
LncRNA GATA3-AS1促进乳腺癌细胞的增殖迁移及其机制
2022-04-28 03:15:48习一清
医学新知 2022年2期
江 琪,习一清,魏 蕾
1. 武汉大学基础医学院病理生理学教研室(武汉 430071)
2. 武汉大学中南医院甲乳外科(武汉 430071)
乳腺癌(breast cancer)是女性最常见的恶性肿瘤之一,据2020年全球癌症统计报告显示,乳腺癌新发病例数已超肺癌,位居女性癌症发病人数的首位[1]。2020年,我国女性乳腺癌的发病率和死亡例数分别位列第一及第四位,严重威胁女性健康[2]。近几十年来,尽管包括手术切除、化放疗、靶向治疗等在内的综合治疗手段有效改善了乳腺癌患者的预后,但局部进展及合并远处转移乳腺癌患者的中位生存期仍然较短(约24个月)[3],因此加强对乳腺癌新型生物标志物和治疗靶点的深入探索具有重要的意义。
LncRNA是一类超过200个碱基,且几乎无法编码蛋白质的RNA[4]。LncRNA涉及多种生物事件,并可参与癌症的发生和进展,是敏感和特定的癌症生物标志物之一[5-8]。根据邻近编码基因的染色体区域,可将LncRNA分为正义LncRNA、反义LncRNA、双向LncRNA、内含子LncRNA和基因间LncRNA五类[9]。研究表明,越来越多的反义LncRNA在肿瘤中起调控细胞增殖、迁移和侵袭的作用[10]。例如,LncRNA FEZF1-AS1通过靶向miR-30a/Nanog轴促进乳腺肿瘤的发生[11];LncRNA PCNA-AS1在肝细胞癌中促进细胞增殖[12];LncRNA AFAP1-AS1在非小细胞肺癌中促进细胞迁移[13]等。
LncRNA GATA3-AS1是一种新型反义LncRNA,与T细胞发育和分化有关[14]。GATA3-AS1已被证实可促进肝细胞癌的细胞增殖和迁移[15],但其在乳腺癌中的作用及其机制报道较少,因此本文拟研究GATA3-AS1在乳腺癌中的生物作用并对其分子机制进行初步探索,为乳腺癌新型生物标志物与药物靶点的探索提供新思路。
1 资料与方法
1.1 生物信息学分析
通过 UALCAN在线数据库(http://ualcan.path.uab.edu/index.html)和 bc-GenExMiner在线数据库(http://bcgenex.ico.unicancer.fr/BC-GEM/GEM-requete.php)分析多种肿瘤组织和乳腺癌中GATA3-AS1与GATA3的表达情况,以及不同临床和病理参数乳腺癌患者中GATA3-AS1与GATA3的表达差异。运用lncATLAS在线数据库(https://lncatlas.crg.eu/)预测GATA3-AS1的细胞定位,并通过starBase数据库(https://starbase.sysu.edu.cn/)在线预测GATA3-AS1与GATA3的表达相关性。GATA3-AS1与GATA3的生存分析由Kaplan-Meier Plotter数 据 库(https://kmplot.com/analysis/index.php?p=service&cancer=breast)中提供的乳腺癌数据分析获得。以上结果均认为P<0.05时,差异有统计学意义。
1.2 实验方法
1.2.1 细胞培养
人乳腺上皮细胞MDA-MB-10A和两种乳腺癌细胞系(MDA-MB-231、MCF-7)购于中国科学院上海生命科学研究院细胞资源中心。MDAMB-10A细胞在37℃、5% CO2浓度的培养箱中培养;MCF-7、MDA-MB-231细胞均在含10%胎牛血清的高糖DMEM培养基中培养,其中添加1%的青霉素和链霉素。
1.2.2 质粒转染
特异性shRNA和NC-shRNA购于GenePharma公司,使用Lipofectamine 2000 转染试剂(Invitrogen,美国)对 MDA-MB-231和 MCF-7细胞中的LncRNA GATA3-AS1进行沉默。
1.2.3 CCK8实验
将细胞按照每孔2×103个的密度接种于96孔板,使用含10%血清培养基培养24 h、48 h、72 h、96 h后,每孔中加入10 μL的CCK8试剂(北京庄盟生物),37℃孵育2 h后,使用酶标仪检测各组细胞450 nm处的吸光度值。
1.2.4 划痕实验
将细胞2×105个/mL接种于6孔板中,当细胞达到80%~90%密度时,使用黄色移液器直线刮擦细胞,PBS洗涤划痕处两次除去漂浮细胞,于0 h拍摄划痕宽度,在含5%血清培养基中持续孵育48 h后,再次拍摄并计算划痕宽度及伤口愈合率。
1.2.5 Transwell实验
将细胞用无血清培养基重悬,加入transwell小室(康宁公司)的上腔室,并在transwell下腔室中加入600 μL含1%双抗和10%胎牛血清的培养基。细胞培养24 h后固定细胞,并用0.5%结晶紫染色,擦去上腔室内部细胞后,在显微镜下计数下腔表面的细胞数。
1.2.6 实时定量PCR
采用TRIzol试剂(Invitrogen,美国)从细胞中提取总RNA,通过观察琼脂糖凝胶电泳中28S条带与18S条带的比值检测RNA的纯度和完整性;使用逆转录试剂盒(Thermo,美国)将1 μg RNA逆转录合成cDNA,在CFX96 real-time PCR系统上进行qRT-PCR程序;使用10 μL SYBR Green Realtime PCR Master Mix(Bio-Red,美国)、200 nM正向和反向引物、100 ng cDNA模板,用双蒸馏水使最终反应体系为20 μl。PCR条件设置为:95℃变性30 s,58℃退火30 s,72℃延伸30 s,50个周期。目的基因表达量采用2-ΔΔCt法测定,并以GAPDH为内参,每个实验进行3个重复。其中引物订购于武汉擎科生物公司。

1.2.7 核质分离
使用Ambion PARIS™(Thermo)试剂盒,按照说明书内容从细胞中分离细胞质和核RNA,并进行纯化。使用qRT-PCR检测GATA3-AS1的表达水平。
1.2.8 蛋白印迹实验
首先,使用增强的RIPA裂解缓冲液(北京索拉博)从细胞中提取总蛋白,用BCA蛋白测定试剂盒(上海翊圣生物)对提取的蛋白进行定量;其次,使用聚丙烯酰胺凝胶电泳分离蛋白质,采用湿转移法将分离的蛋白转移到聚偏二氟乙烯膜上;再次,使用5%脱脂牛奶对膜进行封闭处理,且敷育相应的抗体;最后经TBST冲洗后,用显像液显像。
1.3 统计学分析
本研究各项实验均重复三次,研究数据以平均数和标准差表示,使用t检验和方差分析比较各组数据之间的差异,当P<0.05时表示差异有统计学意义。
2 结果
2.1 GATA3-AS1在乳腺癌中的表达量与预后相关
通过UALCAN在线数据库对GATA3-AS1进行表达分析,发现GATA3-AS1在多种肿瘤组织中存在差异表达(图1-A),且相较正常乳腺组织,GATA3-AS1在乳腺癌组织中的表达明显升高(图1-B)。根据数据库提供的两个GATA3-AS1探针(240192_at和 240827_at),Kaplan-Meier生存曲线提示GATA3-AS1表达与乳腺癌总生存期显著负相关,即高表达GATA3-AS1的乳腺癌患者预后较差(图1-C)。
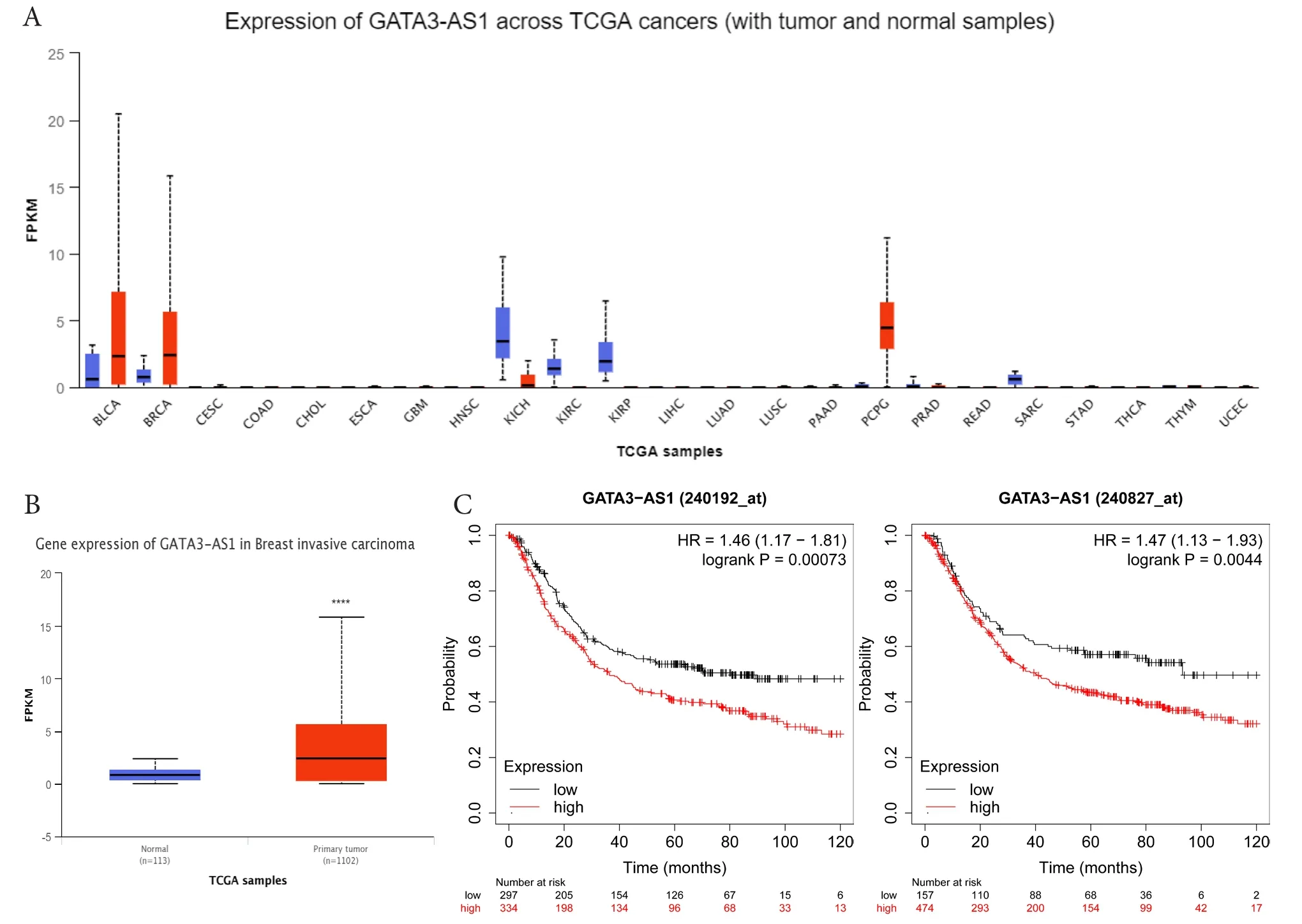
图1 GATA3-AS1的表达水平与预后分析Figure 1. The relationship between the expression level of GATA3-AS1 and the prognosis of breast cancer
利用bc-GenExMiner在线数据库分析不同临床和病理参数乳腺癌患者中GATA3-AS1的表达差异,发现GATA3-AS1的表达与乳腺癌患者年龄之间存在显著相关性,年龄≤51岁的患者GATA3-AS1表达水平明显低于年龄>51岁的患者(图2-A)。同时,GATA3-AS1的表达与Scarff-Bloom-Richardson(SBR)等级(图2-B)以及P53基因突变(图2-C)也存在显著相关性。此外,GATA3-AS1在人类表皮生长因子受体2阴性(HER2-)、雌激素受体阳性(ER+)和孕激素受体阳性(PR+)的乳腺癌患者中表达量明显上调(图2-D、图2-E和图2-F)。相较于基底细胞样型(Basallike)和三阴性乳腺癌(TNBC),GATA3-AS1在非基底细胞样型和非三阴性乳腺癌中的表达水平更高(图2-G和图2-H)。综合数据库中GATA3-AS1在乳腺癌各类分子亚型中表达水平可看出,激素受体A阳性(luminal A)乳腺癌GATA3-AS1表达水平显著高于其他亚型(图2-I)。另外,lncATLAS在线数据库显示GATA3-AS1在人乳腺癌细胞(MCF-7)中主要分布于细胞核内(图2-J),表明该LncRNA可能参与细胞核内的转录调控。
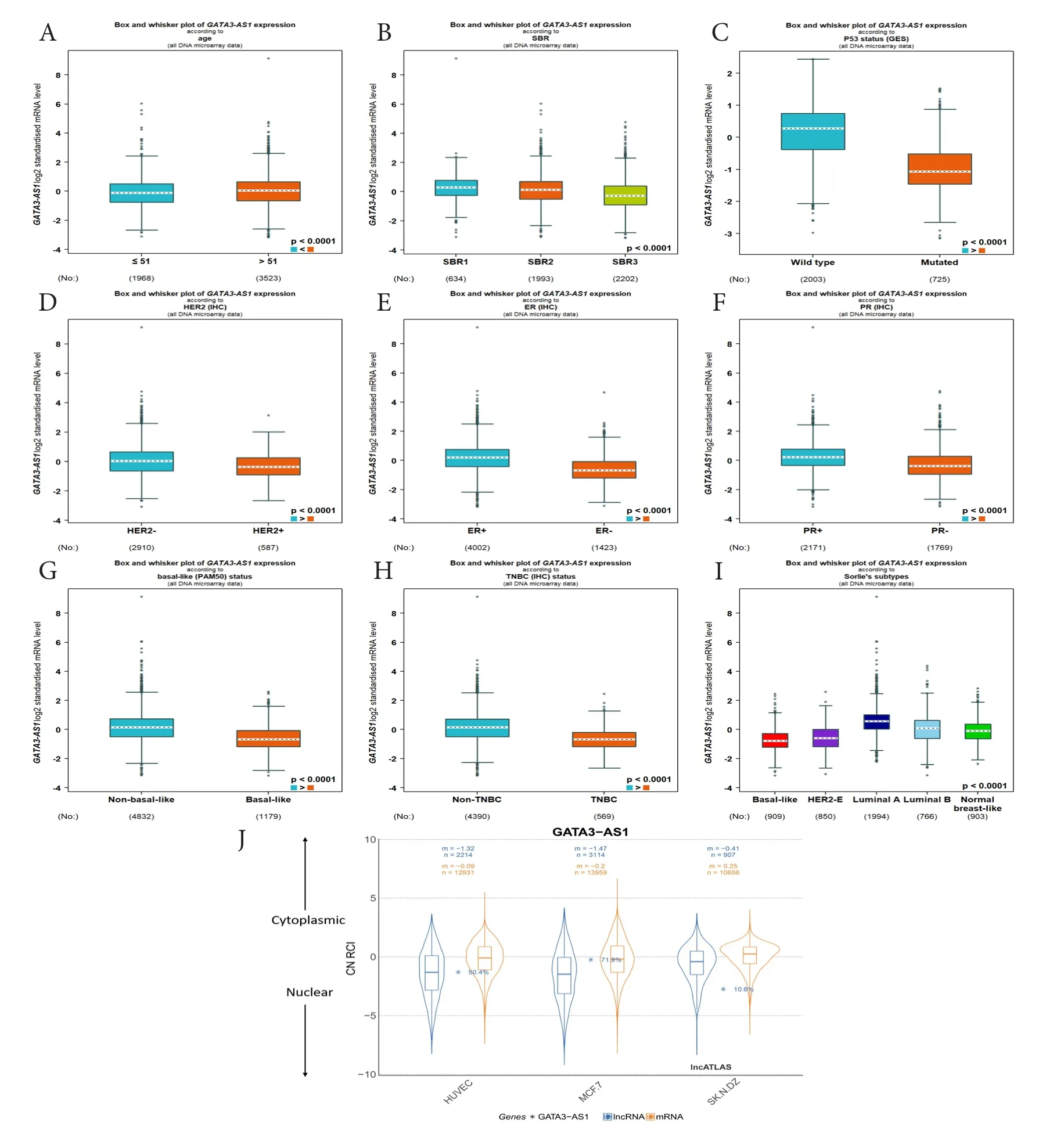
图2 bc-GenExMiner在线数据库关于GATA3-AS1的表达水平与乳腺癌不同临床病理特征关系的分析Figure 2. The relationship between the expression level of GATA3-AS1 and different clinicopathological features of breast cancer was analysed via bc-GenExMiner online database
2.2 GATA3-AS1促进乳腺癌细胞的增殖与迁移
本研究通过qRT-PCR对人类乳腺癌细胞系(MDA-MB-231和MCF-7)以及正常乳腺上皮细胞系(MCF-10A)中GATA3-AS1的表达水平进行验证,结果显示,与MCF-10A细胞相比,GATA3-AS1在MDA-MB-231和MCF-7中显著高表达(P<0.001,图3-A)。核质分离提示GATA3-AS1主要位于MDA-MB-231和MCF-7细胞核内(图3-B),推测GATA3-AS1可能通过影响乳腺癌细胞核内相关基因转录调控乳腺癌的发生发展。此外,通过shRNA获得低表达GATA3-AS1的乳腺癌细胞株,对该细胞株的多种生物功能变化进行观察,qRT-PCR证实MDA-MB-231和MCF-7细胞中的GATA3-AS1可被shRNA有效沉默(P<0.001),且sh1的沉默效率高于sh2,达到80%(图3-C)。因此,本研究使用sh1进行后续体外功能实验。
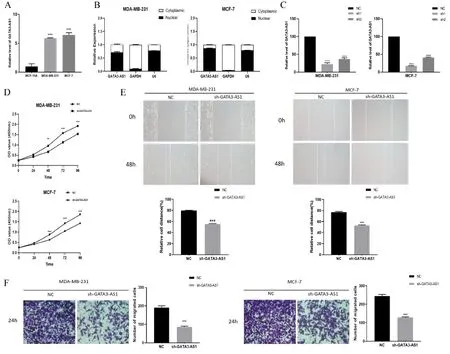
图3 GATA3-AS1促进乳腺癌细胞的增殖与迁移Figure 3. GATA3-AS1 promotes breast cancer cell proliferation and migration
CCK8实验结果显示,与对照组相比,沉默GATA3-AS1后, MDA-MB-231和MCF-7的增殖速度明显下降(P<0.001,图3-D)。后续细胞划痕实验以及Transwell实验证实,沉默GATA3-AS1明显抑制了乳腺癌细胞的二维及三维迁移能力(P<0.001,图3-E、图3-F)。
2.3 乳腺癌组织中GATA3的表达与GATA3-AS1正相关
starBase和 UALCAN数据库均发现 GATA3-AS1的蛋白编码基因GATA3在乳腺癌中的表达与GATA3-AS1的表达呈正相关(图4-A),且GATA3在乳腺癌中表达水平高于正常乳腺组织(P<0.001,图4-B)。Kaplan-Meier生存曲线显示,GATA3的表达与乳腺癌的总生存期显著正相关(图4-C)。利用bc-GenExMiner在线数据库分析不同临床和病理参数的乳腺癌患者GATA3的表达差异,结果显示GATA3在年龄≤51岁的患者中表达水平明显低于年龄>51岁者(图5-A),且与SBR等级(图5-B)以及P53基因突变(图5-C)存在显著相关。此外,GATA3在乳腺癌各分子分型上的表达差异与GATA3-AS1的表达趋势基本一致(图5-D、图5-I)。
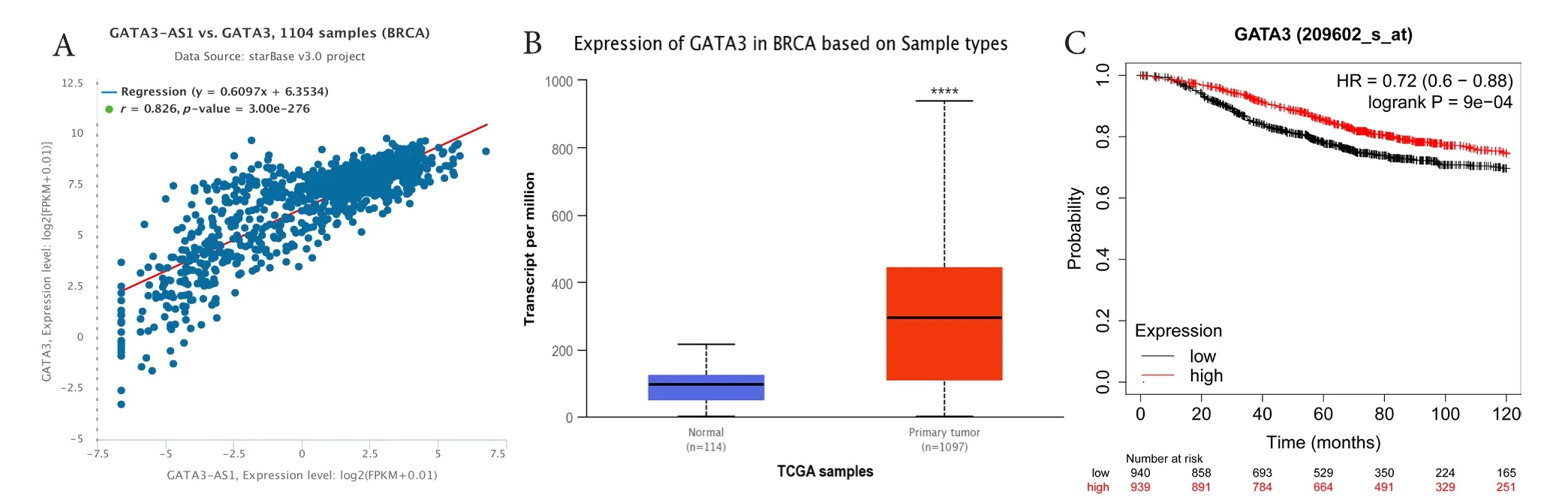
图4 与GATA3-AS1表达相关基因的预测Figure 4. Prediction of genes that associated with GATA3-AS1 expression
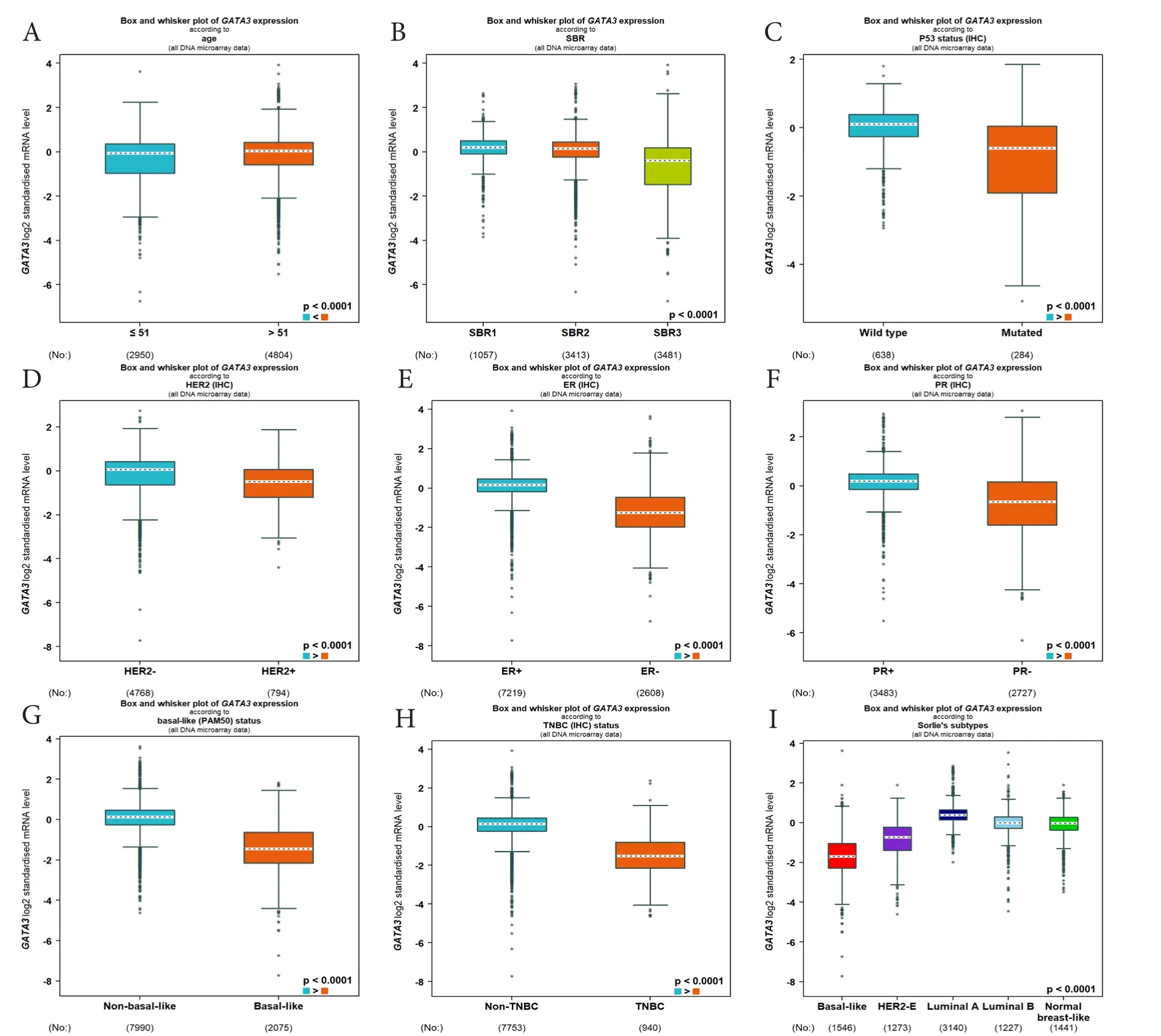
图5 bc-GenExMiner在线数据库对GATA3的表达水平与乳腺癌不同临床病理特征关系的分析Figure 5. The relationship between the expression level of GATA3 and different clinicopathological features of breast cancer was analysed via bc-GenExMiner online database
2.4 沉默GATA3-AS1导致GATA3蛋白水平上升
本研究对GATA3-AS1与其蛋白编码基因GATA3所表达的蛋白进行相关性分析。蛋白印迹实验结果显示,沉默GATA3-AS1后,MDAMB-231和MCF-7细胞中的GATA3蛋白水平随之上升(图6,P<0.01)。
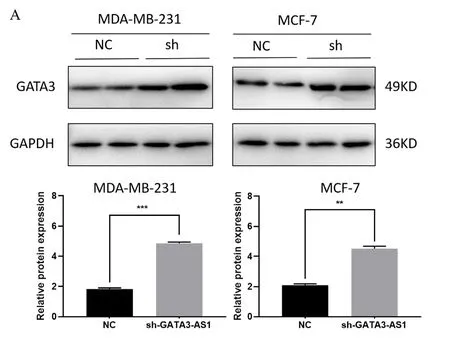
图6 沉默GATA3-AS1之后GATA3蛋白水平上升Figure 6. The protein level of GATA3 increases after the silencing of GATA3-AS1
3 讨论
LncRNA在细胞生长和分化、免疫激活/失活、转录和转录后调控等多种生物学过程中发挥重要作用[16]。近期研究证实多种LncRNA参与乳腺癌的发生发展,如LncRNA HOTAIR通过激活STAT3信号通路,参与乳腺癌组织的上皮-间充质转化[17];LncRNA FGF13-AS1通过抑制乳腺癌细胞糖酵解和干性特性起肿瘤抑制作用[18]。作为基因反义链的转录产物,反义LncRNA也参与了疾病复杂的病理生理过程[19]。GATA3-AS1作为一种新型反义LncRNA,在人类CD4+T细胞亚群中首次被发现[20]。Zhu等应用生物信息学技术对人类膀胱癌组织中的LncRNA表达谱进行分析,发现GATA3-AS1在膀胱癌组织中显著上调[21]。Luo等研究也证实GATA3-AS1可通过沉默抑癌基因促进肝细胞癌细胞增殖和转移[15]。
本研究对乳腺癌细胞中GATA3-AS1表达水平进行检测,结果显示相较正常的乳腺细胞,GATA3-AS1在乳腺癌细胞系中的表达显著上调。进一步通过UALCAN数据库分析发现,GATA3-AS1高表达的乳腺癌患者预后较差,推测在乳腺癌的发生发展中,GATA3-AS1发挥促癌作用。bc-GenExMiner数据库分析显示,GATA3-AS1在非基底细胞样型和非三阴性乳腺癌中的表达水平显著高于基底细胞样型和三阴性乳腺癌,而共识认为三阴性乳腺癌往往预后较差。导致结果矛盾的原因可能为:首先,在UALCAN数据库中有关GATA3-AS1生存分析的对象是未分层的全体乳腺癌患者,未对乳腺癌亚型进行分层预测;其次,不同数据库间纳入的样本存在差异,结果不具有严格意义上的可比性;最后,乳腺癌的发生发展受多基因多因素综合调控,且不同亚型乳腺癌中,LncRNA发挥的作用可能不尽相同。尽管如此,上述生物信息学分析结果提示GATA3-AS1作为潜在生物标志物,在乳腺癌早期预测、疗效评价及预后评估方面具有一定价值。然而,GATA3-AS1与乳腺癌预后关系及其在乳腺癌发生发展中的具体机制,仍需大样本、分层深入的研究。
本研究还利用乳腺癌细胞系MDA-MB-231和MCF-7对GATA3-AS1进行了体外功能实验,下调GATA3-AS1的表达抑制了MDA-MB-231和MCF-7的增殖和迁移能力,证实GATA3-AS1可能在乳腺癌的进展中起促癌作用。研究进一步对GATA3-AS1在乳腺癌中发挥的作用机制进行探讨。通过starBase数据库预测发现,GATA3-AS1的蛋白编码基因GATA3在乳腺癌中的表达与GATA3-AS1呈正相关。有研究表明,反义LncRNA基因转录起始位点与其蛋白编码基因转录起始位点并在一起,可能与mRNA的基因启动子或近端增强子存在竞争关系,进而影响mRNA的转录过程[22-24]。
GATA3在淋巴细胞、乳腺、中枢神经系统、内耳和甲状旁腺等多种人体组织中均有表达,且其表达是这些器官发育的基础,它的失调与急性T细胞淋巴样白血病、乳腺癌、神经母细胞瘤、感音神经性耳聋和甲状旁腺机能减退等多种疾病相关[25]。其中,GATA3的表达与乳腺癌预后的关联已有一定研究基础。Mehra等报道了高表达GATA3患者10年生存率高于低表达患者(84% vs. 55%),表明低表达GATA3提示预后不良[26]。另一项为期10年的随访研究发现,与低表达GATA3肿瘤相比,高表达GATA3的肿瘤发生激素无反应的风险降低了近90%,且预后更好[27]。GATA3作为肿瘤抑制因子[28-29],是GATA3-AS1的蛋白编码基因,且又与GATA3-AS1共享一个启动子区域[30]。此外,核质分离证实GATA3-AS1定位于MDA-MB-231和MCF-7的细胞核内,进一步通过蛋白印迹实验证实,在MDA-MB-231和MCF-7细胞中下调GATA3-AS1表达后,GATA3蛋白水平明显上升,即二者的表达量呈负相关。然而,此结果与starBase数据库预测结果相反。究其原因,一方面可能是在运用starBase数据库预测二者表达量之间关系时,是在mRNA水平分析获得,而实验结果是在蛋白水平上实践得出,结果之间可能存在转录后调控所致的表达差异;另一方面,GATA3被认为是乳腺建立和发育过程中不可或缺的转录因子,在乳腺癌中参与多种分子调控机制[31-35],因此GATA3蛋白的表达受多基因影响,并不一定完全由GATA3-AS1调控。由此猜测GATA3-AS1可能通过抑制其蛋白编码基因GATA3的表达促进乳腺癌的发展,可能成为乳腺癌治疗的新靶点。
生存分析显示,高表达GATA3的乳腺癌患者预后更好,与既往研究一致。此外,通过bc-GenExMiner数据库对不同临床和病理参数的乳腺癌患者GATA3的表达差异进行分析,结果显示具有抑癌作用的GATA3与具有促癌机制的GATA3-AS1在表达量方面与乳腺癌患者的临床病理特征间的相关性趋势十分相似。这是否意味着GATA3-AS1与GATA3之间存在某种平衡,尚有待进一步研究。
基于细胞学实验可知,GATA3-AS1在乳腺癌组织和细胞中表达上调。GATA3-AS1可能通过抑制其蛋白编码基因GATA3的表达,参与调控乳腺癌细胞的增殖和迁移。
猜你喜欢
中老年保健(2022年6期)2022-08-19 01:41:48
小学生学习指导·低年级(2021年6期)2021-09-10 14:07:49
考试与评价·七年级版(2021年4期)2021-08-14 10:45:54
中国生殖健康(2019年2期)2019-08-23 08:11:42
中国生殖健康(2019年6期)2019-01-06 09:20:12
小学阅读指南·低年级版(2018年5期)2018-11-02 10:19:50
祝您健康(2018年5期)2018-05-16 17:10:16
财经(2017年2期)2017-03-10 14:35:35
财经(2016年15期)2016-06-03 07:38:02
财经(2016年3期)2016-03-07 07:44:46
