
北京奥林匹克森林公园蚊虫停落指数时间—空间分布特征与环境关系的研究*
2022-05-14 03:54刘美德张洪江李秋红葛军旗周小洁付学锋曾晓芃
寄生虫与医学昆虫学报 2022年1期
刘美德 张 勇 佟 颖 张洪江 李秋红 刘 婷 葛军旗 周小洁 钱 坤 付学锋 曾晓芃**
(1.北京市预防医学研究中心,北京市疾病预防控制中心,北京 100013;2.北京市朝阳区疾病预防控制中心,北京 100021)
蚊虫监测是预测病原体传播媒介的重要工具,可用于了解流行病的潜力并规划早期有效的控制策略。蚊虫停落指数是影响人与蚊虫接触之间准确关联的重要参数(陆宝麟等,1961; Hameretal.,2011)。从蚊媒流行病学的角度来看,它不仅表明蚊虫的密度,而且还预测了蚊媒疾病暴发的可能性(Jaletaetal., 2013)。因此,停落指数调查在保护人们免受蚊虫侵袭和预防蚊媒疾病方面起着关键作用(Wallaceetal., 2014)。
自然条件下不同时间尺度和不同空间范围内的停落指数可以为指导灭蚊提供重要信息。因此,许多研究集中在蚊虫停落指数随时间变化的特征(高强等, 2014a; Renetal., 2015)以及与环境特征的相关性(Cumminsetal., 2012; 高强等, 2014b)。尽管一些研究人员已经注意到环境对蚊虫停落活动的影响,但只关注人诱停落数据,而没有进一步定量分析环境特征与蚊虫停落指数之间的关系。随着3 S技术(特别是RS遥感和GIS地理信息系统)的广泛应用,研究人员能够通过环境特征的定量提取(Arjunanetal., 2015)应用于蚊虫种群指数研究(Regisetal., 2013)以及蚊媒疾病的预防(Espinosaetal., 2016)。本研究以北京奥林匹克森林公园(简称奥森公园)为实验场所,研究蚊虫停落指数的时空动态格局及其与周围环境特征的关系,明确降落率的时空分布特征,以期为人们避开蚊虫叮咬以及奥森公园内的灭蚊工作提供科学建议。
1 材料与方法
1.1 实验区域及监测点的设置
研究选择亚洲最大的城市绿化景观——北京奥森公园作为研究区域,该区域位于中国北京的中心地区,覆盖着约680 hm2的草场、湿地、一个带有中心岛的小湖以及茂密的灌木丛和高大的乔木;公园由城市环路分为南部公园和北部公园,这两部分通过模拟的自然生态走廊相连。采用文献(刘美德等, 2019) 方法,本研究在南园的东门(树林环境)、南门(草坪环境)、东北门(灌木环境)、湿地、中心岛分别确定一个监测点。为满足后期环境空间分析的需要,研究中使用GPS设备对每个监测点进行了定位;并基于GPS数据获取WorldView-2(美国DigitalGlobe公司)高分辩率卫星影像数据(分辨率为0.5 m)以用于后续遥感与空间分析。
1.2 环境遥感分析
采用文献(刘美德等, 2019) 方法,对奥森公园地面环境分类。应用GRASS 7.6.1采取无监督的k均值聚类算法(k-means clustering algorithm),基于WV2卫星遥感影像数据完成对研究现场环境的遥感分类。结合现场勘察,将奥森公园的环境因素大致分为住宅区(RA)-人类生活环境,开放水体(OW)-没有任何覆盖物的水体,乔木(HT)-高大乔木树林,灌木(LT)-人工或自然生长的低矮灌木,水生植物水体(GW)-种植或自然生长有水生植物的水体,草坪区(LA)-人工或自然草坪覆盖的区域等。基于奥森公园的WV2数据(2013年8月),应用GRASS 7.6.1中波段运算的方法计算与生成研究区域的NDVI值(归一化差异植被指数)栅格地图和NDWI值(归一化差异水指数)栅格地图(刘美德等,2019)。
1.3 空间分析
采用文献(刘美德等, 2019) 方法,基于奥森公园卫星影像,应用QGIS(3.8.3) 分别生成以监测点为中心不同空间距离(100、200、400、600、800、1 000 m半径)的缓冲区;使用空间分析方法提取每个缓冲区的各类环境定量数据,同时建立环境分类数据库(刘美德等,2019)。经遥感与空间分析,能够提取到各采样点不同缓冲距离(100、200、400、600、800、1 000 m半径)内的:人工建筑、开放水面、乔木、灌木、水体植物水体、草坪等地物面积,NDVI和NDWI值,以备后续相关与回归分析。
1.4 蚊虫停落指数的调查
本研究蚊虫停落指数实验始于2013年7月,并于当年9月结束。在每个调查月份中,停落指数实验每月进行2次,一次在月初旬,另一次在月末旬。每次降落实验持续约15 min,并在1 h内进行2次。随机选择5名志愿者在每个站点进行测试,计算测试时间内停落于他们身体上的蚊虫的只数,并用吸蚊器捕获停落的蚊虫。然后在下次测试中依次轮换他们的顺序,以避免个体引诱力带来的系统性误差。最后,捕获到的蚊虫被带到实验室进行形态学鉴定,停落指数的单位计为:只/(人·次)。
1.5 统计分析
应用SPSS 19.0软件对蚊虫停落指数与环境因素的关系进行Pearson相关与多元线性回归模型分析。根据AIC值和P值,筛选出与蚊虫停落指数相关的环境因素,并应用于多元线性回归分析。相关与多元线性回归模型统计分析中P值以0.05作为统计显著性的判断标准。
2 结果
2.1 蚊虫停落指数的时间动态
从7—9月,平均蚊虫停落指数呈逐步上升的趋势,分别为3.6、5.1和10.3只/(人·次);其中9月的停落指数达到峰值(图1)。
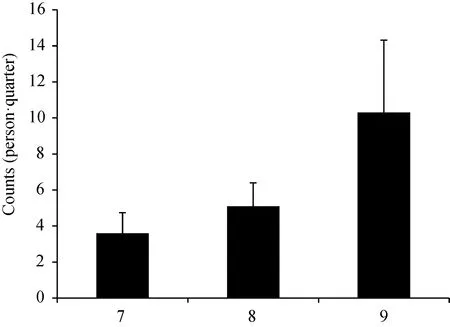
图1 7—9月平均蚊虫停落指数
在整个调查期间,奥森公园的5个点位的停落指数显示出空间异质性。9月在湿地、东门和中心岛的停落指数达到全年最高点;在东北门、南门和东门,最低停落指数在7月可见;在湿地和中心岛,最低停落指数在8月可见(图2)。各个点位中不同月份组别间停落指数比较情况如下:在东北门,月份组别间(F= 1.129,P= 0.365)以及所有月份之间差异均无统计学意义,即使在图形上显示为8月和9月高于7月的停落指数。在湿地地区,月份组别(F= 1.223,P= 0.344)以及月份之间差异均无统计学意义。在南门区域,月份组别(F= 0.444,P= 0.657)和月份之间差异均无统计学意义。在东门位置,月份组别间(F= 2.433,P= 0.158)和月份之间差异均无统计学意义。在中心岛区域,月份组别之间有统计学差异(F= 6.509,P= 0.021),其中8—9月的停落指数具有统计学意义(P= 0.007),而其他几个月份之间差异均无统计学意义。
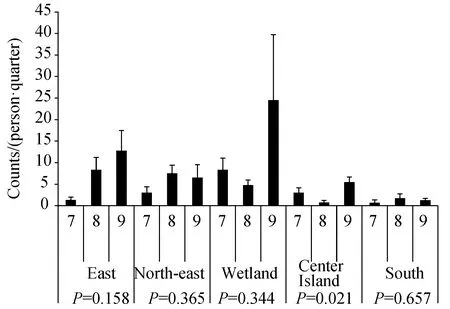
图2 不同点位上7—9月蚊虫停落指数
2.2 蚊虫停落指数的空间动态
在整个研究期间,湿地(13只/(人·次))的停落指数在5个地点中排名最高,然后按停落指数从高至低依次为东门、东北门、中心岛和南门,停落指数分别为8、6、3.17和1.5只/(人·次)(图3)。
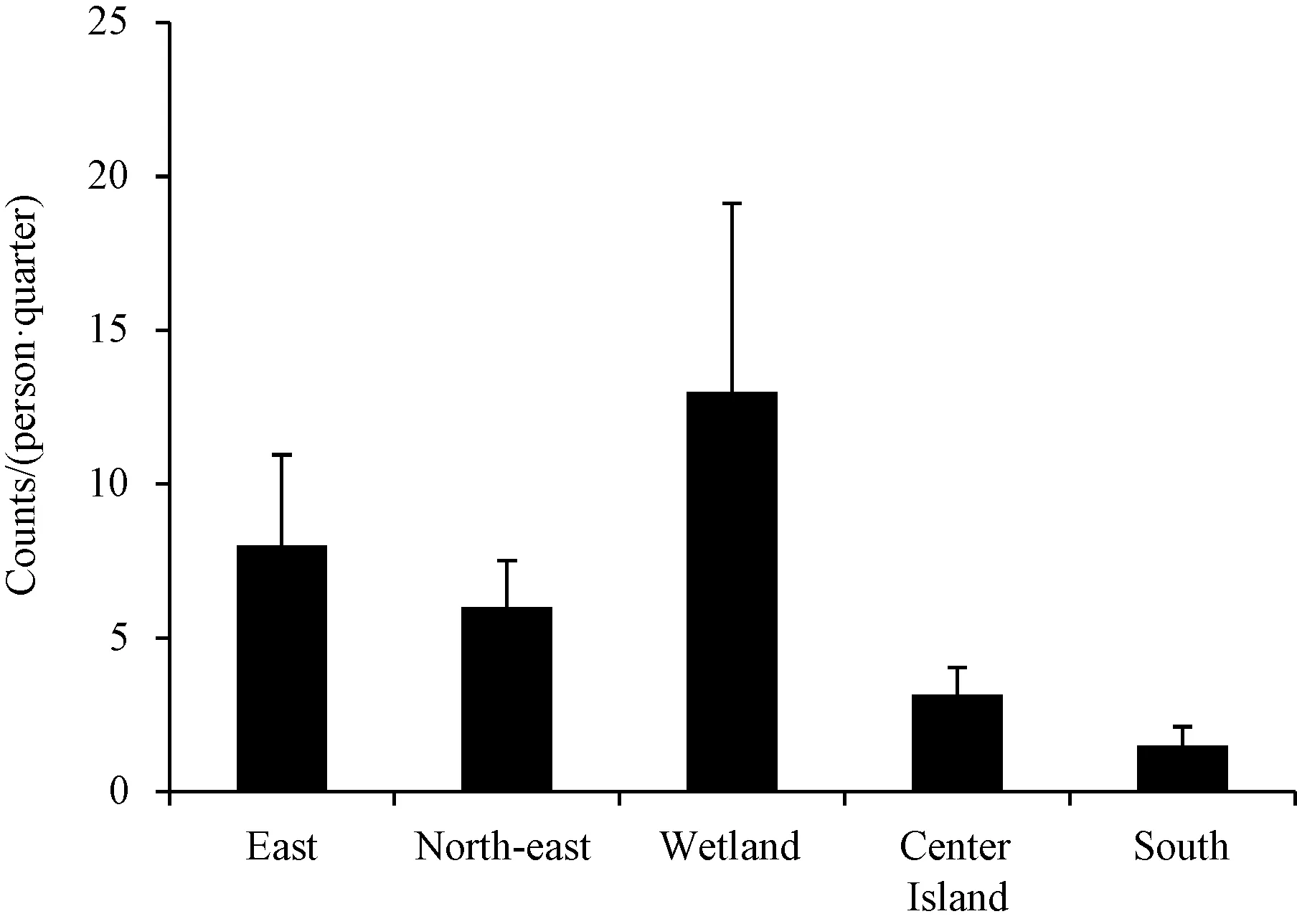
图3 不同点位蚊虫停落指数
从7—9月,奥森公园内的蚊虫停落指数在不同月份、不同站点之间显示出不同的分布模式。其中,7、9月在湿地出现了最高停落指数点位,在南部出现了2次最低停落指数点位(图4)。
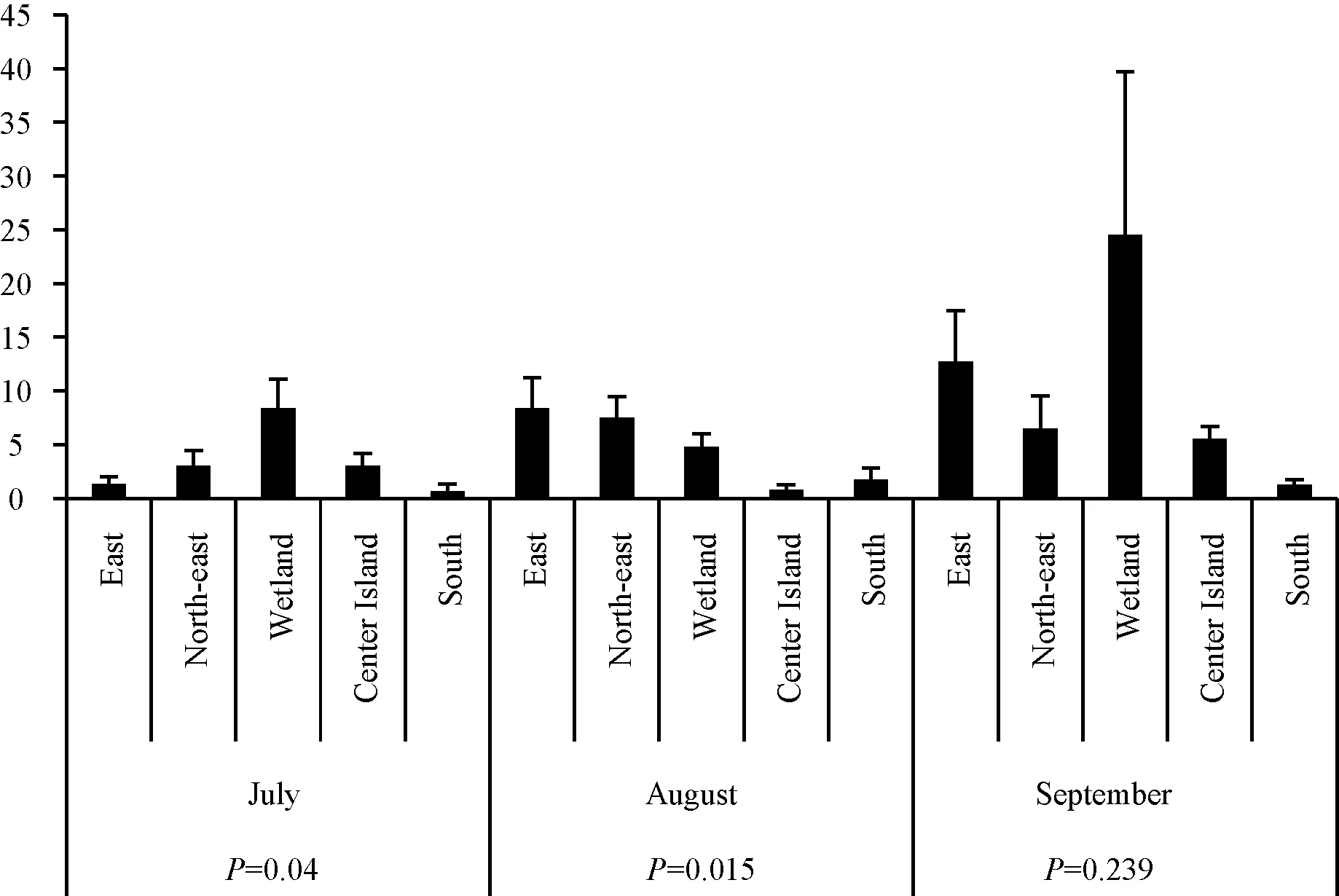
图4 7月至9月中各个点位的蚊虫停落指数
7月,蚊虫停落指数从高到低依次为:湿地、中心岛、东北门、东门和南门(图2)。此外,7月份各点位组别之间的差异具有统计学意义(F= 3.655,P= 0.04)。LSD分析结果还显示,湿地与其他4个地点之间的停落指数存在显著性差异(湿地对东北门:P= 0.027;湿地对南门:P= 0.006;湿地对东门:P= 0.009;湿地与中心岛:P= 0.036),其他点位组别之间没有显著性差异。
8月,停落指数在东门位置最高,而这里的停落指数高于7月停落点的最高点(在湿地)。8月份湿地的停落指数低于7月份相应点位,甚至低于东北门停落指数。5个地点的蚊虫停落指数从高到低按以下顺序排列:东门、东北门、湿地、南门、中心岛(图2)。此外,对5个点位级别进行了方差分析,8月份的站点组之间的差异具有统计学意义(F= 4.491,P= 0.015);此外,级别间的LSD分析结果还显示,各站点之间停落指数存在4个显著差异组,包括东北门与南门(P= 0.018),东北门与中心岛(P= 0.007),南门与东门(P= 0.013)和东门与中心岛(P= 0.006)。
9月,停落指数与7月一样,再次在湿地达到峰值,同时也是所有监测月份和所有点位中的峰值。9月份以及整个监视期间,东门点位的停落指数在所有月份的各点位中排名第2(图2)。此外,通过对5个点位组别进行比较,9月份该地点的组之间的差异在统计学上不显著(F= 1.547,P= 0.239)。但是,LSD分析结果表明,各点位组别之间仍有一个显著差异组合:湿地对南门(P= 0.039)。
2.3 BOFP停落指数与环境特征之间相关性的时空动态
从奥森公园内蚊虫停落指数与环境因素不同时间段的线性回归分析发现,奥森公园各点位上蚊虫停落指数相关的环境因素也呈现出时间—空间尺度上的特异性。其中,7月的停落指数与400 m缓冲范围内有水草水体的面积(GW_400)正相关(系数= 2.816,P= 0.003),8月的停落指数与100 m缓冲区水体指数值(MDWI_100)负相关(系数= -0.003,P= 0.043),而9月的停落指数与100缓冲区内乔木的面积(HT_100)正相关(系数= 0.003,P= 0.004)。
3 讨论
3.1 奥森公园内蚊虫停落指数的空间尺度分布特征
本研究结果表明,同一个监测月份内,在奥森公园的不同点位之间的停落指数存在一些显著性差异点位组合,特别是湿地与其他点位之间显著性差异的机率更高(图2),这说明随着时间—空间的变化,不同点位的蚊虫停落指数是有显著性的时间—空间异质性。蚊虫繁殖是需要一定的时间—空间环境条件的,不同的环境条件决定了不同环境之间停落指数的多样性(Cumminsetal.,2012; 高强等, 2014);在合适的蚊虫繁殖环境下,蚊虫的停落指数可以始终保持很高水平(Smithetal., 2004),这可能是出现本研究结果的生态学原因。水体以及其中的植物是蚊虫幼虫高密度繁殖的良好条件(Govoetchanetal., 2014);本研究中,湿地里有丰富的水生植物,是很好的蚊虫繁殖地,所以奥林公园的湿地在不同调查时间窗口中都显示出很高的蚊虫停落指数;相反,由于植被稀少,生态系统简单的南门地区只能维持较低的蚊虫停落指数,这些都与蚊虫的生态特征非常吻合。为了达到最佳控制效果,不同的时间窗口与空间位置上需要制定特异化的预防和控制措施( Sogoba, 2007; 刘京利等, 2012; Prosperetal., 2012),而了解蚊虫停落指数的时间—空间特异性可以为制定这种特异化蚊虫控制策略提供支持。此外,了解蚊虫停落指数的时间—空间分布,也有助于在公园内指导人群在合适时间—空间上采取相应的保护措施,更好地降低人群受蚊虫叮咬的机率。最后,了解公园内蚊虫停落时间—空间分布的特异性,也可以指导科学用药,减少杀虫剂使用的盲目性,降低杀虫剂残留以及对环境的破坏。
3.2 奥森公园内蚊虫停落指数的时间尺度分布特征
研究发现,奥林公园内湿地在蚊虫停落指数上排名第一,而南门则是最低的(图3);而且在同一个监测点位上,不同月份间停落指数多数没有显著性差异;这说明奥森公园的蚊虫停落指数时间—空间分布模式主要体现在空间尺度上的变化。另一方面,在时间尺度上不管是总体的还是具体点位上的,蚊虫停落指数都是从7—9月呈现上升的趋势,在9月达到高峰。通常9月不是北京全市水平的蚊虫平均密度高峰,但本研究通过停落指数监测却发现其高峰期出现在9月,这与前期奥森公园的蚊虫密度CO2诱蚊灯监测结果一致(刘美德等,2019)。同一个城市中的不同环境区域中出现蚊虫密度动态不一致的现象已经被很多研究证实 (Smithetal., 2004; 高强 等, 2014), 而且这些研究也证实,这些差异性主要是归因于城市不同区域的环境特征。本研究发现,奥森公园蚊虫的停落指数密度表现出与全市平均水平不一致的现象是与奥森公园内特定的环境因素相关。深入对奥森公园内蚊虫停落指数与环境因素关系进行研究,可以帮助居民如何在奥森内合理选择活动时间与区域,以减少叮咬或传播疾病的风险。
3.3 奥森公园内蚊虫停落指数与环境因素的关系
蚊虫的停落指数受周边环境特征的影响 (Machaultetal., 2009)。本研究发现蚊虫停落指数与不同月份的特征环境动态有关,包括9月的乔木(HT_100),7月的长有植物水体(GW_400)和8月的水体指数(NDWI_100),这种现象可能与蚊幼虫高密度孳生的水体与植被特征相关(Jacupsetal., 2009; Kuruczetal., 2009)。同样,本研究证明,停落指数可被视为蚊虫繁殖的重要环境指示指标(Fondjoetal.,1992; Kigadyeetal., 2011; Zhuetal., 2015)。在本研究中,与停落指数相关的环境因素不断变化,这表明蚊虫停落指数受不同环境的影响。本研究结果可以科学客观地确定奥森公园需要处理的特定环境因素,以提高蚊虫防治的效率与准确性。这种有针对性的环境管理措施包括:间歇性灌溉生长有水生植物的水体;避免对公园内灌木丛、花卉和乔木的漫灌以减少蚊虫孳生地;可能的话,还可以对低矮灌木丛进行滞留喷洒,降低蚊虫繁殖或栖息的风险来降低蚊虫的停落指数。另一方面,本研究中的空间分析也表明蚊虫停落指数只与特定空间距离内的环境因素有关(例如:GW_400、NDWI_100和HT_100),这种相关的空间特征对于指导公共卫生部门确定公园内蚊虫孳生地处理空间范围具有积极的作用。
3.4 本研究的公共卫生意义
奥森公园对于维持北京城市生态系统的结构(连续性、生物多样性保护、物种及其栖息地保护)稳定性和城市生态安全性具有重要意义。研究奥森公园蚊虫停落叮咬的时空动态特征可以保护人们免受蚊虫的骚扰(Prosperetal., 2012),并降低蚊媒传播疾病的感染风险(Panetal., 2012)。此外,本研究也可以支持有针对性的生态环境治理,防止盲目地进行生态环境改造、更有效地发挥奥森公园作为北京重要生态屏障的功能。
猜你喜欢
矿山安全信息(2022年22期)2022-11-24
小学生学习指导(爆笑校园)(2022年9期)2022-09-30
保健医苑(2022年6期)2022-07-08
今日农业(2021年12期)2021-11-28
航天制造技术(2020年4期)2020-09-11
中国新闻周刊(2019年46期)2019-12-20
神州·上旬刊(2018年5期)2018-06-05
证券市场红周刊(2018年37期)2018-05-14
广东第二课堂·小学(2016年9期)2016-05-14
绿色科技(2014年3期)2014-07-11
