
两种石漠化等级下樟树-马尾松混交林林分结构及物种多样性
2022-05-18 03:39陈双双朱宁华周光益
中南林业科技大学学报 2022年4期
陈双双,党 鹏,朱宁华,周光益,尚 海
(1.中南林业科技大学 林学院,湖南 长沙 410004;2.中国林业科学院 热带林业研究所,广东 广州 510520;3.武陵山石漠化综合治理国家长期科研基地,湖南 永顺 416700)
石漠化是指在热带、亚热带的湿润或半湿润气候下,伴随着极度发育的岩溶现象,加之人为干扰,导致地表原有的植被遭到破坏,大面积基岩裸露在外的现象[1]。湘西自治州青坪国有林场地处武陵山区中部,石漠化是导致武陵山区经济贫困、生态恶化的主要因素之一。目前,该区存在严重石漠化的区域多达3 万多平方公里,约占该地区总面积的30%。
森林植被的重建与恢复是石漠化地区生态恢复的关键[2-3],包括种子扩散、幼苗的定居与生长、幼树更新等多个环节,每个环节都不同程度地受到环境的影响[4]。同时,林分空间结构作为森林更新的驱动因子[5],它不仅反映了种群的属性和数量特征[6-7],还对物种的出现与消亡产生重要影响[8-9]。相关学者[10-11]利用混交度、大小比数和角尺度等林分空间结构参数进行研究,表明林分结构的差异性对林下树种的分布[12]和幼苗更新[13]会产生不同影响,而林下幼苗的分布格局是影响未来森林群落结构的关键[14]。同时,林分空间结构对物种多样性也存在影响,如混交度是影响林下植物多样性的重要因素[15]。但针对不同石漠化程度下林分空间结构与物种多样性之间的关系研究仍处于空白,本研究以青坪国有林场樟树-马尾松混交林为研究对象,对林分空间结构、物种多样性的变化规律进行研究,探讨两者之间的关系,以期为后续石漠化地区植被恢复提供造林树种选择的理论依据。
1 研究区概况与研究方法
1.1 研究区概况
试验地位于湖南省湘西自治州永顺县的青坪国有林场(29°2′58.1″N,110°13′35.44″E),大陆性季风气候,年降水量1 300~1 500 mm,年均温度16.3℃。该地区遍布石灰岩,土壤以黄壤为主,土层分布不均、厚度不一。该地区前身为茅山坡,岩石大面积裸露,属于极严重石漠化区域(图1)。1965年后开始人工补植造林,如今造林已取得显著成效。林场内的樟树-马尾松混交林为1980年等比例栽种,每个树种按照3 m×3 m 的株行距造林,但其间经过两次抚育间伐,现保存密度为1 125~1 450 株/hm2。
1.2 研究方法
1.2.1 样地设置及调查
在对樟树-马尾松混交林进行全面踏查后,采用典型样地取样调查法,选取立地条件大体一致的样地,按照岩石裸露率、植被类型、植被综合盖度和土层覆盖厚度等因素进行石漠化等级评分[16],确定为Ⅰ轻度石漠化和Ⅱ重度石漠化两种类型。共设置6 块20 m×40 m 固定标准地,调查林分的郁闭度、海拔、坡度、幼苗更新及枯立木等情况。按照10 m×10 m 划分小样方,对小样方内所有胸径≥1 cm 的树挂牌编号,进行每木检尺,记录树种名称、胸径、树高、枝下高、冠幅、位置。随机在每块样地中各设置3 块2 m×2 m 的有代表性的灌木样方,并在每个灌木样方中设置1 个1 m×1 m 的草本样方,调查其灌草种类、高度、盖度、株数等因子[17]。
标准地的基本情况及石漠化等级评分情况分别如表1~2所示。
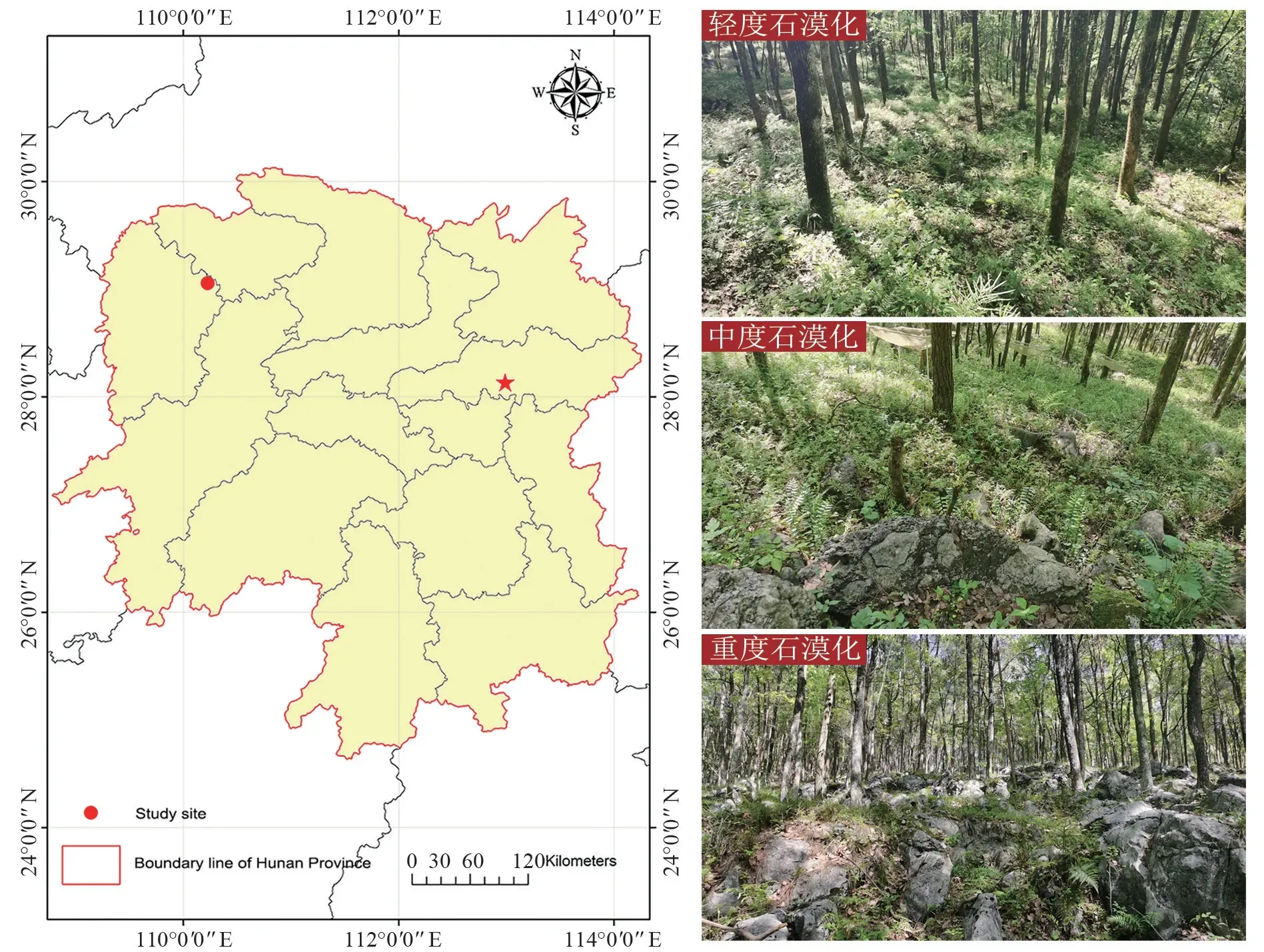
图1 研究区位置及石漠化现状Fig.1 Location of the study area and status of rocky desertification
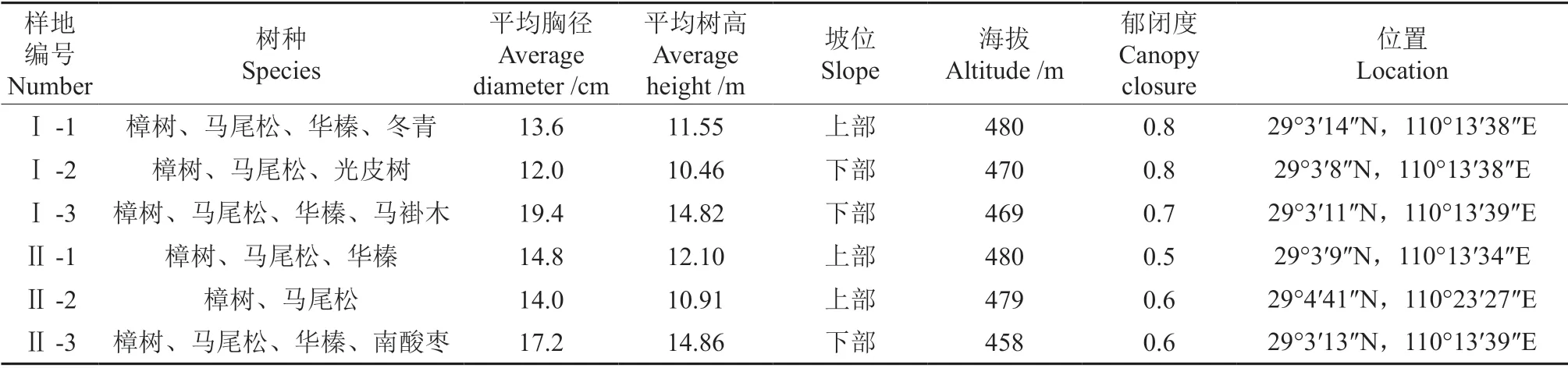
表1 样地基本概况Table 1 Situaion of plots
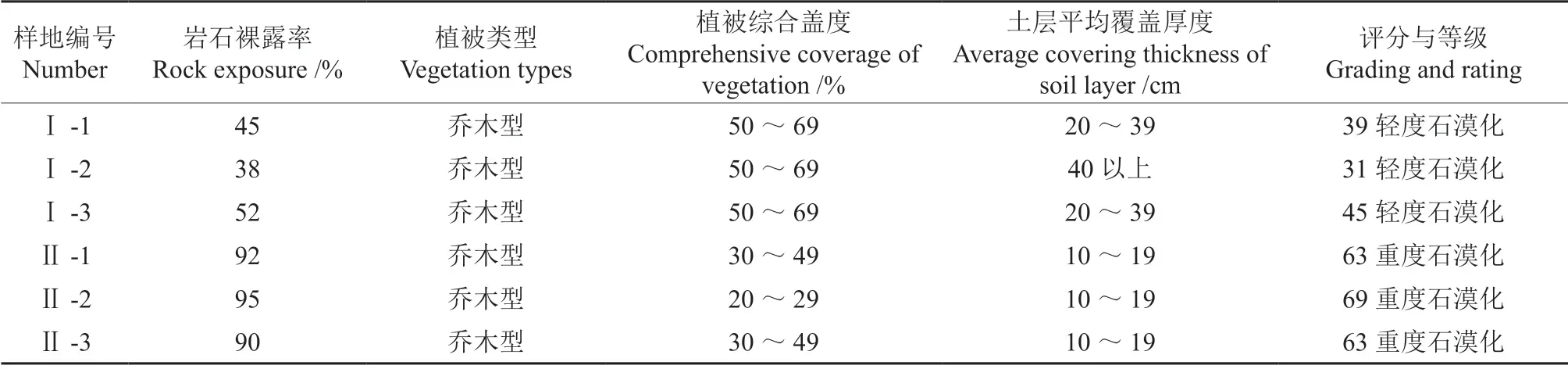
表2 石漠化等级评定因子及结果Table 2 Evaluation factors and results of rocky desertification grade
1.2.2 空间结构参数及评价
在样地周围设置5 m 缓冲区,以对象木及其相邻的4 株树构建空间单元,绘制样地树种分布图(图2),并计算空间结构参数(混交度、角尺度、大小比数)[18]。计算公式如下:
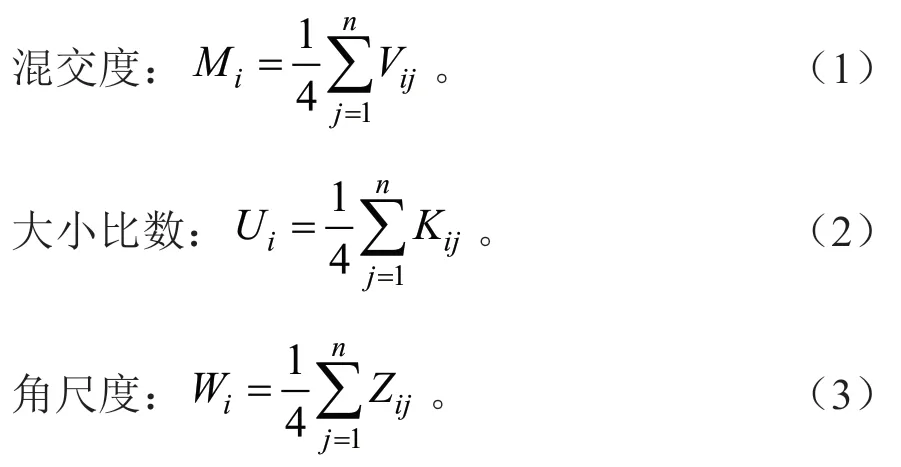
式(1)~(3)中:n为邻近木株数,若对象木i与邻近木j的树种不同,Vij=1;否则,Vij=0。若对象木i比邻近木j小,则Kij=1;否则,Kij=0。若对象木i与邻近木j的夹角小于标准角,则Zij=1;否则,Zij=0。
为研究两种石漠化等级下整体林分空间结构和同一等级下不同样方内的结构差异,利用空间结构参数计算林分空间结构指数(FSSI)和林分空间结构距离(FSSD)[19],对此进行整体评价。计算公式如下:
林分空间结构指数:

式(4)中FSSI、M、U、W取值范围为[0,100],FSSI 越大,林分结构越理想,即当(FSSI=100,M=100、U=0、W=50)时,林分达到最理想的结构。
林分空间结构距离:
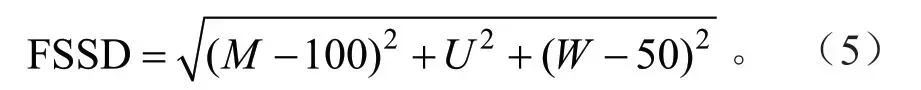
式(5)中FSSD 取值范围为[0,150],M、U、W取值范围为[0,100],当M=100、U=0、W=50 时,FSSD 越小,林分结构越理想,即从现实林分到理想林分距离最短。
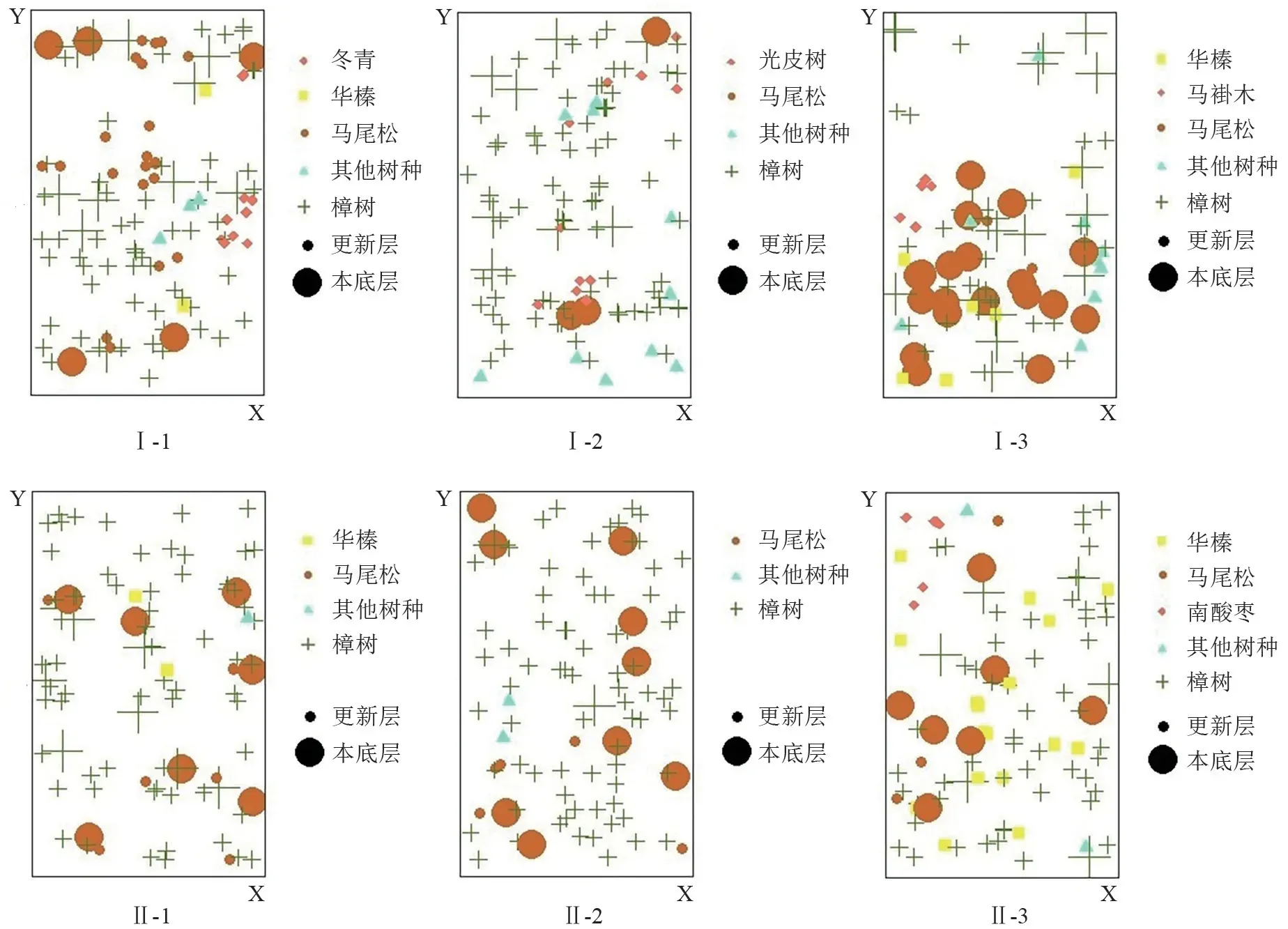
图2 调查样地树种分布Fig.2 Distribution of tree species in the sampling site
1.2.3 物种多样性指数
在野外调查数据的基础上,计算样地内灌草层不同物种的重要值(Ⅳ),并以此为基础,计算物种多样性指数[20]。具体公式如下:

式(6)~(10)中:Pi=Ni/N;Ni为第i个种的个体数量,N为总个体数量。S为样地内物种数目[21]。
1.2.4 数据分析
数据分析主要使用winklmass 和Excell 软件对林分空间结构参数进行计算,使用R-4.0.3 软件进行物种多样性指数计算;同一林分内胸径使用单因素方差分析中的Duncan 检验进行显著性检验;不同石漠化等级下林分空间结构与物种多样性的关系进行Pearson 相关性分析。数据作图均在R-4.0.3 软件中进行。
2 结果与分析
2.1 物种组成重要值
重要值可以反映出某一物种在森林群落中的作用和占据的地位,通过对重要值的分析可以找到该群落中的优势种[22-23]。樟树-马尾松混交林林下灌草较为丰富,通过统计调查,林下物种共有78 种,隶属于46 科73 属,其中灌木层28 科39 属43 种,草本层25 科35 属35 种(表3)。对林下植被重要值>10%的物种进行分析,发现:随着石漠化程度的加重,灌木层物种重要值逐渐高于草本层。研究区乔木层隶属于14 科14属14 种,其中以樟树Cinnamomum camphora、马尾松Pinus massoniana、华榛Corylus chinensis为优势种。不同样地内还分布着特有的树种如冬青Ilex chinensis、光皮树Cornus wilsoniana、马褂木Liriodendron chinense、南酸枣Choerospondias axillaris等(图2)。随着石漠化程度的加重,马尾松的比例大幅降低,而樟树的比例则大幅度上升,这表明马尾松在石漠化地区的天然更新能力较差。何友均等[24]发现在马尾松林群落演替过程中,本土阔叶树种的天然更新能力优于马尾松林。随着阔叶树种的增加,土壤肥力得到改善,从而促进林分的更新[25]。在本研究中,南酸枣、光皮树、马褂木等阔叶树种均为天然更新而来,能够适应当地环境条件,且均为阳性树种,马尾松冠层较好的透光条件为这些树种的生长提供了有利条件[26],这表明经过一段时间的更新后,该林分总体演替趋势良好。

表3 样地乔灌草层群落重要值Table 3 Important values of tree,shrub and herb layer community in the sample plot

续表3Continuation of table 3
2.2 林分树种胸径分布
不同石漠化程度下各样地树种胸径分布见图3。如图所示,同一样地中,不同树种的胸径分布存在显著性差异(P<0.05)。樟树在整个林分中占有绝对优势地位,胸径分布整体较为均匀,更新情况良好,小径阶马尾松在重度石漠化区域更新表现更为优良。总体而言,轻度石漠化区域乔木层更新更好,物种丰富,小径阶的其他树种(冬青、光皮树、马褂木等)的生长状况都比较优良,存活率较高。
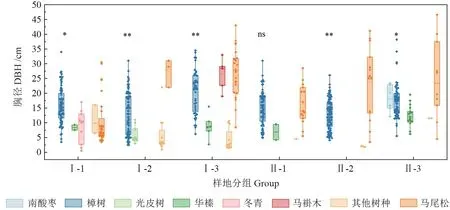
图3 各样地不同树种胸径分布Fig.3 Diameter distribution of different species of each plot
2.3 林分空间结构参数
从表4可以看出,重度石漠化地区林分平均混交度为0.38,处于弱度混交和中度混交的中间状态,其中只有Ⅱ-2 号样地的零度混交的比例达到43%,另外两块样地基本都处于弱度混交以上状态,说明Ⅱ-2 号样地树种混交程度低,林分不稳定。而轻度石漠化地区林分平均混交度为0.47,非常接近中度混交。其中Ⅰ-3 号样地处于中度和强度混交的中间状态,是6 块样地中混交程度最好的。总体而言,轻度石漠化地区林分混交程度比重度石漠化地区高,林分更加稳定,这与乔木层树种组成结构一致。
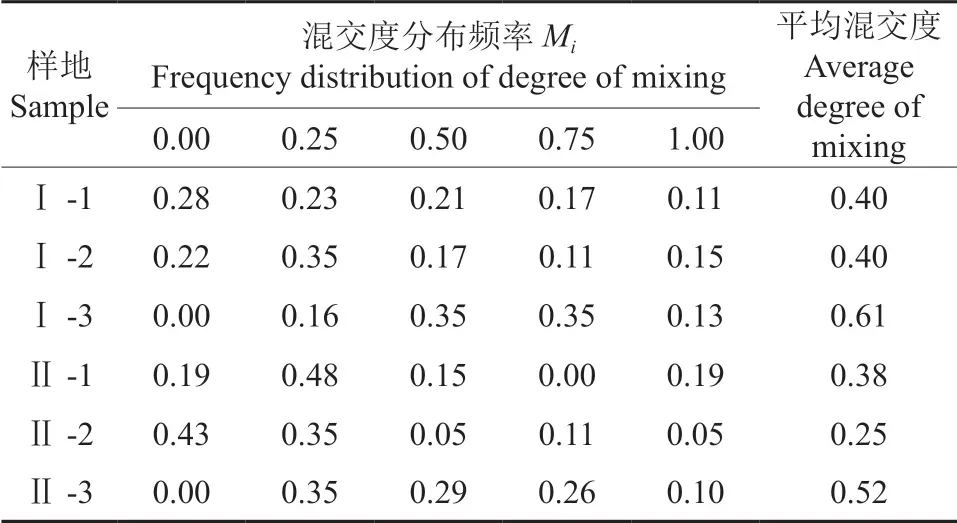
表4 各样地林分混交度分布频率†Table 4 Frequency distribution of mixing degree of tree species
表5表明,轻度和重度石漠化地区平均大小比数都为0.49,表现为接近中庸状态。根据全林分平均大小比数的分布频率来看,6 块样地中Ⅱ-2、Ⅰ-2、Ⅱ-3 号都处于中庸状态,另外三块则处于亚优势和中庸之间。这说明樟树-马尾松混交林单个样地不同树种之间胸径大小有所差异,但林分整体没有出现明显分化。轻度石漠化地区处于优势状态的频率比重度石漠化地区相对大一些,且大小比数在各个等级上的分布频率也基本均匀。
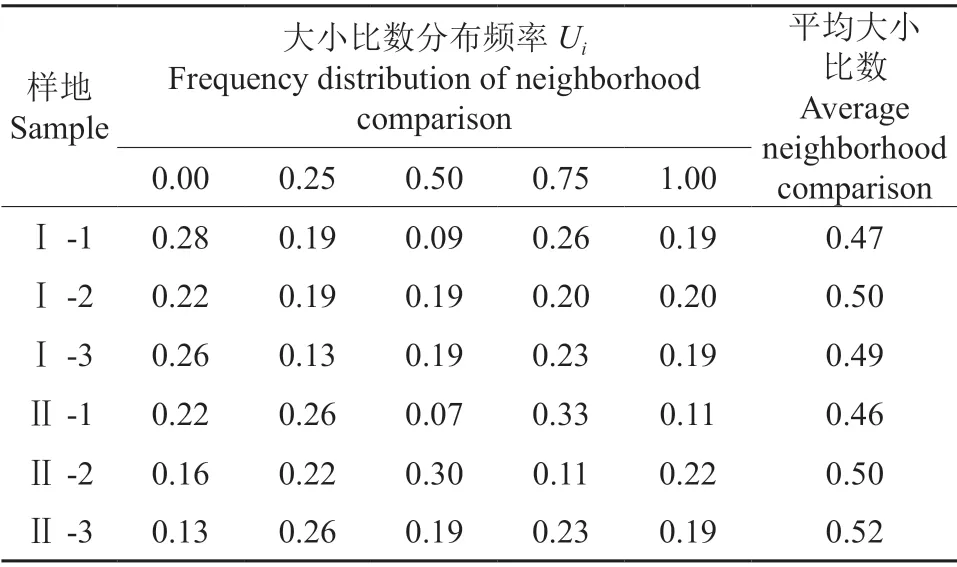
表5 各样地林分大小比数分布频率†Table 5 Frequency distribution of neighborhood comparison of tree species
从表6可以看出,从整体林分来看,处于随机分布的频率是最高的,基本超过了均匀分布和不均匀分布的比例之和,说明整个樟树-马尾松混交林林分的单株木以随机分布为主。重度石漠化地区平均角尺度为0.50,轻度石漠化地区平均角尺度为0.56,两者相差不大,都表现为随机分布,但轻度石漠化地区更接近聚集分布,说明重度石漠化地区林分树种聚集分布的程度很低。
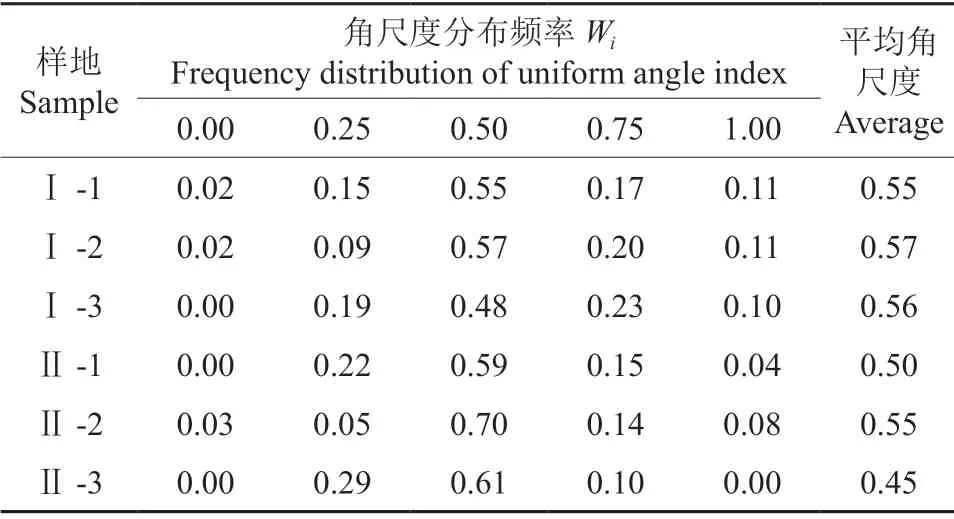
表6 各样地林分角尺度分布频率†Table 6 Frequency distribution of uniform angle index of tree species
2.4 林分空间结构评价
林分空间结构指数(FSSI)能定量反映现实林分空间结构的状态和变化动态,林分空间结构距离(FSSD)进一步反映了现实林分与理想林分空间结构之间的差距[20]。FSSI 越大,FSSD 越小,林分空间结构越好。从表7可知,Ⅰ-3 样方的林分空间结构最好,Ⅱ-2 的结构最差,这与各样地混交状态结果一致,表明混交度在林分空间结构评价指标中占据主导地位。轻度、重度石漠化区域的FSSI、FSSD 均值分别为59.21、72.49、55.80、79.54,表明轻度石漠化区域的樟树-马尾松混交林空间结构更为良好,但这两个区域离达到理想林分空间结构均存在较大差距。
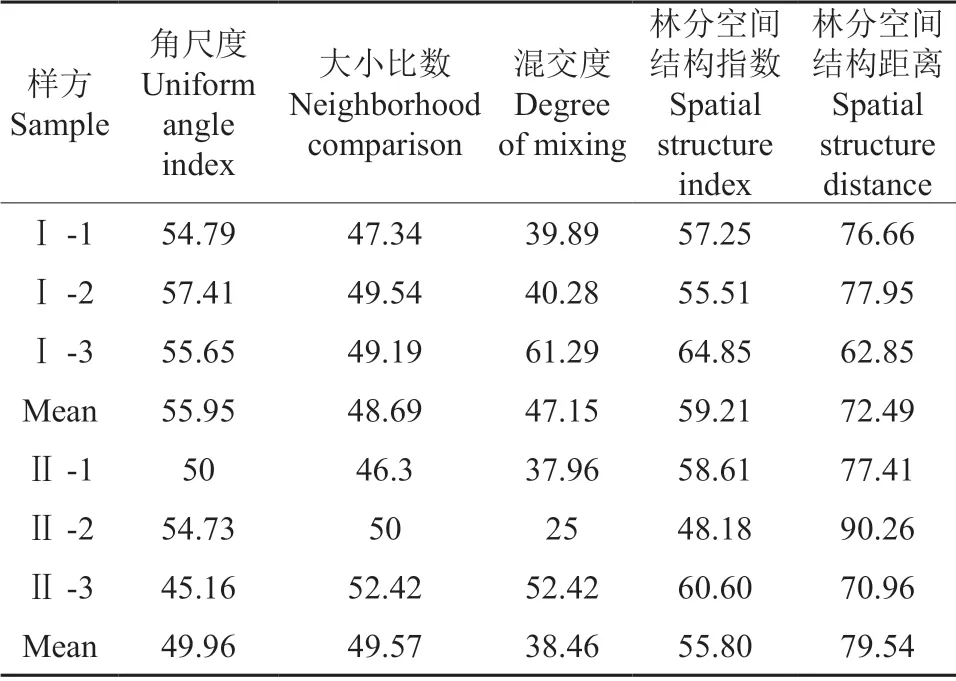
表7 林分空间结构指数与空间结构距离Table 7 Stand spatial structure index and spatial structure distance
2.5 物种多样性指标
由表8可知,随着石漠化程度的加重,灌草层的Shannon 指数、Pielou 指数、Simpson 指数、物种数都有所降低。轻度石漠化区域灌木层的物种多样性指数比草本层普遍较高,相反地,重度石漠化区域灌木层的Simpson 指数小于草本层,Shannon 指数相差不大,这是由于草本层多为一年生植物,根系分布较浅[27],具有易生长和生长快的特点[28],比灌木丛在土壤瘠薄的地区更易存活,分布更广。
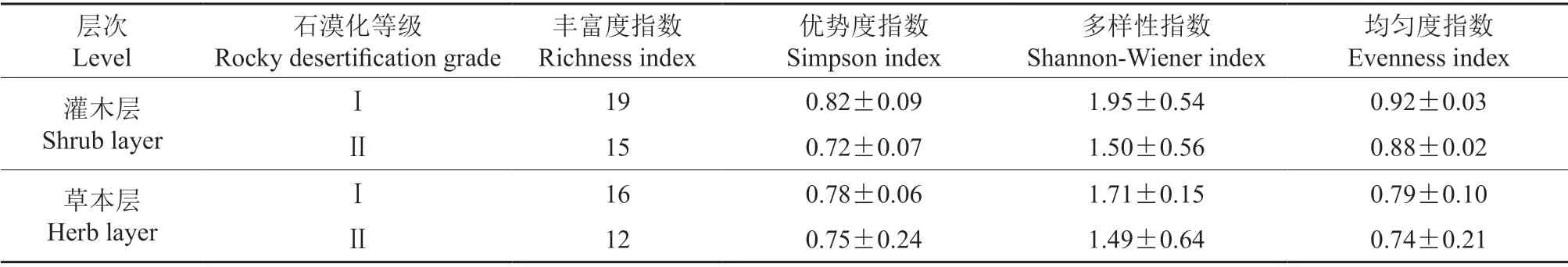
表8 不同类型林分林下植被物种多样性Table 8 Species diversity of understory vegetation in different types of stands
2.6 混交林空间结构参数与物种多样性指标的相关分析
由表9可以看出,灌草层中混交度和大小比数和物种多样性指数二者之间均不存在显著相关性(P>0.05),角尺度与灌木层Shannon 多样性指数、Simpson 优势度指数存在显著正相关性(P<0.05),但与草本层物种多样性指数的相关性均不显著。混交度与角尺度和石漠化等级程度之间均为负相关,大小比数与石漠化等级程度为正相关,相关性均不显著(P>0.05)。
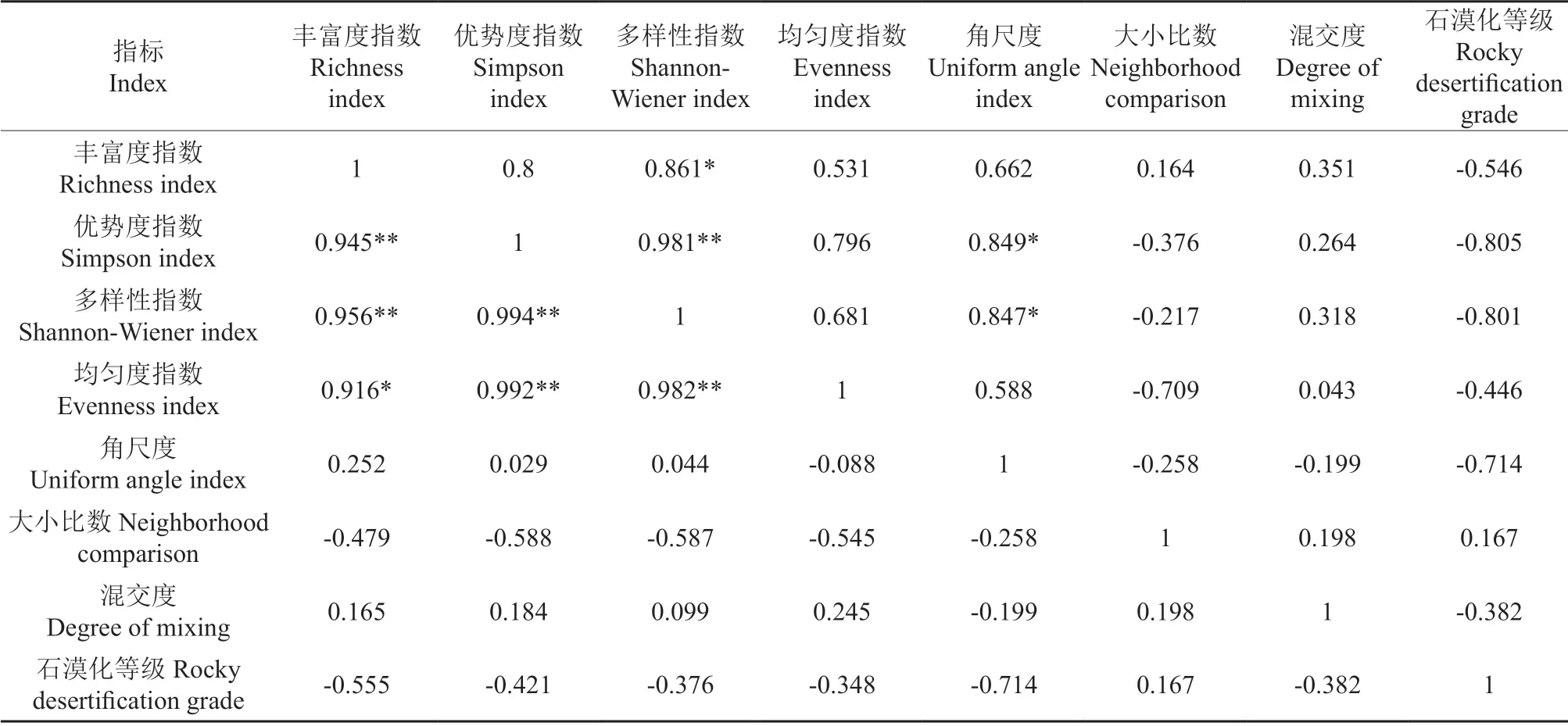
表9 不同石漠化等级下林分空间结构和物种多样性之间的相关性†Table 9 Correlation between stand spatial structure indexes and diversities in different grade of rocky desertification
3 结论与讨论
通过对两种石漠化程度下樟树-马尾松混交林林分空间结构参数、物种多样性的变化规律以及空间结构参数与物种多样性的关系进行分析,发现:经过40 a 的演替,研究区乔木层由原来的樟树、马尾松等比例混交林结构逐渐演变为以樟树为主,马尾松为辅,零星分布着其他树种(华榛、南酸枣、马褂木等)的异龄复层群落。综合分析发现,林下植被对马尾松幼苗的定居存在不利影响,樟树等其他阔叶树种的迅速成长,导致林分郁闭度增大,透光减少,不利于马尾松幼树的生长,因此马尾松种群呈现衰退趋势。轻度石漠化区域的林分空间结构明显优于重度石漠化区域,但距离理想林分空间结构仍有差距。就目前林分结构而言,建议对樟树进行适度间伐,以促进其他树种的更新,加快群落演替进程,构建更为稳定的群落结构,从而有利于石漠化地区的森林恢复与重建。
物种多样性是维持生态系统功能和结构稳定的基础[29]。研究发现,轻度石漠化地区林下物种多样性指数比重度石漠化地区更高,物种更丰富。从重要值来看,灌木层普遍大于草本层,且耐阴植物逐渐增加,部分阳性植物的重要值逐渐下降直至消亡。这与林分的光照条件存在一定的关系,在造林初期,草本层物种多为喜阳草本,随着演替的进行,乔木层树种生长茂盛,林分郁闭度变大,透进林内的光线逐渐变少[30],加之灌木的遮盖作用,导致灌木生长迅速,而草本生长缓慢。在重度石漠化地区,由于土壤瘠薄,土被分布不均,浅根系的草本比灌木更易存活。从乔木层重要值能明显看出,樟树占比最大,超过60%,其次是马尾松、华榛。与当初造林相比,整体林分中出现了很多外来树种。这表明除母树天然更新外,还有其他树种通过其他更新方式促进了林分更新,根据野外踏查结果,发现在样地的附近存在成片的光皮树、马褂木林分,但南酸枣、冬青未有发现。可能是由于南酸枣、冬青的果实色泽艳丽,肉质鲜美,鸟类喜食,将其带到此地[31]。
植物个体的生长、死亡和更新都会影响到群落的空间结构,林分空间结构是否优良是判断群落能否维持稳定发展的重要前提[32]。通过对两种石漠化程度下林分空间结构进行评价,发现混交度在FSSI 和FSSD 体系中占据重要地位。轻度石漠化区域接近中度混交,乔木层树种更为丰富,林分结构比重度石漠化区域更稳定。研究发现,马尾松的径级大小和分布情况影响林分的整体结构。结合树种和胸径结构分布图,发现樟树接近随机分布,而马尾松幼树则多聚集于母树周边,实地观察发现马尾松松果多常见于粗壮的母树周围,聚集分布有利于小径级个体的存活和生长[33]。
从林分更新情况来看,樟树在林分构成中占有绝对优势地位,而马尾松正在逐渐被其他阳性阔叶树种所替代。这主要是由于树种自身的生物学特性和其生存环境共同决定[34]。研究指出,樟树能适应喀斯特地区的立地条件,其水平根系发达,可以穿透岩石缝隙,且种子结实多,除依靠重力传播种子外,鸟类的镊食行为也使其大面积扩散,因此它在群落中自然更新良好[1]。但由于石漠化地区土壤瘠薄,养分较少,樟树的生长情况不如其他地区。马尾松作为常见的先锋树种,喜光、耐干旱瘠薄,能够在石漠化地区存活。稀少的林下植被可以使马尾松幼苗获得充足的光照资源,但也易使其面临极端的干热化胁迫[35],所以重度石漠化区域虽出现马尾松更新,但数量不多。轻度石漠化区域林下植被较为丰富,加上上层乔木的遮阴作用使部分幼苗出现死亡,最终导致小径阶的马尾松在种群结构中比重下降,马尾松成树个体的数量显著提升,这表明林下植被在一定程度上能够影响马尾松的生存环境。小径阶马尾松的大量死亡,这表明该区域马尾松种群已经步入衰退状态,预测随着演替的进行马尾松种群会逐渐被其他阔叶树种所替代[36]。当然,除了光照条件,土壤、坡位、海拔等生境因子对马尾松种群的衰退是否存在影响尚未可知,仍需进一步研究。
林分空间结构对林分物种多样性存在较大的影响[37-38]。前人研究发现混交度是影响物种多样性的关键[17],但本研究尚未得出此结论,可能由于紫金山为城市森林,人为干扰严重,所选样地林木资源丰富,而本研究所选样地人为干预较少,前期为荒山裸地,虽人工补植过,但立地条件差,自然更新缓慢,混交度较低。针对前人的结论,仍需等待一段时间的演替后,再进行验证。同时,本研究发现角尺度对灌木层物种多样性存在显著相关性,这与张亚昊[39]的研究结果一致。研究也发现,不同石漠化程度下的角尺度差异显著,不同石漠化程度导致林分树种隔离程度不同,从而影响群落内部的物种多样性的变化。据统计,除冬青以外,群落内新出现的树种均为落叶乔木,适应性强,能在钙质土上生存,预测该地区未来会朝着以落叶阔叶林为主的异龄复层混交林进行演替[40]。
本研究只是针对轻度、重度石漠化区域的樟树-马尾松混交林的空间结构和物种多样性进行初步探讨,今后仍需增补潜在石漠化和中度石漠化区域的研究,从而形成完整的石漠化演替研究体系。针对林分空间结构与环境因子的研究尚未进行深入探讨,以后可以通过测定土壤物理化学特征、温度、降水等因子,探讨环境因子与林分空间结构因子之间的关系,为进一步探明上层乔木树种与林下植被、环境因子之间的相互作用机理提供科学依据。
猜你喜欢
绿色天府(2022年9期)2022-11-19
世界热带农业信息(2022年8期)2022-07-19
贵州畜牧兽医(2022年3期)2022-06-28
南方农业·中旬(2021年5期)2021-10-25
中学生百科·小文艺(2020年8期)2020-01-13
安徽农学通报(2017年6期)2017-04-11
绿色科技(2017年3期)2017-03-14
安徽农学通报(2014年18期)2014-10-23
安徽农学通报(2014年9期)2014-06-23
