响产蛋。
近年来,转录组测序逐渐用于研究各种生物过程[15-16]。已有一些转录组测序在鸡肝脏中的研究报道,如对卢氏绿壳蛋鸡产蛋前期和高峰期的肝脏、雌激素处理蛋鸡的肝脏进行转录组测序研究,均发现基因参与肝脏代谢[17-18],本研究室成员也运用转录组测序发现LNA-122、GH处理的鸡肝脏细胞中若干基因参与代谢过程,尤其是脂质代谢[19-20]。本研究室后期对高能饲喂的育成期蛋鸡进行肝脏转录组测序,发现与低能组相比,有188个基因差异表达,对差异表达基因的功能和信号通路分析发现,脂质代谢相关过程富集的基因较多,且显著富集的信号通路都和脂质代谢相关[21]。然而在高能组中的鸡仍有开产早晚之分,有必要对其进行深入分析。本研究选取高能饲喂育成期蛋鸡,并对其中开产与未开产鸡的肝脏进行转录组测序,分析比较其表达谱,探明在高能饮食状态下肝脏中影响开产的基因,为提高产蛋打下基础。
1 材料与方法
1.1 试验动物饲养
选用江苏省家禽科学研究所(本单位)培育的C3蛋鸡品系,8周龄前常规饲养,满8周龄时选取体质量相近母鸡80只,分为8个重复,每个重复10只。9~18周龄,定量饲喂代谢能水平为12.14 MJ/kg的高能饲粮,18周龄后自由采食标准营养水平的饲粮。整个试验过程参照NRC(1994)标准配制试验日粮,分9~18周龄和19周龄后两个阶段配制,配以粉料饲喂。9~16周龄在育成鸡舍四层阶梯笼内饲养(5只/笼),17周龄起在产蛋鸡舍三层阶梯笼内饲养(1只/笼)。整个试验期间,所有鸡只可以自由饮水,并执行常规光照和免疫等程序。
1.2 RNA-seq高通量测序
以20周龄作为开产早晚的界限,开产鸡均为19周龄开产,未开产鸡均在20周龄末仍未开产,20周龄末每个重复选取1只开产鸡和1只未开产鸡,称体质量后各取最接近平均值的3只,未开产鸡分别为N1、N2、N3,开产鸡分别为R1、R2、R3,屠宰并取出肝脏组织,放入液氮中保存。
使用TRIzol试剂提取肝脏组织总RNA,使用琼脂糖电泳和Agilent 2100检测RNA质量。利用rRNA去除试剂盒去除rRNA,并通过离子打断的方式将RNA打断成200~300 bp的片段。RNA经过反转录形成cDNA,再经PCR富集构建cDNA文库,该文库通过荧光定量检测其大小和浓度,上机至Illumina Nextseq500平台进行 测序。
1.3 序列分析
下机得到原始数据,经过reads数、Q20、Q30等初步统计,经碱基质量分布检测后去除接头,进行质量过滤,得到clean reads和clean data。这些数据使用Tophat2与鸡参考基因组进行比对,统计比对到外显子、内含子和基因间区的reads。比对结果使用RSeQC进行插入片段长度分布、reads重复率等质控分析。
比对到各基因的reads使用HTSeq统计,得到各基因的原始表达量,开产鸡和未开产鸡的表达量经过RPKM标准化后,再使用DESeq分析其差异性,按照表达量倍数差异(fold change)> 2、差异显著性P<0.05筛选出差异表达基因。挑选上调基因和下调基因各2个,使用荧光定量PCR(qRT-PCR)进行表达量验证,以ACTB作为内参,引物信息见表1。对差异表达基因使用超几何分布进行GO功能富集分析和KEGG信号通路富集分析。

表1 qRT-PCR引物Table 1 Primers for real-time qRT-PCR
2 结果与分析
2.1 蛋鸡肝脏的测序质量
未开产与开产蛋鸡的肝脏经过RNA-seq测序,得到转录组原始序列。表2为测序数据的初步分析,可见,6个样品测得的原始序列经质量过滤后,纯净序列仍超过80 000 000个,占原始序列的99%以上,说明低质量的序列少,测序效果较好。开产鸡与未开产鸡的原始序列和纯净序列数量均相当,说明两组测序质量相近。
将各样品的纯净序列与鸡参考基因组比对,由统计比对上的序列(表 3)可见,6个样品能比对到基因组的序列均占纯净序列的80%左右,且绝大部分是基因序列,约90%,外显子序列在基因序列中所占比例也很高,均占80%左右。以上结果表明6个样品测序结果均较好,得到的序列基本为有效的基因序列,可进行后续研究。
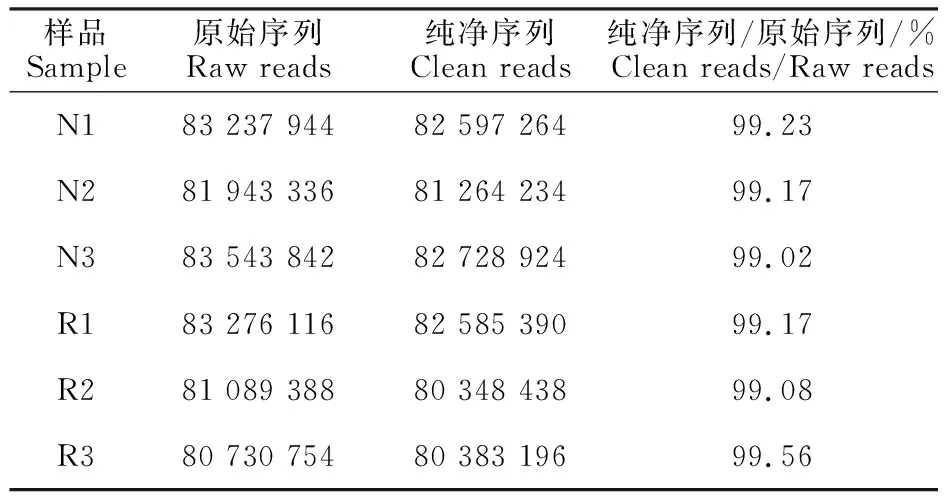
表2 测序数据初步分析Table 2 Preliminary analysis of RNA-seq data
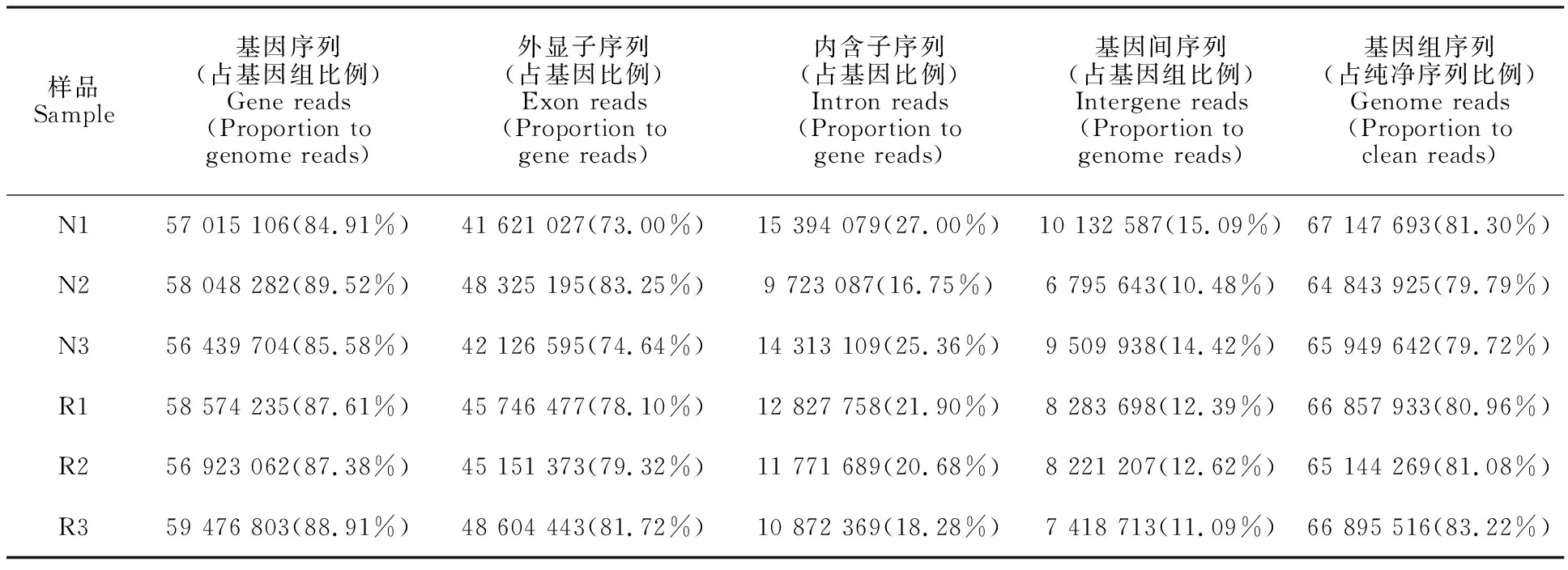
表3 基因鉴定Table 3 Identification of gene
2.2 肝脏基因差异表达分析
对鉴定出的基因进行RPKM表达量分析,并对开产与未开产样品的基因表达量进行比较分析,筛选出差异表达基因(图1),共筛选出494个差异表达基因(P<0.05),相比未开产鸡,开产鸡有285个基因上调,209个基因下调。对差异表达基因进行聚类分析(图2),结果可见,基因大多汇聚在同一大类,说明它们具有相关的功能。表达差异最大的20个基因(10个上调基因,10个下调基因)见表4,可以看出,未开产鸡有7个基因未检出表达,开产鸡有5个基因未检出表达,上调和下调差异为10~3 000倍,提示它们可能对开产性状具有特殊的重要作用。
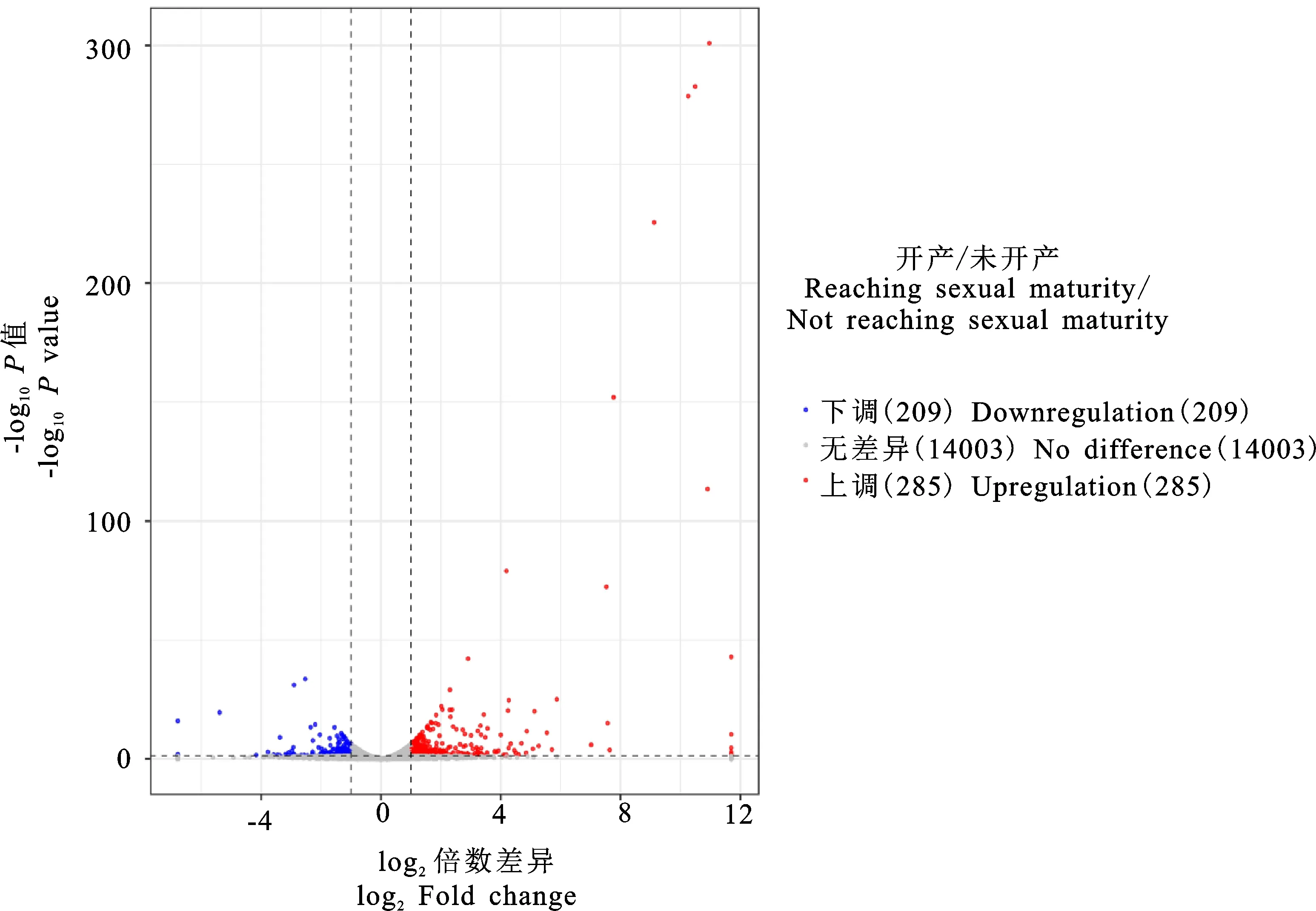
图1 差异表达基因火山图Fig.1 Volcano map of differentially expressed genes
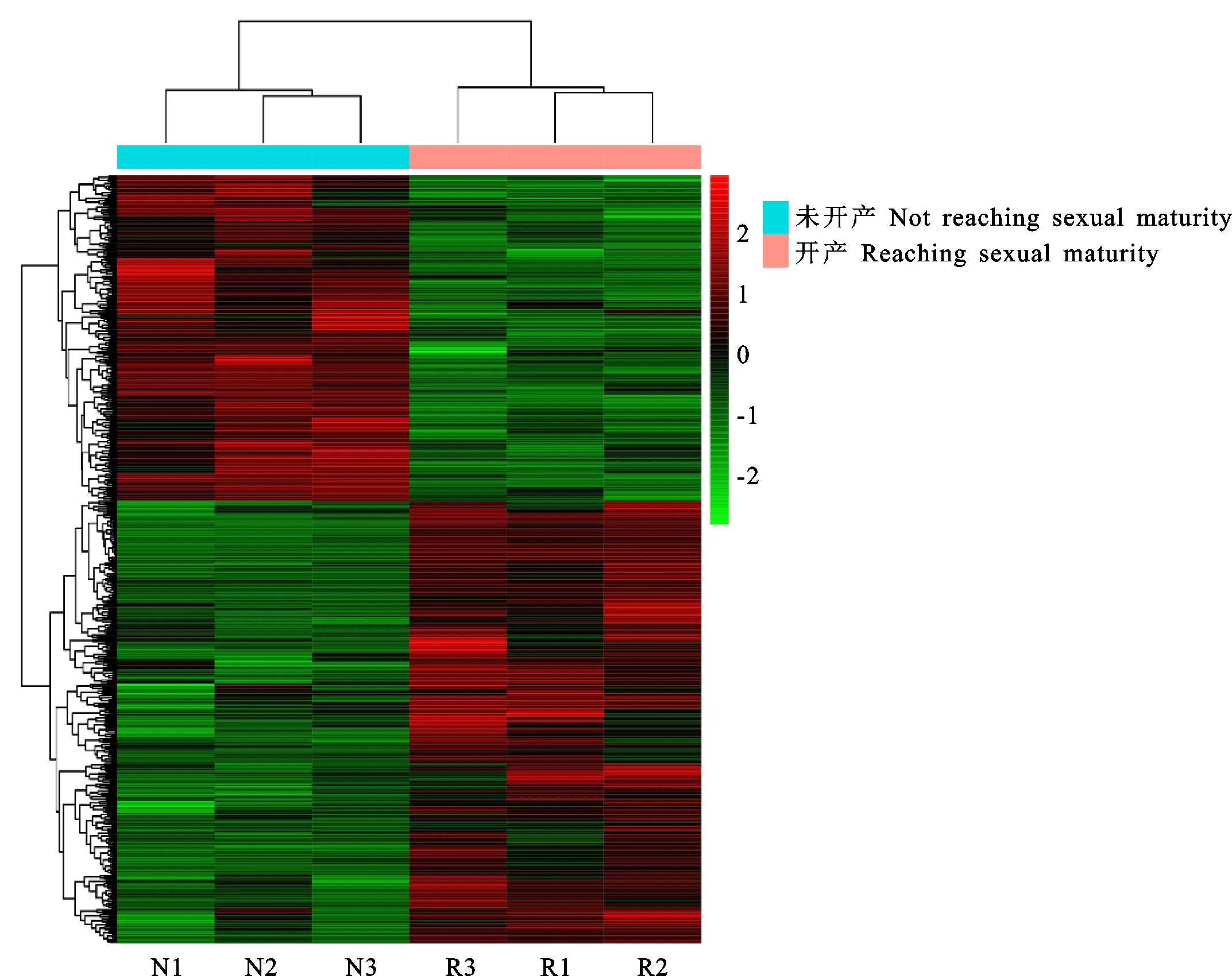
红色为高表达,绿色为低表达 Red means high expression,green means low expression
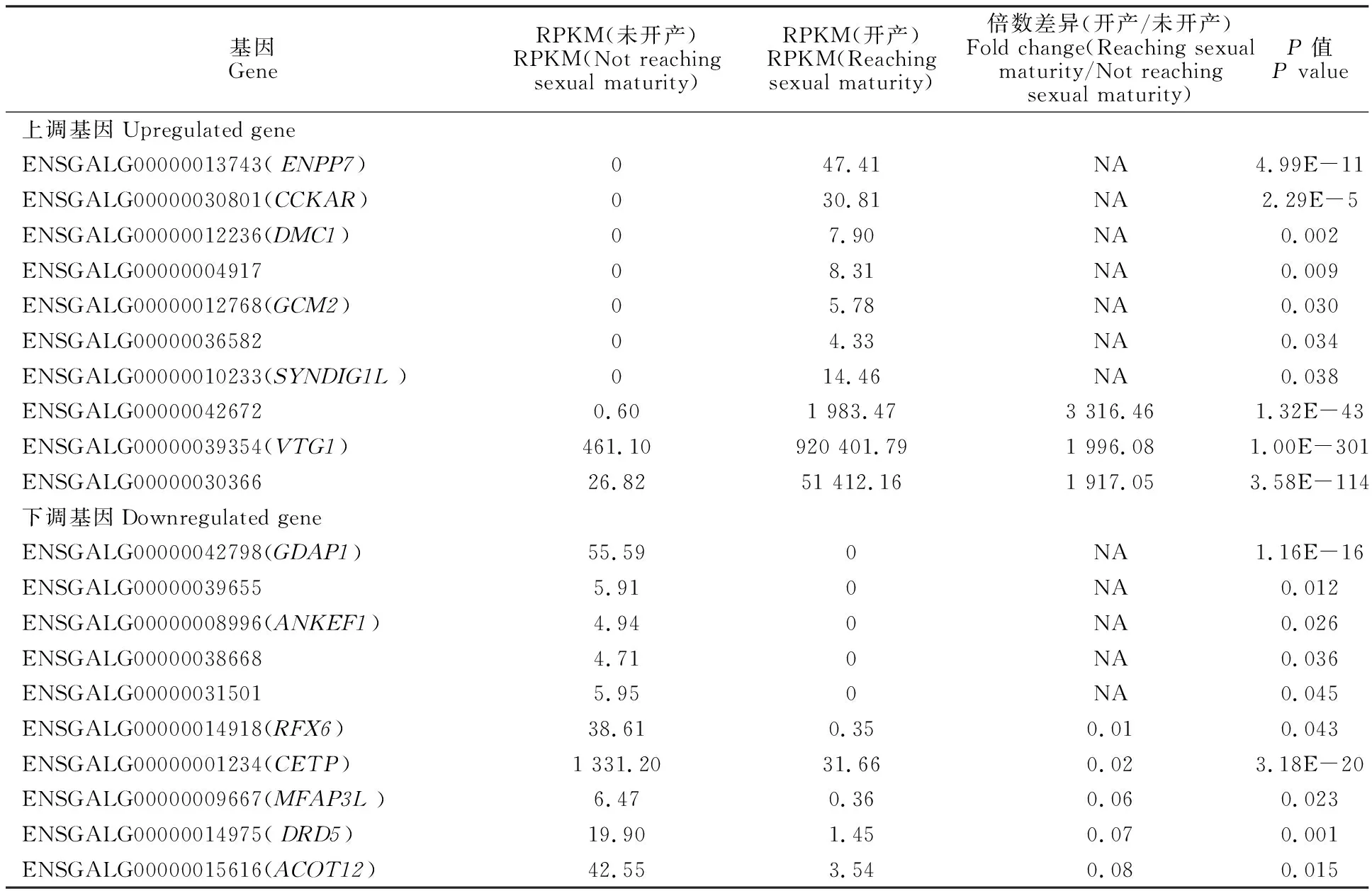
表4 表达差异最显著的20个基因Table 4 Top 20 differentially expressed genes
挑选部分差异表达基因进行qRT-PCR验证(图3),结果可见,4个基因表达量的qRT-PCR结果与RNA-seq结果基本一致,说明差异表达基因分析结果较可靠。
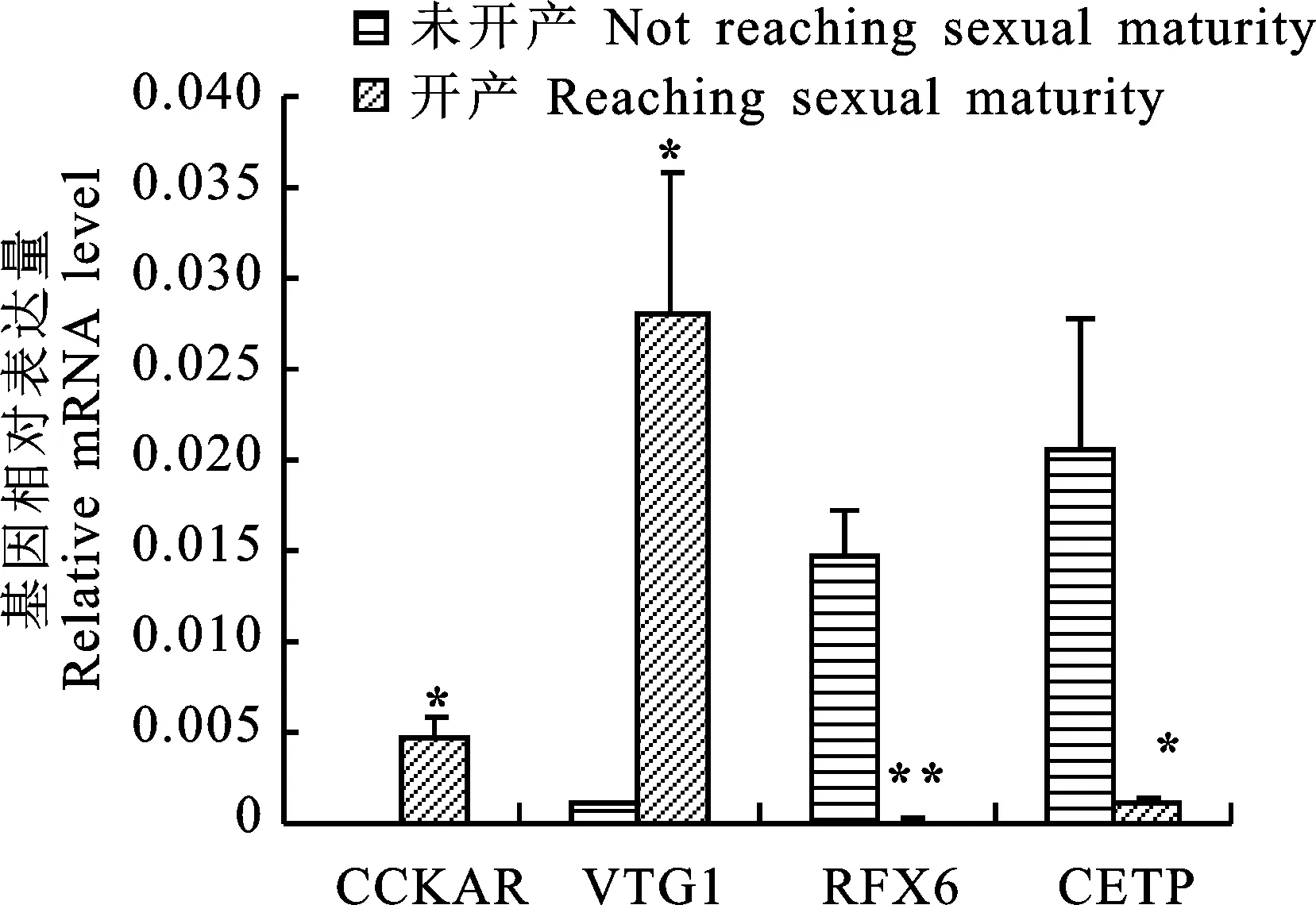
*.P<0.05; **.P<0.01
2.3 肝脏差异表达基因的功能分析
2.3.1 GO功能富集分析 对开产鸡和未开产鸡的差异表达基因进行GO功能富集分析,发现有60个显著富集的GO功能(FDR<0.05),其中44个富集在生物过程,11个富集在细胞组分、5个富集在分子功能。图4显示富集最显著的20个GO功能,除少数细胞组分的GO高度富集外,大部分均为脂质代谢相关功能,说明脂质代谢对开产很重要。这些GO功能富集的基因多则53个,少则5个。
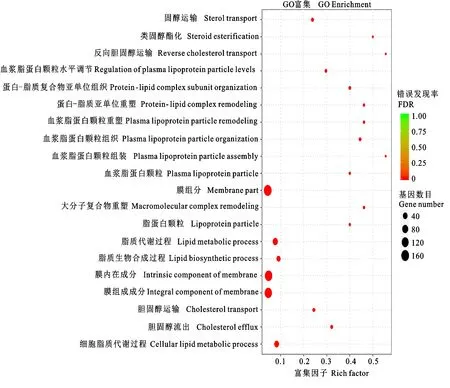
图4 差异表达基因GO富集分析:富集最显著的20个GO功能Fig.4 GO enrichment analysis of differentially expressed genes: top 20 enriched GO
2.3.2 KEGG信号通路富集分析 对开产鸡和未开产鸡的差异表达基因进行KEGG信号通路富集分析,发现仅有1个显著富集的KEGG信号通路(FDR<0.05),即非不饱和脂肪酸生物合成(Biosynthesis of unsaturated fatty acids)(表5)。此为脂质代谢相关的信号通路,包含基因FADS1、FADS2、ACOT7、ACAA1和ELOVL6,说明非不饱和脂肪酸生物合成信号通路与开产有密切关系。

表5 差异表达基因KEGG富集分析Table 5 KEGG enrichment analysis of differentially expressed genes
3 讨论与结论
高通量测序技术发展迅速,现在较常用的RNA-seq数据量一般都是千万级别的原始序列[22-23],本研究RNA-seq所得原始序列就在千万级别。同时,各组测序数据能比对上基因组的序列均在80%左右,而其中比对上基因的序列均超过80%,结果表明转录组测序所得数据质量较高,后期分析结果更准确,更可信。
对差异表达基因进行RT-qPCR验证,证明转录组测序结果比较可靠。进一步分析后,发现在差异最大的20个基因中,有7个(ENPP7、CCKAR、DMC1、GCM2、SYNDIG1L、ENSGALG00000004917、ENSGALG00000036582)和 5个(GDAP1、ANKEF1、ENSGALG00000039655、ENSGALG00000038668、ENSGALG00000031501)基因分别在低能组和高能组未检测出表达,ENPP7与肿瘤风险相关[24],CCKAR与生长和体质量相关[25],DMC1与减数分裂相关[26],GCM2与维持甲状旁腺功能相关[27],SYNDIG1L与哺乳动物乳头数相关[28],GDAP1与线粒体分裂等相关[29],其他基因尚无功能研究报道。这些基因在肝脏对能量摄入量的影响中虽然发挥了重要作用,但是机理尚不明确,需要进行更深入的研究。
对差异表达基因进行GO富集分析,发现与脂质代谢相关的功能在富集程度最高的20个功能中最多,共有16个GO,说明脂质代谢过程是对开产影响最大的生物过程。李云雷等[30]对北京油鸡开产前和开产后腹脂与性发育的关系研究发现,开产前腹脂质量与卵巢质量和卵巢指数呈中等正相关,表明育成期性腺发育一定程度上受脂肪沉积的影响;开产后腹脂质量与卵巢质量、卵巢指数和输卵管质量均呈中等程度负相关,且低腹脂率组的开产率高于高腹脂率组,其开产一致性较好,腹脂沉积过量导致北京油鸡性成熟和开产延迟。对卢氏鸡青年鸡及产蛋鸡肝脏转录组的研究发现,鸡肝脏脂质转运相关基因MTTPL影响蛋鸡性成熟[31]。另外,对白来航鸡产蛋前后的肝脏转录组测序研究也发现RNF186等与脂质代谢相关的差异表达基因,提示它们影响蛋鸡的产蛋性能[32],这些研究均表明脂质代谢及其相关基因对蛋鸡产蛋有重要影响。
对差异表达基因进行KEGG富集分析,发现唯一显著富集的通路非不饱和脂肪酸生物合成也与脂质代谢相关,通路中差异表达基因FADS1、FADS2是一类脂肪酸脱氢酶基因,参与多种多不饱和脂肪酸的合成,表达量与脂肪酸组成显著相关[33-34]。ACOT7是脂酰辅酶A硫脂酶7,能将脂酰辅酶A水解成游离脂肪酸,参与调节游离脂肪酸含量[35]。ACAA1是乙酰辅酶A酰基转移酶1,催化β-氧化途径的最后一步,参与脂肪酸的延伸和降解[36]。ELOVL6是长连脂肪酸延伸酶6,能催化延长C12-C16的饱和或单不饱和脂肪酸,调节脂肪酸组成总体平衡[37]。而李慧锋等[32]对高产鸡产蛋前后肝脏差异表达基因进行KEGG分析,发现富集的通路及相关基因主要与脂质代谢和氨基酸代谢相关,由此可见,脂质代谢相关信号通路确实影响蛋鸡开产。
综上所述,育成期高能饲喂的蛋鸡中,与肝脏脂质代谢相关的一些基因差异表达会影响肝脂代谢,最终影响开产时间。
猜你喜欢
今日农业(2022年2期)2022-11-16
畜牧兽医学报(2022年7期)2022-08-05
今日农业(2022年4期)2022-06-01
家庭科学·新健康(2020年6期)2020-07-06
科学导报(2019年45期)2019-09-23
保健与生活(2019年23期)2019-09-10
分析化学(2018年1期)2018-01-18
农家科技下旬刊(2017年7期)2017-08-22
山东农业科学(2016年4期)2016-11-19
农村百事通(2016年6期)2016-05-14
