
北京综合性社区鸟类与植物的关系*
2022-05-10 03:28孙振凯韩丛海孙睿霖殷鲁秦姜莎莎
中国城市林业 2022年2期
徐 诗 王 成,2 孙振凯,2 韩丛海 孙睿霖,2 殷鲁秦,2 姜莎莎,2
1 中国林业科学研究院林业研究所 北京 100091 2 国家林业和草原局城市森林研究中心 北京 100091 3 北京市园林绿化局 北京 100013
随着城市化进程的不断加速,城市生态系统受到严重破坏,城市生物多样性水平持续下降[1]。 在创造人与自然协调的景观过程中,如何保护生物多样性是本世纪最大的保护类挑战之一[2]。 城市绿地在为城市野生动物提供合适的栖息地方面发挥着重要作用[3-5],而植物作为大多数野生动物赖以生存的物质环境,是维持城市生态系统自身平衡和保护生物多样性的重要元素。 因此,需要了解和把握城市不同种类野生动物的活动规律与植物的关系,并通过规划、设计和管理提高城市绿地的生物多样性[6-8],实现在有限的城市绿地内孕育出丰富物种的可能。
由于鸟类易调查、 易识别(与无脊椎动物或爬行动物等群体相比)、 种类繁多、 个体数量较大, 调查结果易统计分析, 许多关于城市化对生物多样性影响的研究, 都是利用鸟类来寻找影响城市野生动物种群分布和保护状况的因素[9-10]。因此, 城市鸟类不仅是评价某一地区生态环境质量的重要指标[11], 也被大多数学者视为研究城市生物多样性的最佳对象。
近年来, 城市鸟类与植物关系的研究范围仅限于城市公园和自然保护区[8,12-19], 鲜有在人类居住社区内的鸟类群落生态学研究。 Pyle[20]早在1978 年就提出, 社区间生物多样性差异是城市规划学者和动物保护科学家们需要高度关注的问题。如今在城市区域范围内, 各类社区已成为城市的重要组成部分, 对提升城市生物多样性有着举足轻重的作用。 生物多样性对人们(尤其是城市居民) 的生活质量至关重要[16], 合理地管理鸟类和其他野生动物可以抵消城市化带来的一些负面影响[2]。 因此, 对城市鸟类栖息场所的研究, 不应只局限于城市公园和城市周边的自然保护区。
Maeda 等[21]于1990—1991 年在日本东京一个居民区的12 个样点内进行繁殖和越冬季节的鸟类群落调查研究, 以确定与鸟类群落有关的生境特征, 结果发现植被尤其是乔木是保护居住区鸟类的关键因素。 郑光美[22]分别于1959、 1961—1964、 1973 年对北京及附近地区的夏季鸟类进行调查研究, 结果发现: 与公园鸟类组成相比, 居民点内的鸟类种类和数量相对匮乏, 鸟类种类组成单纯, 优势种数量多, 大型树栖鸟类罕见, 并未涉及到鸟类与植物的关系研究。 目前国内对于居住社区内鸟类与植物关系的研究近乎空白。 因此, 本文以北京市海淀区中国林业科学研究院社区为例, 从“哪些植物种类、 什么特征的植被形态, 以及何种结构的植物生境更加受鸟类喜爱”3 个方面, 对鸟类与植物的关系进行探究。
1 研究区概况
中国林业科学研究院社区(以下简称“林科院社区” ) 位于北京市海淀区香山路北侧(40°00′14″N, 116°14′27″E), 南望玉泉山, 属于典型的北温带半湿润大陆性季风气候, 冬季寒冷干燥, 夏季高温多雨, 平均海拔58 m, 占地面积26.3 hm2, 其中绿地面积10.3 hm2, 林木覆盖率高达48%。 林科院社区具有办公、 教育、 科研、居住等性质, 是比较典型的综合性社区, 院内绿地类型多样, 绿地空间众多, 植物种类丰富, 为鸟类提供了良好的栖息条件, 适宜开展小尺度城市社区鸟类与植物关系的研究。
2 研究方法
2.1 研究区空间类型划分及样线设置
根据植被类型、 生境、 功能等社区不同空间类型特点, 将林科院社区划分为28 个调查斑块, 设置一条串联所有斑块、 长度约3.4 km 的路线, 有序开展调查。 在每个斑块内各设置1 条鸟类调查样线, 确保样线覆盖调查斑块内的所有植被类型。
2.2 植物调查方法
采用全面调查法, 对林科院社区边界内的全部植物进行每木检尺调查、 量化测定其植被特征并确定其空间位置: 乔木记录树种名称、 数量、高度、 胸径、 冠幅、 枝下高; 灌木记录名称、 数量、 高度、 基径、 面积; 草本记录种名、 面积等;藤本记录种名、 面积等。
2.3 鸟类调查方法
利用样线法每2 周选择1 天晴朗天气进行鸟类调查, 全年共调查24 次。 每次调查1 天, 并通过系统调整整体路线上斑块调查的起始顺序, 消除调查时间差异带来的影响。 每次2 人, 以1~2 km·h-1的速度, 利用双筒望远镜、 数码相机和录音笔观测样线两侧鸟类情况, 记录观测的时间、 地点, 以及鸟的种类、 数量、 栖息树种、 行为特征、 所在的植被层, 调查者的鸟类鉴别技能来自北京市爱鸟协会野外调查训练以及《中国鸟类野外手册》 的学习。
2.4 植物冠层及生境划分方法
不同植物群落会对鸟类分布、 行为及栖息地选择产生一定的影响。 本研究权衡林科院社区28个斑块内现状绿地的不同空间位置、 乔灌草藤的比重, 以及植被的食源性、 观赏性和外部环境等因素, 综合垂直和水平两个方向, 按照不同植物群落空间层次和结构类型、 疏密类型对植物冠层及生境进行类型划分。
2.5 数据处理
根据Root[23]对集团的定义, 即以相似方式利用相同等级生境资源的一个类群, 本文提出9 个用以描述不同鸟类集团的变量: 虫食性、 杂食性、植食性、 乔木层、 灌木层、 地被层、 拾取、 探取、飞取。 然后用PCA 主成分分析法, 对记录到的和植物产生关系的29 种鸟类进行集团划分, 得到4个鸟类集团, 分别是第1 鸟类集团(乔木-虫食性集团)、 第2 鸟类集团(虫食-探取-乔木层集团)、 第3 鸟类集团(杂食性-拾取集团)、 第4鸟类集团(拾取集团)[24]。
用pearson 相关分析和偏相关分析方法, 分别对单株乔木高度(H)、 胸径(DBH)、 枝下高(BH)、 冠幅(C) 等植物形态特征指标和鸟类丰富度、 鸟类多度、 鸟类频数等鸟类活动特征指标进行相关性检验。 鸟类丰富度 (Bird richness,BR) 指单株乔木上的鸟类种类数量的累积值, 鸟类多度(Bird abundance, BA) 指单株乔木上的鸟类数量累积值, 鸟类频数 (Bird frequency,BF) 指鸟类落在单株乔木上的次数累积值。
由于林科院社区内不同植物种类在数量上存在明显差异,数量上有优势的植物种类可能会记录到更多的鸟类个体。 因此,为减弱树种数量差异带来的影响,在借鉴Lamont 等[25]物种重要值计算方法基础上,考虑植物数量对重要值的影响能够较为平等地对比不同树种对鸟类的重要程度[17]。 根据研究区域内所有乔木树种在28 个斑块内出现的频度,对其进行排序,将出现频度排名前20 的树种定义为常见植物,不同植物种类对鸟类群落的重要值为相对鸟类丰富度、相对鸟类多度、相对鸟类集团数三者之和与该种植株数的比例[17],如式(1)所示:
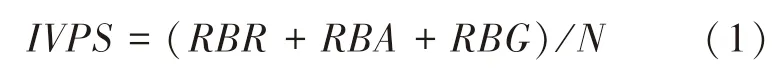
式(1) 中:IVPS为植物种类对鸟类群落的重要值;RBR为相对鸟类丰富度, 表示每个植物物种上观察到的鸟类物种数占常见植物上观察到鸟类物种数的比例;RBA为相对鸟类多度, 表示每个植物物种上观察到的鸟类数量占常见植物上鸟类数量的比例;RBG为相对鸟类集团数, 表示每个植物物种上鸟类集团数占常见植物上鸟类集团数的比例;N为该种植株数, 表示该植物在所有观测斑块中的种植总数量[17]。
同理, 植物生境对鸟类群落的重要值为相对鸟类丰富度、 相对鸟类多度、 相对鸟类集团数三者之和。 但是与IVPS不同, 计算植物生境对鸟类群落的重要值时, 相对丰富度为该植物生境下的鸟类物种数与全部鸟类物种数的比值, 相对多度为该植物生境下的鸟类数量与全部鸟类数量的比值, 相对鸟类集团数为该植物生境下的鸟类集团数与全部鸟类集团数的比值[17]。
3 结果与分析
3.1 林科院社区植物种类构成
本研究共记录到维管束植物71 科155 属212种(包含品种和变种)。 其中, 乔木97 种, 隶属39科65 属, 包括常绿针叶乔木19 种、 常绿阔叶乔木1种、 落叶阔叶乔木76 种、 落叶针叶乔木1 种; 灌木46 种, 隶属22 科38 属, 包括常绿针叶灌木3种、 常绿阔叶灌木7 种、 落叶阔叶灌木36 种; 草本植物75 种, 隶属于26 科65 属, 包括一二年生草本36 种、 多年生草本39 种; 藤本植物9 种, 包括草质藤本5 种、 木质藤本3 种、 攀援藤本1 种。
3.2 植物冠层及生境划分
本研究划分3 种结构类型、 6 种疏密类型和13种典型植物生境类型, 提取主要植物群落特征代表林科院社区整体植物群落生境风貌(表1), 进而从不同植物层次探讨鸟类对植物生境的偏好情况。

表1 林科院社区典型植物生境类型划分
3.3 不同植被层对鸟类活动的吸引力
城市鸟类与植物产生关系时发生的活动有停息、 取食、 营巢、 眺望与躲避天敌等, 同时, 不同的鸟类活动对不同类型的植物表现出不同的利用价值。 本次调查共观测到鸟类31 种5 967 只。乔木层共有71 种、 1 415 棵乔木累计出现29 种、3 914 只鸟, 分别占总数的93.55%和65.59%; 出现鸟类种数累积值前10 名的树种是雪松、 圆柏、银杏、 白皮松、 侧柏、 悬铃木、 油松、 枫杨、 水杉、 刺槐; 出现鸟类数量累积值前10 名的树种是圆柏、 白皮松、 柿树、 油松、 水杉、 刺槐、 雪松、银杏、 侧柏、 国槐。 灌木层共有30 种、 110 棵灌木累计出现8 种、 411 只鸟, 分别占总数的25.81%和6.89%; 出现鸟类种数累积值前5 名的树种是金银木、 丁香、 腊梅、 石榴、 大叶黄杨;出现鸟类数量累积值前5 名的树种是紫荆、 金银木、 大叶黄杨、 石榴、 卫矛。 草本层累计出现11种、 1 339 只 鸟, 分 别 占 总 数 的 35.48% 和22.44%。 藤本层中只有紫藤、 葡萄和猕猴桃上有落鸟, 累计出现3 种(麻雀、 白头鹎、 灰喜鹊)共175 只。 竹丛中共记录到3 种、 69 只鸟。 此外,还在菜地上记录到了麻雀、 灰喜鹊2 种59 只鸟。
通过分层统计落鸟类种数和个体数发现, 落鸟种数排序为乔木层>草本层>灌木层>藤本层=竹层>菜地, 落鸟个体数排序为乔木层>草本层>灌木层>藤本层>竹层>菜地。 可见, 乔木层鸟类种数和个体数都远远高于其他植被层, 占绝对优势; 其次为草本层和灌木层; 藤本层、 竹层和菜地出现种类数和个体数都比较少。
3.4 植物种类对鸟类活动的重要值
由表2 可知, 在乔木层中, 林科院社区鸟类群落对泡桐的喜好度最高, 重要值为0.036。 排名前10 的树种依次为泡桐、 水杉、 加杨、 栾树、国槐、 臭椿、 刺槐、 雪松、 玉兰、 华山松, 其中仅水杉、 刺槐和雪松同时出现在鸟类种类和数量累计前10 的乔木中, 且株数也在20 个常见乔木种中均偏小, 但对鸟类的重要性排名靠前。
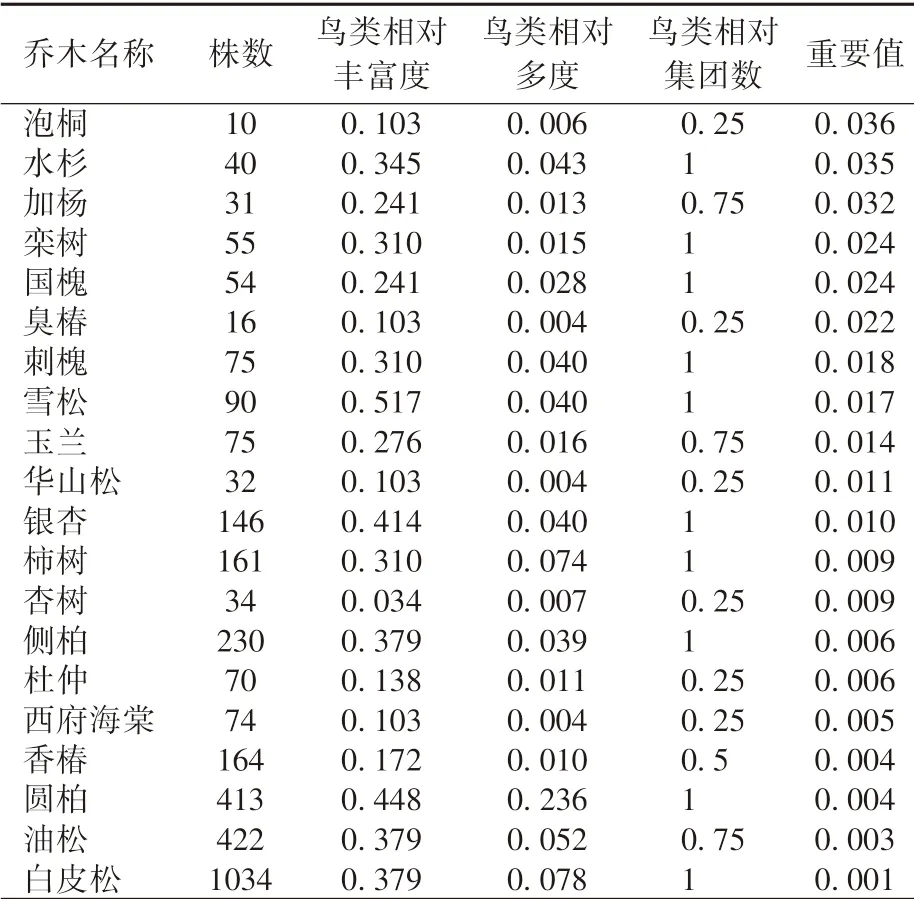
表2 林科院社区20 种常见乔木对鸟类群落的重要值
3.5 鸟类活动与乔木形态特征的关系
通过对植物形态特征值中的乔木高度、 胸径、枝下高、 冠幅4 个因子做pearson 相关分析发现,4 个变量因子两两之间存在显著(P<0.05) 正相关关系。 为了消除4 个植物形态特征因子之间的相互影响, 本文采用偏相关分析方法, 在控制其他植物形态特征因子影响的前提下, 观察鸟类特征变量是否受到乔木形态特征变量的影响, 结果(表3) 表明, 乔木高度和鸟类丰富度、 鸟类多度、 鸟类频次呈显著(P<0.05) 正相关关系, 即乔木越高, 吸引前来栖息的鸟类的种类数、 个体数和落脚次数就越多。 乔木冠幅和鸟类丰富度呈显著(P<0.05) 正相关关系, 即乔木冠幅越大,吸引的鸟类种类数就越多。 胸径和枝下高对鸟类活动特征无显著影响。

表3 偏相关分析鸟类活动与植物形态特征值的相关系数
3.6 不同植物生境对鸟类集团的吸引力
由表4 可知: 本研究划分的4 个鸟类集团都出现过的植物生境有柿子林、 油松林、 疏林、 杂木园1、 杂木园2、 庭院林1、 庭院林2; 雪松林、月季园未曾出现第2 集团鸟类; 白皮松林和乔松林未曾出现第2、 4 集团鸟类; 温室小游园、 庭院林3 未曾出现第4 集团鸟类。 庭院林1 中4 个鸟类集团的物种数都达到最大值, 分别是6、 3、 4、4 种。 可见, 在13 个植物生境中, 庭院林1 对4个鸟类集团的吸引力最强。

表4 林科院社区13 个植物生境中的鸟类集团及物种
3.7 不同植物生境对鸟类的重要值
13 种植物生境对鸟类的重要值排序为庭院林1>庭院林2>油松林>疏林>杂木园2>柿子林>杂木园1>雪松林>温室小游园>庭院林3>月季园>白皮松林>乔松林(表5)。 庭院林1 和2 排序靠前,可能是因为两者面积比其他生境均偏大(表1),从而导致记录到的物种和数量相对较多。 因此,13 个生境对鸟类的重要值排序只作为背景值, 以反映林科院社区不同生境在空间上对鸟类重要性的差异现状, 本文仅对相同植物群落结构和疏密类型的植物生境面积大小相似的生境进行对比分析。
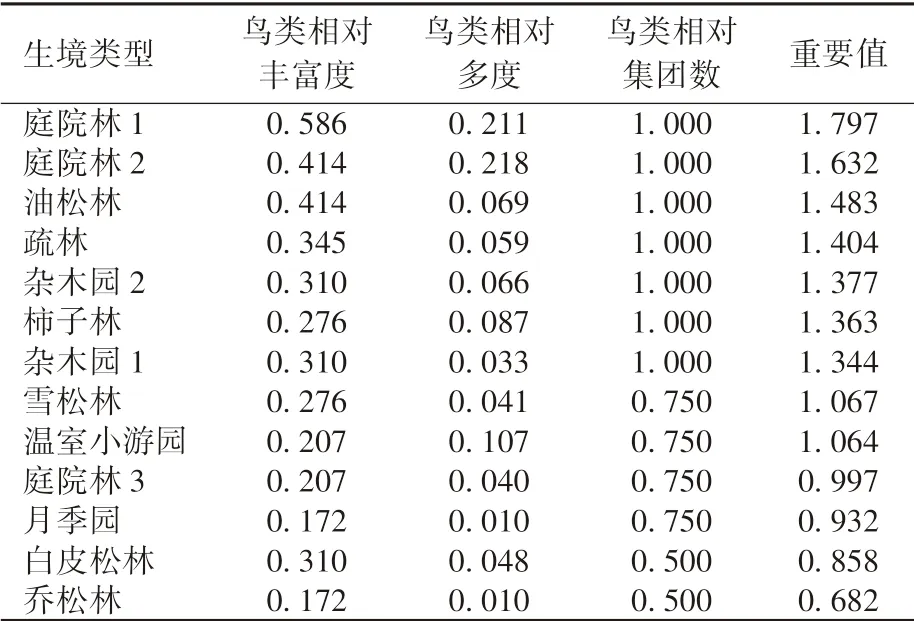
表5 13 个植物生境对鸟类活动的重要值
在4 种乔草类群落—密林草地型植物生境中,以油松为主的密林对鸟类的重要性最大, 以柿树为主的密林次之, 之后是以雪松为主的密林, 以白皮松为主的密林对鸟类的重要性最低。 在2 种乔草类群落—疏林草地型植物生境中, 疏林比乔松林对鸟类更具有吸引力。 在2 种乔灌草类群落—密林灌草型植物生境中, 杂木园1、 杂木园2对鸟类的吸引力程度相似, 考虑到两者的面积均小于前4 名(庭院林1、 庭院林2、 油松林和疏林) 的面积, 其总体上对鸟类的重要性都比较高。 在3 种乔灌草类群落—疏林果蔬型植物生境中, 庭院林1、 庭院林2 和庭院林3 都是位于居民楼宅旁和楼间的组团绿地, 均为房前屋后的庭院式镶嵌种植模式, 主要乔木树种有柿树、 香椿、圆柏等, 乔灌草搭配丰富, 食源树种多样, 为不同鸟类提供多种食物来源。 庭院林1 和2 面积差别不大, 重要值相近, 但庭院林3 面积约为前两者的一半, 重要值也减小一半左右, 说明庭院林的生境斑块面积越大, 对鸟类的重要值就越大。
4 讨论
4.1 不同植被层对鸟类的吸引力
乔木和灌木是鸟类栖息地中重要的生态因子[12,26], 其群落结构和物种组成直接影响鸟类栖息地[16,27]。 林科院社区乔木种类占比43%, 乔木层对鸟类的吸引力远大于其他植被层, 作为社区植被群落结构的骨架, 对鸟类的生存起到至关重要的作用。 调查发现, 大嘴乌鸦、 灰喜鹊等经常立于乔木顶端, 以眺望、 寻找食物, 而大部分种类生性胆小, 常藏于乔木上层树冠内活动。 虽然有研究表明灌木层的丰富度对鸟类物种多样性有积极影响[12,28-29], 林下灌木为鸟类提供更多的觅食机会和繁殖资源[30], 灌木配置的复杂程度为鸟类提供更多隐蔽空间, 但是本研究发现林科院社区的灌木层对鸟类种类和数量的吸引程度低于乔木层和草本层。 这可能是因为鸟类大多数在草本层觅食或在乔木层停歇、 觅食、 避险和筑巢, 而灌木层则因为高度和食物不足导致吸引力降低。未来研究将进一步分析鸟类行为特征、 鸟类集团数量与灌木层的关系, 以探究社区中灌木层对鸟类活动的意义。
社区内草本层栖息鸟类数量和种类仅次于乔木层, 成为鸟类活动的重要空间, 对整个社区鸟类群落构成有重要意义。 调查发现: 灰椋鸟、 珠颈斑鸠、 乌鸫等偏爱在草地活动; 麻雀、 珠颈斑鸠等会捡拾草籽以及乔木尤其是圆柏、 侧柏等落下的种子; 乌鸫、 灰椋鸟等虫食性鸟类会在草地上行走, 觅食叶片上的虫类, 而且在寒冷冬季昆虫不足的情况下还会捡拾地被中各种植物种子。单一的人工草坪不能满足鸟类在草坪觅食的需求,而杂草中的草籽、 花芽以及落在草丛内的乔木果实等可作为植草性鸟类的食物。
4.2 鸟类多样性与植物种类的关系
有研究表明多样的植被对鸟类物种多样性有积极的影响[31]。 最具吸引力的食物来源是吸引昆虫的植物物种, 其次是生有花蜜、 水果和谷物的树种, 而乡土植物种类能够为鸟类提供全年的食物资源和更好的庇护所[32]。 本研究发现, 泡桐对林科院社区鸟类的吸引力最大, 原因可能是泡桐植株个体高大、 小枝粗壮、 冠幅和叶片面积相对较大, 且多为孤植、 树形舒展, 从而使鸟类的落脚面相对较大并能够为鸟类提供必要的隐蔽条件。但在泡桐上驻足活动的鸟类集团只有以喜鹊为代表的第3 集团, 而在水杉和雪松上4 个集团的鸟类均有出现, 并且位于鸟类数量和种类排名前8,表明水杉和雪松对鸟类活动表现出一定的重要性。栾树、 国槐、 臭椿作为乡土植物, 其重要值在20种常见乔木中排名前6, 这也进一步说明了乡土植被对城市鸟类的重要意义。 另外, 研究区还有26 种树种未观测到鸟类栖息活动, 很大程度上可能是因为这些树种作为科研引种或景观树种, 数量稀少, 在观测中未能记录到鸟类活动, 但并不代表这些树种对鸟类没有吸引力, 需要今后更多观察研究。
4.3 鸟类多样性与植物形态特征的关系
植被形态特征是影响鸟类栖息地质量最为重要的因子, 如植被结构、 高度、 盖度等与城市鸟类丰富度关系密切[15,33-34], 尤其是对小尺度的鸟类栖息地[35]。 有研究表明, 高大的植被可以为鸟类提供丰富的食物资源和隐身空间[36], 大型针叶树对于增加鸟类的物种数量有显著作用[15]。 本研究发现: 乔木高度在控制或增加鸟类种类和数量等方面具有重要作用; 乔木冠幅对增加鸟类物种数有显著影响, 对增加鸟类数量无影响; 乔木胸径对鸟类种类和数量的变化均无影响。 因此, 拥有大树冠的高大乔木更能够创造出适合不同生活习性鸟类的多个生态位, 进而吸引多种不同的鸟类前来栖息。
4.4 鸟类多样性与植物生境的关系
在绿地设计中, 异质多样的复合生境可以为野生生物营造丰富多样的栖息地[37], 生境结构和复杂性是决定鸟类群落多样性的两个主要因素[38-39]。 本研究发现不同植物生境可以容纳不同种类和数量的鸟类: 比如黄腰柳莺、 金翅雀只出现在油松林中; 大斑啄木鸟只出现在庭院林1 中;在同样群落结构的油松林、 柿子林、 雪松林和白皮松林中, 油松林的鸟类相对丰富度和相对多度较高, 而雪松林和白皮松林吸引较多第1 集团鸟类。 整体来看, 相同疏密类型的北京社区森林环境中, 一定面积的庭院林、 油松林、 柿子林以及乔灌草搭配种植的多树种混交林对鸟类的重要性较高, 鸟类更加偏好多树种、 多层次、 多结构的近自然结构林。
5 结论
本文从鸟类与植物种类、 植物形态特征、 植物生境3 方面的关系探究林科院社区城市鸟类活动与植物的关系, 结论如下:
1) 社区内乔木层观测到鸟类种类和数量最多, 其次为草本层和灌木层。
2) 不同乔木对鸟类的吸引力不同, 树体高、冠幅大的乔木会吸引更多的鸟类。 林科院社区常见乔木对鸟类群落的重要值排序为泡桐>水杉>加杨>栾树>国槐>臭椿>刺槐>雪松>玉兰>华山松。
3) 不同植物生境类型中出现的鸟类差异较大。 这表明通过合理的植物选择和配置可以吸引不同种类的鸟类, 人类活动频繁的庭院林地带也可以成为鸟类重要的生活空间, 这在增加社区鸟类多样性方面具有巨大的潜力。
4) 树种单一、 绿地结构简单的人工纯林对鸟类的吸引力较低。 多种植物混交、 多层次结构配置、 拥有整体近自然林、 局部空间相对开敞、 植物生长舒展的社区森林对保护和提高社区鸟类多样性更有吸引力。
5) 以高大乔木为主、 具有多种生境类型的社区有利于吸引和承载更多的鸟类活动。
猜你喜欢
中国农村水利水电(2021年11期)2021-11-29
风景园林(2021年2期)2021-04-24
落叶果树(2021年6期)2021-02-12
现代园艺(2020年18期)2020-12-22
农村实用技术(2020年1期)2020-12-16
智富时代(2019年8期)2019-09-23
智富时代(2019年8期)2019-09-23
小天使·一年级语数英综合(2019年8期)2019-08-27
读写算·小学中年级版(2017年4期)2017-05-08
小学阅读指南·低年级版(2016年6期)2016-05-14
