
不同类型森林植被群落多样性与土壤养分的关联分析
2022-05-10 12:02张栋
山东农业大学学报(自然科学版) 2022年2期
张栋
不同类型森林植被群落多样性与土壤养分的关联分析
张栋
河南农业职业学院, 河南 郑州 451450
为了探究不同植被类型群落分布多样性与土壤养分之间的关系,本研究将琅琊山不同林木作为研究对象,所选择林木不仅包括天然次生林和针叶林,还包括阔叶林及人工林,为了进一步降低实验误差,本研究从2013年开始进行为期五年的持续性实验,通过实验对比得知:(1)无论是次生林和针叶林,还是阔叶林及人工林,虽然其群落分布特点存在较大的差异,但是其整体的群落变化态势较为一致,无论是丰度、匀度指数方面,还是Wiener多样性指数方面,最高的属于次生林,而最低的则是针叶林;在Wiener多样性方面,不同林木类型之间的差异尤为突出,且通过了0.05显著检验。在Simpson指数方面,最高的则是针叶林,其次是人工林,而次生林最低,这与其他多样性指数恰好相反;(2)在Cody指数方面,最高的是次生林,其次是阔叶林,而针叶林最低,前两者之间的差异并不突出,且未通过显著检验,但与后两者的差异较为突出,且通过了0.05显著检验。对于Sorenson指数来讲,最高的是针叶林,其次是人工林,而次生林最低,不同林木类型之间的差异并不突出,未通过显著检验;(3)在土壤方面,无论是有机碳、磷等养分,还是微生物量,随着深度增加,其下降态势尤为突出,且这种减小幅度较为突出的是在20 cm内的表层土壤,而在80~100 cm的土层则幅度微小,对于同一土层而言,次生林含量最高,其次是阔叶林,而针叶林最低;(4)通过交互因子分析得知,林木多样性分布不仅受土壤深度制约,还受到林型差异的影响,且达到了0.05显著水平,尤其是林型和深度的交互作用更为突出;(5)通过开展冗余分析得知,土壤养分状况直接制约着林木群落分布多样性,成为关键性的作用因子。
森林; 植被多样性; 土壤养分
全球生态系统包括大气、陆地、海洋等,这些是关键的生态系统构成,除此之外,土壤、森林及山地高原等多种生态因子也是不可或缺的构成部分[1-3],整体而言,生态系统具有复杂的多样性[4],此外,其还体现着突出的复杂性,且多种环境因子之间会产生明显的交互作用,受此影响之下形成了复杂的生态局地环境[5,6];对于土壤而言,作为农作物等植被生长的养分及水分供应载体,直接关乎人类粮食作物及蔬菜种植等[7],制约居民生活状况,否则作物难以获取必要的水分及养分维持生长,对于土壤和植被而言,其在陆地生态的地位难以替代,贫瘠的土壤将难以承载作物等植被的生长发育[8,9],土壤质量与微生物状况紧密相连,同时与酶的关系密切,微生物能够有效地进行有机质降解,在酶的参与作用下,形成高效的养分转化,进而维持土壤肥力及活性,为植被生长提供所需能量。对于土壤而言,其有机碳库在陆地碳库方面作用显著,制约着生物多样性及土壤养分状况。植被根系对于固土、保持土壤通透性至关重要,在根系及其分泌物作用下,土壤颗粒能够对于孔隙度有利,同时对于植被生长有利。森林生态在陆地生态中尤为关键,能够有效地调节局地气候,在涵养水源方面效果突出,探究其植被、土壤的关系,能够对森林生态加以分析,对其土壤质量、植被分布群落特点等进行对比分析,从而探究其复杂的作用机理,从微气候、土壤、动植物的角度探究森林生态,并对不同土壤质量下的植被分布加以分析,对多种环境因子及其交互效应对植被的影响进行探究。对于不同的植被分布区域,其土壤养分的时空分布一般呈现较大差异,大量学者对此进行了研究,并进行了实证分析,植被和土壤之间的作用机理较为复杂。
对于安徽琅琊山而言,作为国家重要的森林公园,其生物多样性尤为突出,这里分布着广泛的热带森林[10],对于森林而言,其不仅分布着大量的植被,还分布着大量的生物,尤其是微生物分布较为普遍,微生物不仅是土壤生态的关键构成,在有机质分解方面发挥着无可替代的作用,加之酶的参与,直接制约着整个土壤养分循环和能量交换,进而影响着植被生长发育。不同的森林分布,使得植被的多样性分布也出现较大差异。对于琅琊山而言,其不仅分布着大量的针、阔叶林,还包括大量的天然次生林,为了森林资源保护,还进行了大量的人工林建设,为了探究不同森林资源下的植被群落分布,本研究将以上森林分布作为研究对象,从植被及土壤的角度探究其群落分布,并从时间、空间的角度加以分析,从而为森林资源的合理开发利用提供数据参考。
1 材料和方法
1.1 研究区
本研究所选择的区域位于安徽琅琊山,该区域是国家重要的森林公园,紧邻滁州市西南,为淮阳山脉的丘陵区域,海拔在300 m左右,受置于所在地理分布,其亚热带季风气候特点尤为突出,无论是降雨还是气温,其季节变化尤为突出,对于夏季而言,其不仅具有近1 400 mm的较高降雨量,且主要集中在四至六月,同时具有较高的气温,根据近年来该区域气象资料分析得知,其年均温达到15 ℃,最高温在7月份,其月均温达到29 ℃;对于冬季而言,其不仅气温较低,且天气干燥,最低温集中在1月,均温不足5 ℃。对于该区域而言,土层较浅,一般不超过五十厘米,其pH值低值一般为6.5,而高值达到8,呈现微碱性或者中性,其肥力并不高,甚至不少区域出现了明显的岩石裸露。对于该区域而言,其主要的土质不仅包括黄棕壤,还有褐色土、石灰岩土,整体而言,其土层并不厚。不仅分布着大量的针、阔叶林,还包括大量的天然次生林,为了森林资源保护,还进行了大量的人工林建设。
1.2 物种多样性的测度
本研究采取实证分析的方法进行连续的观测研究,正式的实验开始于2013年,为了最大程度降低实验误差,本研究实验持续五年,选择的林木类型不仅包括针、阔叶林,还包括大量天然次生林、人工林。对于各个林木类型进行5个样地设置,且长、宽均为20 m,从而形成样地20个;接下来还需要进行样方的设置,对于乔木和灌木样方而言,数量各4个,其长、宽均为10 m;对于草本样方而言各8个,其长、宽均为1 m;合计实验样方240个。并对生境及长势进行记录。
主要从以下方面对植被多样性加以分析,计算如下[11-13]:
Wiener指数=-∑(PlnP)
Margalef指数=(-1)/ln
Simpson指数=1-∑(P)2
主要从以下方面对植被β多样性加以分析,计算如下:
Cody指数()=(+)/2
Sorenson指数()=2/(+)
1.3 土壤养分和微生物量的测定
土壤取样:本研究利用钻土取样,取样间隔为20 cm,直至土壤100 cm深度,为有效降低误差,借助于3次重复。然后对其过筛风干处理,对水分、有机质测定通过烘干法、加热法,微生物量碳、氮用浸提法测定[9-12]。
土壤微生物周转估算方法如下[14,15]:
=Σ
=Σ/
=1/
=××/
利用Excel 2010软件进行数据的预处理后,用SPSS 21.0进行均值间的差异性检验。
2 结果与分析
2.1 不同森林类型α多样性
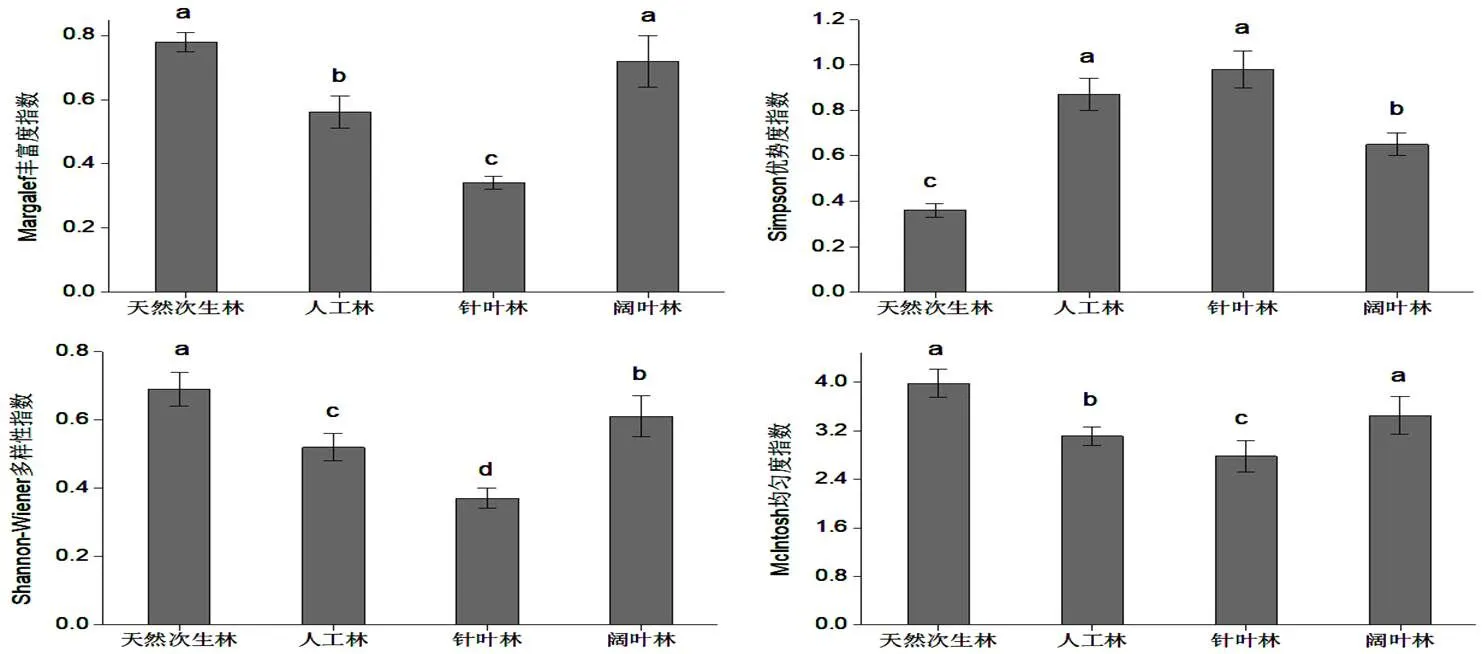
图 1 不同林型α多样性
注:不同小写字母表示差异显著(<0.05),下同。
Note: Different lowercase letters indicate significant differences (<0.05), the same as below.
通过对图1的分析不难得知,无论是阔叶林和天然次生林,还是针叶林和人工林,虽然其群落分布特点存在较大的差异,但是其整体的群落变化态势较为一致,无论是丰度、匀度指数方面,还是Wiener多样性指数方面,其最高的是次生林,而针叶林最低;前者之间的差异并不突出,而后者较为突出,且通过了0.05显著检验。在Wiener多样性方面,不同林木类型之间的差异尤为突出,且通过了0.05显著检验;Simpson指数大致:天然次生林<阔叶林<人工林<针叶林,其中人工林和针叶林差异不显著(>0.05),二者显著高于天然次生林和阔叶林(<0.05)。
2.2 不同森林类型β多样性
通过对图2的分析得知,在Cody指数方面,最高的是次生林,而针叶林最低,前两者之间的差异并不突出,且未通过显著检验,但与后两者的差异较为突出,且通过了0.05显著检验。从Sorenson指数来讲,针叶林最高,其次是人工林,而次生林最低,不同林木类型之间的差异并不突出,未通过显著检验。总之,天然次生林和阔叶林相似性较大,表明阔叶林和天然次生林共有物种数较多。
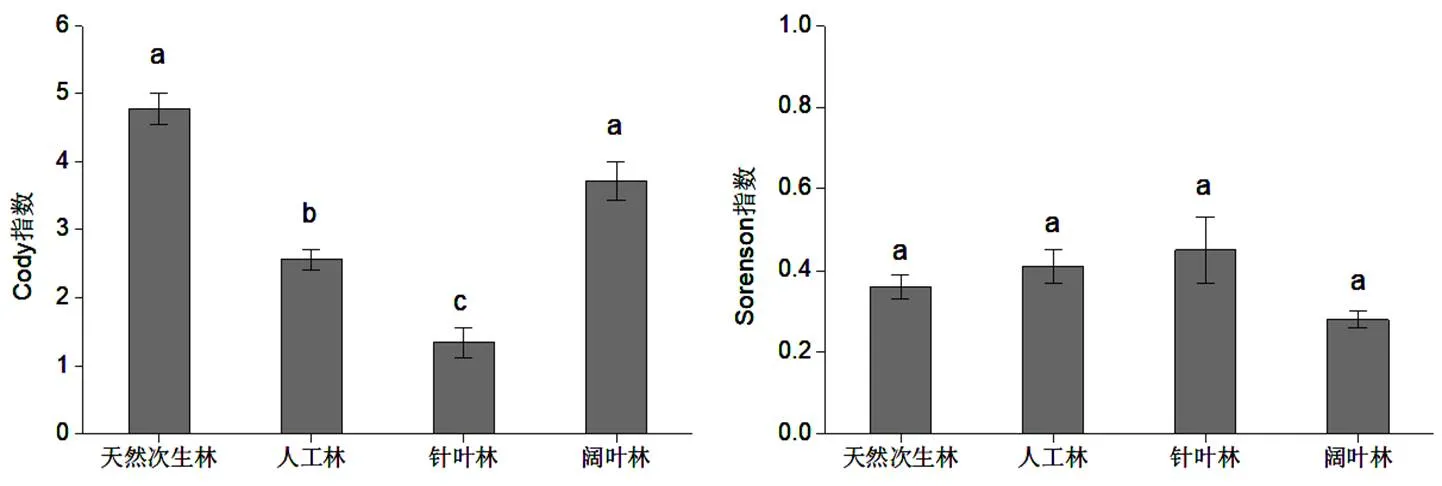
图 2 不同林型β多样性
2.3 不同森林类型土壤养分特征
由图3可知,不同森林土壤养分(全氮、有机碳、全钾)随土层深度而减小,40 cm土层以下急剧减小,80~100 cm土壤养分含量基本一致;相同土层土壤养分大致表现为:天然次生林>阔叶林>人工林>针叶林,深层80~100 cm差异并不大。

图 3 不同森林类型土壤养分变化特征
2.4 不同森林类型土壤微生物量特征
通过对图4分析发现,虽然林木类型不同,但是对于微生物量碳及氮磷而言,在土层深度增加的情况下,其含量下降态势尤为突出,尤其是在40 cm以内的表层土层。80~100 cm土壤微生物量基本一致;相同土层土壤微生物量大致表现为:天然次生林>阔叶林>人工林>针叶林,深层80~100 cm差异并不大。

图 4 不同森林类型土壤微生物量变化特征
2.5 不同森林类型土壤微生物周转
由图5可知,不同森林类型土壤微生物量碳转移量和周转率随随土层深度的增加而减小,其含量下降态势尤为突出,尤其是在40 cm以内的表层土层,而在80~100 cm的土层则幅度微小;相同土层土壤微生物量碳转移量和周转率大致表现为:天然次生林>阔叶林>人工林>针叶林,深层80~100 cm差异并不大。

图 5 不同森林类型土壤微生物周转
2.6 不同森林类型群落多样性与土壤养分的关系
从表1的交互分析得知,林木多样性分布不仅受土壤深度制约,还受到林型差异的影响,且达到了0.05显著水平,尤其是林型和深度的交互作用更为突出。从表2可以看出,对于土壤养分而言,其不仅受土壤深度制约,还受到林型差异的影响,且达到了0.05显著水平,尤其是林型和深度的交互作用更为突出,但土层深度的制约效应并不突出。
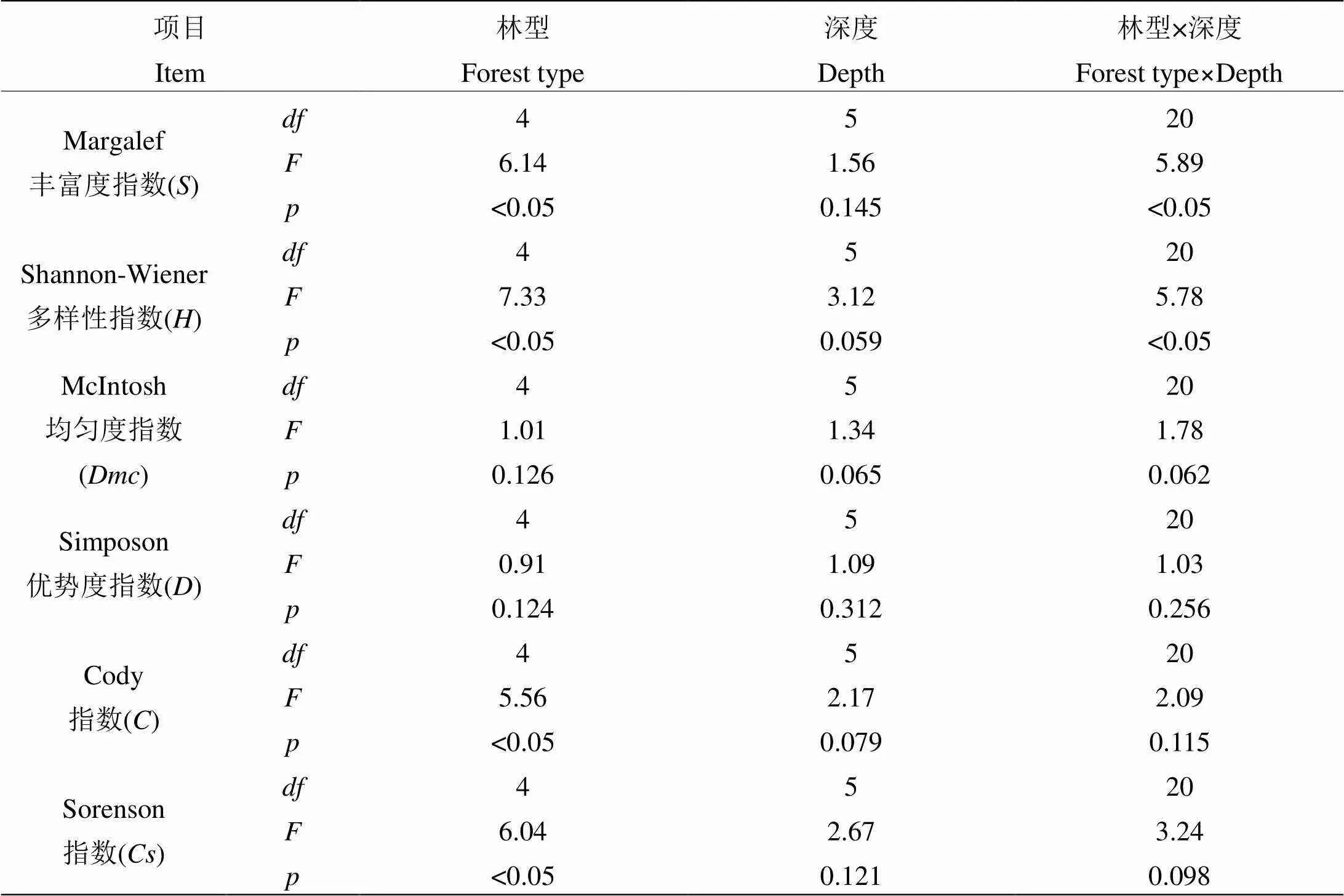
表 1 土壤深度和林型对植被多样性的影响

表 2 土壤深度和林型对土壤养分的影响
植被群落分布受到的影响因子并不是单一的,且不同影响因子之间存在交互作用,本研究从多个角度探究养分等与植被之间的关系,特进行了冗余分析,其中的响应及解释变量分别为群落多样性、土壤养分,从而直观的分析多种影响因子的制约效应。通过对图6可知,对植被群落分布多样性产生更为突出影响的是第一个因子,因子解释度超过了60%,而对于前两个排序轴影响因子而言,其对变量的累积解释度超过了80%,这说明这些环境因子对于植被多样性分布产生了尤为突出的制约效应。通过长期的观测研究得知,无论是全磷,还是微生物量磷,均对植被多样性施加突出制约效应,通过相关分析得知,在植被多样性更为突出的情况下,土壤的养分含量同步变化。
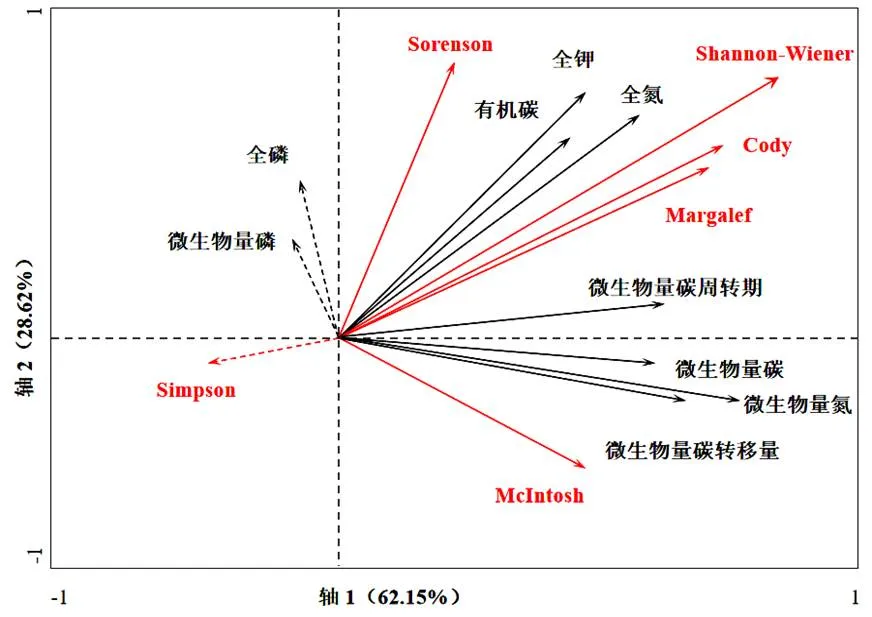
图 6 群落多样性与土壤养分的RDA分析
3 讨 论
对于微生物新陈代谢而言,一方面受制于土壤养分状况,这是直接的作用因子,另一方面依赖于湿热条件,这是其外部环境因子,这些对于微生物产生直接影响,决定着其活动效率,进而制约着有机质分解速率,对于土壤质量施加着无可替代的影响。微生物在养分循环及能量交换过程中的作用尤为突出,加之其处于不断动态变化之中,因此,对于植被的影响也是处于不断变化之中。此外,雨热等环境因子也对植被施加直接的制约效应[16-18]。通过连续实验数据分析得知,对于不同林木多样性来说[19,20],其在Shannon多样性、Margalef丰度、均匀度方面的差异较为突出,这主要在于林木类型差异的制约,但是在时间不断增加的情况下,其上升态势尤为突出,指数最高的是次生林,而最低的则是针叶林。且通过了0.05的显著性检验;对于Simpson指数来讲与之相反。对于Wiener多样性而言,不同林木间的差异尤为突出,且通过了0.05的显著性检验。整体而言,对于阔叶林和次生林而言,其生境因子更为复杂,且具有较高的多样性,这也是自然环境及人为影响综合作用的结果。
通过连续的观测研究不难得知,虽然林木类型的差异对于土壤养分及微生物量碳产生着一定的影响,形成了不同的含量差异,但是其具有较为接近的变化态势,其含量均在土层不断增加的情况下呈现较为突出的下降态势,尤其是40 cm土层以下急剧下降,而80~100 cm土层其含量并无多大差别[3,4];对于同一土层来讲,含量最高的是次生林,其次是阔叶林,而针叶林最低;对于氮、磷的含量变化来讲亦是如此,对于微生物量碳周转率和周转期具有同样的变化特点。对于次生林和阔叶林而言,其较多的枯枝落叶能够带来更多的腐殖质,为有机质的增加奠定基础。通过交互因子分析得知,森林群落多样性不仅受制于林型及土层的制约,还受到其交互因子的制约,且通过了0.05的显著性检验;对于和多样性来讲,土层深度对之并没有显著制约[17-19];土壤养分一方面受制于林木类型的制约,另一方面受土层深度影响。
植被在生长过程中受到多种环境因子的制约,且其交互作用更为复杂,对于多样性群落分布来说亦是如此,且多种环境因子之间会产生明显的交互作用影响。相应的,植被也会调节局地环境因子[21,22]。
[1] 李胜平,王克林.桂西北喀斯特山地草地土壤养分季节变化规律及其对植被多样性的响应[J].水土保持学报,2016,30(4):199-205
[2] 胡芳,杜虎,曾馥平,等.典型喀斯特峰丛洼地不同植被恢复对土壤养分含量和微生物多样性的影响[J].生态学报,2018,38(6):201-211
[3] 黄庆阳,曹宏杰,王立民,等.五大连池火山熔岩台地植物多样性与土壤养分的关系[J].浙江农林大学学报,2019,36(1):80-87
[4] 刘洋,侯占山,赵爽,等.太行山片麻岩山区造地边坡植被恢复过程中植物多样性与土壤特性的演变[J].生态学报,2018,38(15):5331-5339
[5] 王莹,庞晓攀,肖玉,等.高原鼠兔干扰对高寒草甸植物多样性与土壤养分间关系的影响[J].生态学报,2016,36(17):235-241
[6] 李秀清,李晓红.鄱阳湖湿地不同植物群落土壤养分及微生物多样性研究[J].生态环境学报,2019,28(2):385-394
[7] 姜勇,李天鹏,冯雪,等.外源硫输入对草地土壤-植物系统养分有效性的影响[J].生态学杂志,2019,38(4):1192-1201
[8] 杨秉珣,刘泉,王彬.嘉陵江流域不同类型植被多样性与土壤养分和酶活性的关系[J].水土保持研究,2016,23(6):45-51
[9] 刘娜,白可喻,杨云卉,等.放牧对内蒙古荒漠草原草地植被及土壤养分的影响[J].草业科学,2018,5(6):1323-1331
[10] 宋贤冲,郭丽梅,田红灯,等.猫儿山不同海拔植被带土壤微生物群落功能多样性[J].生态学报,2017,37(16):1124-1129
[11] 宁志英,李玉霖,杨红玲,等.沙化草地土壤碳氮磷化学计量特征及其对植被生产力和多样性的影响[J].生态学 报,2019,39(10):3537-3546
[12] 文东新,杨宁,杨满元.衡阳紫色土丘陵坡地植被恢复对土壤微生物功能多样性的影响[J].应用生态学报,2016,27(8):2645-2654
[13] 申家琛,张朝晖,王智慧.石漠化程度对苔藓植物多样性及其结皮土壤化学性质的影响[J].生态学报,2018,38(17):6043-6054
[14] 杨媛媛,陈奇伯,黎建强,等.滇中高原华山松植物多样性与土壤生物肥力特征[J].土壤,2017,49(1):90-96
[15] 孙千惠,吴霞,王媚臻,等.林分密度对马尾松林林下物种多样性和土壤理化性质的影响[J].应用生态学报,2018,29(3):732-738
[16] 薛超玉,焦峰,张海东,等.黄土丘陵区弃耕地恢复过程中土壤与植物恢复特征[J].草业科学,2016,33(3):368-376
[17] 张静,温仲明,李鸣雷,等.外来物种刺槐对土壤微生物功能多样性的影响[J].生态学报,2018,38(14):4964-4974
[18] 梁香寒,张克斌,乔厦.半干旱黄土区柠条林土壤水分和养分与群落多样性关系[J].生态环境学报,2019,28(9):1748-1756
[19] 张变华,靳东升,张强,等.不同植物种植对矿区复垦土壤微生物多样性的影响[J].农业资源与环境学报,2019,36(3):355-360
[20] 韩煜,王琦,赵伟,等.草原区露天煤矿开采对土壤性质和植物群落的影响[J].生态学杂志,2019,38(11):3425-3433
[21] 彭东海,侯晓龙,何宗明,等.金尾矿废弃地不同植被恢复阶段物种多样性与土壤特性的演变[J].水土保持学报,2016,30(1):159-164
[22] 金章利,刘高鹏,周明涛,等.喀斯特山地草地群落多样性海拔特征及土壤理化性质特征[J].生态环境学报,2019,28(4):661-668
Correlation Analysis of Vegetation Community Diversity and Soil Nutrients in Different Forest Types
ZHANG Dong
451450,
Different types of forest vegetation communities (natural secondary forest, broad-leaved forest, artificial forest and coniferous forest) were selected from langya mountain forest park in anhui province to study the relationship between diversity and soil nutrients for 5 consecutive years (2013-2017). Results show that: (1) of natural secondary forest and plantation, coniferous and broad-leaved forest, the species diversity variation law of each index showed a consistent Margalef abundance index, Mclntosh species evenness index and Shannon Wiener diversity index were as follows: the natural secondary forest > broad-leaved forest plantation > coniferous forest, the natural secondary forest and broad-leaved forest Mclntosh evenness index and Margalef abundance index difference was not significant (> 0.05), significantly higher than both plantation and coniferous forest (<0.05); The shannon-wiener diversity index of different forest types was significantly different (<0.05). Simpson dominance index shows that natural secondary forest < broad-leaved forest < artificial forest < coniferous forest;In general, the natural secondary forest and broad-leaved forest are more complex, and the plant diversity index is higher than that of other habitat communities. (2) the Cody index was as follows: natural secondary forest > broad-leaved forest > artificial forest > coniferous forest, among which there was no significant difference in Cody index between natural secondary forest and broad-leaved forest (>0.05), which was significantly higher than artificial forest and coniferous forest (<0.05). The Sorenson index was successively shown as natural secondary forest < broadleaf forest < artificial forest < coniferous forest, and there was no significant difference in Sorenson index among different forest types (>0.05). (3) different forest types of soil nutrients (organic carbon, total nitrogen, total phosphorus and total potassium content), microbial biomass carbon, nitrogen, phosphorus and microbial biomass carbon turnover rate and turnover period decreasing trend with the increase of soil depth, among them, 20-40 cm soil layer under sharply reduce the trend, 80-100 cm difference is not obvious, the basic trend of equal; The same soil layer is roughly manifested as: natural secondary forest > broad-leaved forest > artificial forest > coniferous forest. (4) two-factor interaction analysis showed that soil depth and forest type had a significant impact on forest community diversity (<0.05), in which soil depth had no significant impact on and diversity index (>0.05), and forest type and diversity index had a significant impact on and diversity index (<0.05). Soil depth and forest type had significant effects on soil nutrients (<0.05), in which soil depth had no significant effect on soil nutrients (>0.05), and forest type and forest type × depth had significant effects on soil nutrients (<0.05). (5) the results of redundancy analysis showed that soil nutrients were significantly correlated with the diversity of vegetation communities and were an important factor affecting the diversity of vegetation communities.
Forest; vegetation diversity; soil nutrients
S718.55+7
A
1000-2324(2022)02-0271-07
10.3969/j.issn.1000-2324.2022.02.014
2021-05-14
2021-06-14
安徽高校自然科学重点研究项目(KJ2018A0916)
张栋(1983-),男,硕士研究生,讲师,研究方向:园林技术,园林工程施工技术,园林生态工程技术. E-mail:konka388c@163.com
猜你喜欢
现代农村科技(2022年5期)2022-11-18
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
中南林业科技大学学报(2021年11期)2021-12-17
食品安全导刊(2021年20期)2021-08-30
绿色中国(2019年18期)2020-01-04
现代农业研究(2017年11期)2018-01-12
农家科技下旬刊(2017年8期)2017-11-13
热带农业科学(2017年5期)2017-06-12
科技创新导报(2016年21期)2016-12-17
