
铅、镉胁迫对紫穗槐种子萌发及幼苗生理特性的影响
2022-04-25 07:44朱鸿涛
防护林科技 2022年2期
朱鸿涛
摘要:为揭示重金属铅(Pb2+)、镉(Cd2+)对紫穗槐(Amorpha fruticosa)种子萌发及幼苗生长和生理的影响,探究了不同浓度重金属Pb2+(浓度设置为400、800、1200、1600、2 000 mg·L-1)和Cd2+(20、40、60、80、100 mg·L-1)胁迫对紫穗槐种子萌发、幼苗生长、渗透调节物质、抗氧化酶活性及脂质过氧化的影响。结果表明:Pb2+胁迫对紫穗槐种子发芽率、发芽势没有显著影响,Cd2+胁迫对发芽率和发芽势具有抑制作用,胚根胚轴长度与Pb2+、Cd2+浓度负相关;游离脯氨酸和MDA含量与Pb2+、Cd2+浓度正相关,可溶性蛋白随Pb2+、Cd2+浓度增加其含量先上升后下降;在所设置的范围内,CAT、POD、SOD活性均显著上升,且随着浓度的提高,促进作用呈现先升后降的趋势,在Pb2+浓度为1600 mg·L-1、Cd2+浓度为80 mg·L-1时达到最高而后开始降低。Pb2+、Cd2+使紫穗槐幼苗脂质过氧化,并且通过对渗透调节物质和抗氧化酶活性的影响,导致幼苗生长受到抑制,有明显的毒害作用。
关键词:紫穗槐;铅;镉;种子萌发;抗氧化
分类号:Q945.78
Effects of Pb2+、Cd2+ Stress on Seed Germination and Seedling Physiological Characteristics of Amorpha fruticosa
Zhu Hongtao
(School of Forestry, Northeast Forestry University, Harbin 150040, China)
Abstract: The effects of heavy metal Pb2+ and Cd2+ on seed germination, seedling growth and physiology of Amorpha fruticose. The germination experiment of Amorpha fruticose seed were carried out tostudy the effects of explored different concentrations of heavy metal Pb2+ (concentrations set to 400, 800, 1200, 1600, 2000mg/L) and Cd2+ (20, 40, 60, 80, 100mg/L) on germination, seedling growth, antioxidant enzyme activity,membrane lipid peroxidation,and osmoregulation substances of Amorpha fruticose seeds.The results showed that Pb2+ stress had little effect on the germination rate and germination potential of Amorpha fruticose seeds. Cd2+ stress had an inhibitory effect on the germination rate and germination potential. The length of the radicle hypocotyl was negatively correlated with the concentration of Pb2+ and Cd2+.the content of proline and MDAare positively correlated with the concentration of Pb2+ and Cd2+, and the content of the soluble protein increases first and then decreases with the increase of the concentration of Pb2+ and Cd2+.Within the set range, the activities of CAT, POD and SOD were all increased significantly, and with the increase of concentration, the promoting effect showed a trend of increasing at first and then decreasing, reaching the highest and then decreasing at the concentration of Pb2+ 1600mg/L and Cd2+ 80mg/L.Pb2+, Cd2+ make Amorpha fruticose seedlings lipid peroxidation, and through their effects on osmotic adjustment substances and antioxidant enzyme activities, the growth of seedlings is inhibited, which has obvious toxic effects.
Keywords: Amorpha fruticosa; lead; cadmium; seed germination; antioxidation
鉛、镉是常见的重金属污染物[1],已成为土壤富集含量最多的重金属种类[2],进入土壤后不仅影响土壤的理化性质,还会影响植物的生长发育,通过植物吸收富集于植物体内,最终经由食物链进入人类体内富集,严重危害人体健康[3]。利用生长迅速、生物量大、根系发达的木本植物进行重金属污染修复,具有安全环保、无二次污染且成本低廉的优点,是修复重金属污染土壤的研究热点[4]。在植物生长的过程中,种子萌发时期最为脆弱,受外界影响最为明显,是评价植物抗逆性的重要时期[5]。
紫穗槐(Amorpha fruticosa)是一种原产自美国的落叶灌木,为豆科紫穗槐属,耐瘠、耐旱,具有固氮能力,又因其生长迅速,根系发达,生物量大,是优良的护坡植物,在我国多省份均有栽种[6-8]。已有研究发现,紫穗槐具有较强的對干旱、盐碱和重金属的耐受性[9-13]以及重金属富集能力[14],能够在干旱的沙地和受重金属污染的土壤上生长[15],修复受重金属污染的土壤[16],但对于重金属铅、镉抗逆性的研究较少。本研究以紫穗槐种子为试验材料,研究不同浓度重金属铅、镉对种子萌发、幼苗生长及生理水平的影响,为重金属污染地区的土壤修复提供理论基础和科学依据。
1材料与方法
1.1 试验材料
供试种子于2020年9月15日采集于哈阿高速公路两侧长势良好的紫穗槐植株,种子千粒质量12.21g,置于实验室常温避光保存。
试剂配置成Pb(NO3)2和CdCl2溶液,Pb2+溶液浓度设定为400、800、1200、1600、2000 mg·L-1,Cd2+溶液浓度设定为20、40、60、80、100 mg·L-1,以蒸馏水处理作为对照。
1.2 试验方法
选取大小一致、健康饱满的紫穗槐种子,剥去外壳后使用800目砂纸打磨种皮,使用浓度为75%的乙醇消毒3min,用蒸馏水冲洗数次,将种子在对应浓度溶液中浸泡4h[17],使用无菌滤纸吸干水分备用。将两张滤纸平铺在培养皿(直径9cm)底部,于高压灭菌锅中120℃高温高压灭菌20min。灭菌后的培养皿中加入对应浓度溶液5mL,充分润湿滤纸且无气泡。每对培养皿中放入30粒紫穗槐种子,每个处理3次重复。将培养皿置于人工气候箱中培养,设定温度25±1℃,湿度60%,光照强度8 000 lx,光照时间14h·d-1。根据滤纸湿润情况,每1-2 d补充对应浓度的溶液。每天观察记录种子发芽数,以胚根突破种皮长度达到种子长度的一半为萌发标志,种子萌发恒定后(连续3d没有新的种子萌发),结束试验,进行指标的测定和计算。
1.3指标测定
1.3.1紫穗槐幼苗胚根长度、胚轴长度的测定和计算
采用最小刻度为1mm的刻度尺测量萌发后幼苗的胚根长度、胚轴长度,每个处理3组重复。
1.3.2 紫穗槐种子发芽率、发芽势、发芽指数、活力指数的计算
发芽率(%)=试验期间种子发芽数/供试种子总数×100%;发芽势=发芽高峰发芽种子数/供试种子数×100%;发芽指数GI=∑Gt/Dt,式中:Gt—种子在第t日的发芽数,Dt—相应的发芽天数;活力指数=幼苗长度×发芽指数
1.3.3 紫穗槐幼苗生理指标测定
过氧化物歧化酶(POD)活性采用愈创木酚法测定[18];过氧化氢酶(CAT)活性采用紫外吸收法测定[18];超氧化物歧化酶(SOD)活性采用氮蓝四唑光还原法测定[18];可溶性蛋白采用考马斯亮蓝G-250染色法测定[18];游离脯氨酸含量采用酸性茚三酮法测定[18];丙二醛(MDA)采用硫代巴比妥酸(TBA)法测定[18];
1.4数据处理与统计分析
试验数据采用Microsoft Excel软件进行统计,采用SPSS26.0进行单因素方差分析(ANOVA)和Duncan检验比较差异显著性,采用Origin2021进行图形绘制。
2结果与分析
2.1不同浓度Pb2+、Cd2+溶液对紫穗槐种子萌发的影响
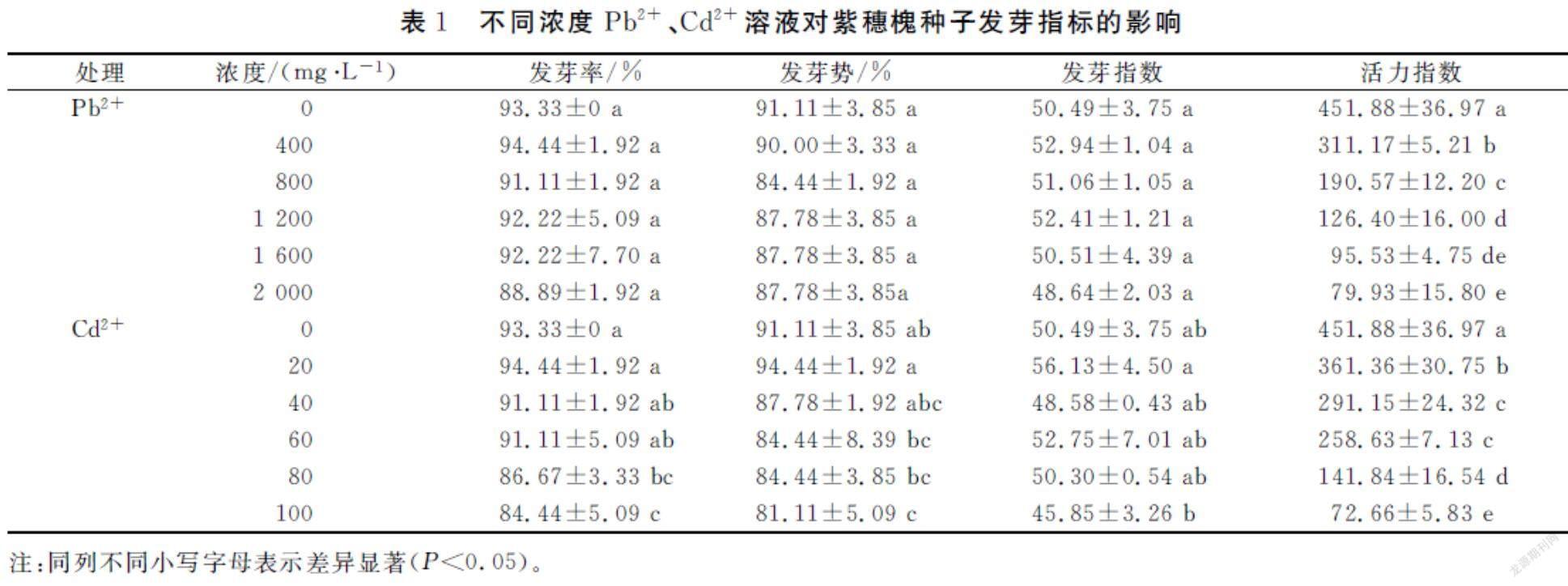
由表1可知,随着Cd2+浓度升高,紫穗槐种子发芽率、发芽势和活力指数显著降低,当Cd2+浓度为100 mg·L-1时,抑制作用最强,发芽率和发芽势分别为84.44%和81.11%,比对照降低了8.89和10.0个百分点,活力指数比对照显著降低83.92%;Pb2+浓度变化对发芽率、发芽势和发芽指数影响差异不显著,对活力指数影响显著,比对照降低了83.92%。
2.2不同浓度Pb2+、Cd2+溶液对紫穗槐幼苗生长的影响
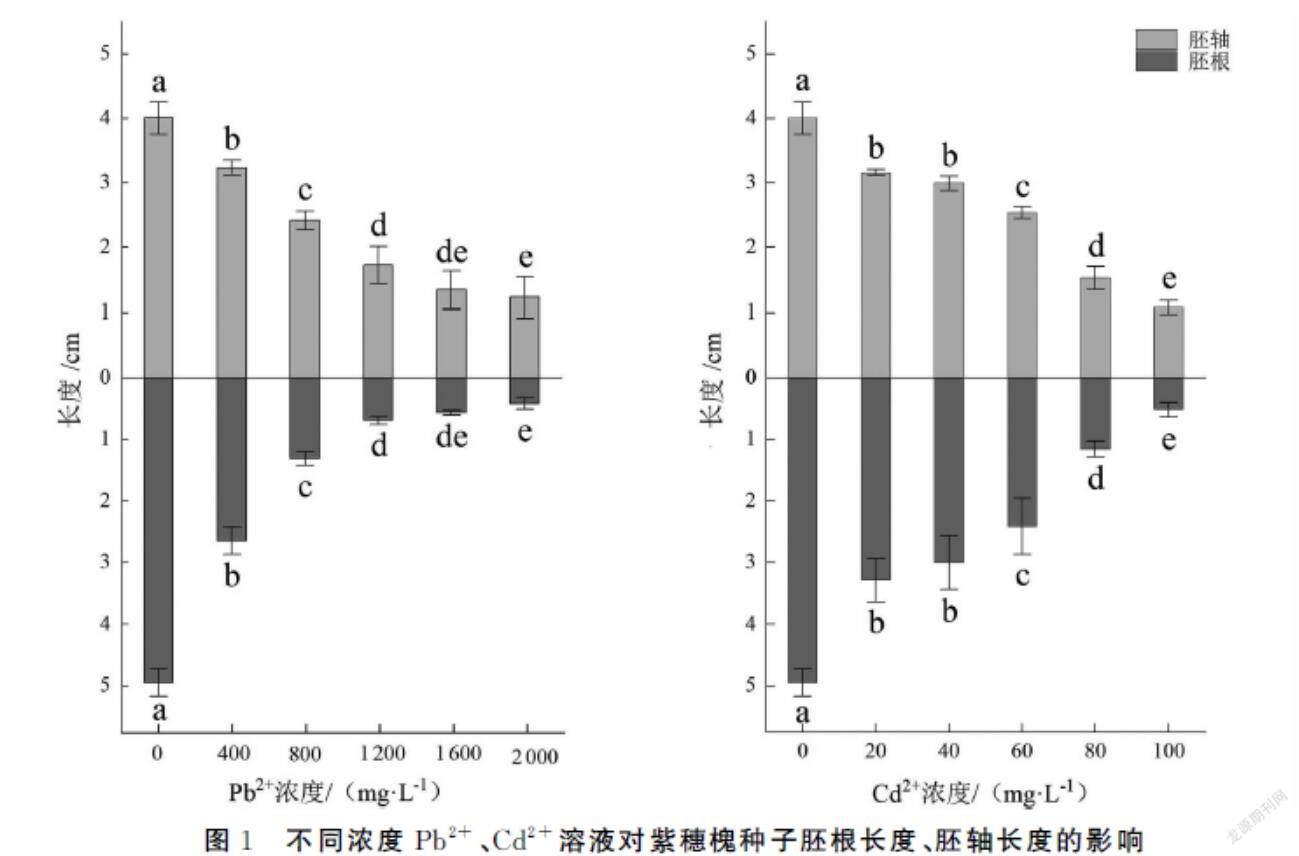
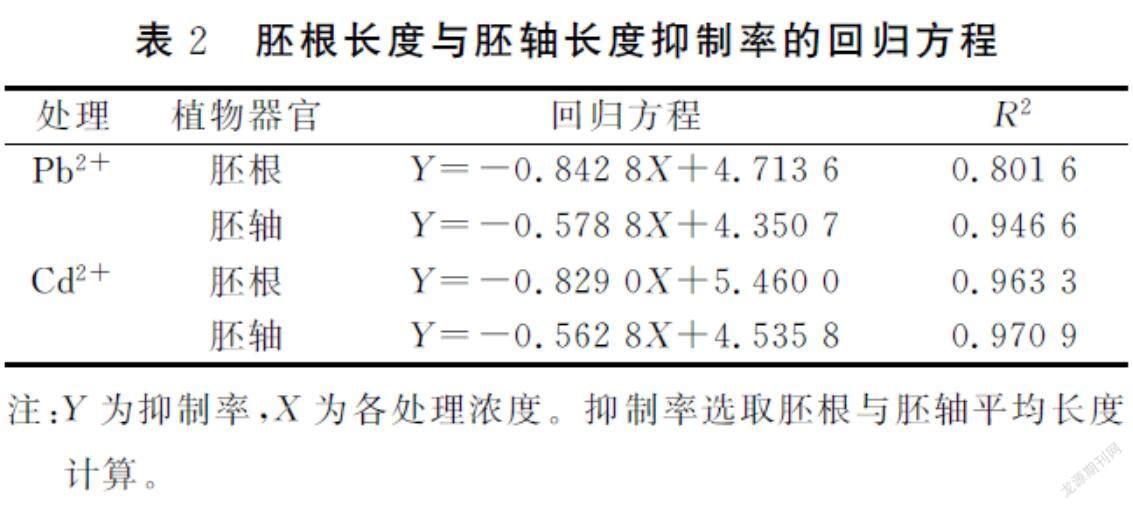
由图1可知,Pb2+、Cd2+浓度与紫穗槐幼苗胚根长度和胚轴长度显著负相关,当Pb2+浓度为2000 mg·L-1和Cd2+浓度为100 mg·L-1时,抑制作用最强,胚根长度分别为0.42和0.52cm,胚根抑制率为91.52%、88.48%;胚轴长度为1.23和1.08cm,胚轴抑制率为69.33%和73.07%。
2.3不同浓度Pb2+、Cd2+溶液对紫穗槐渗透调节物质含量的影响
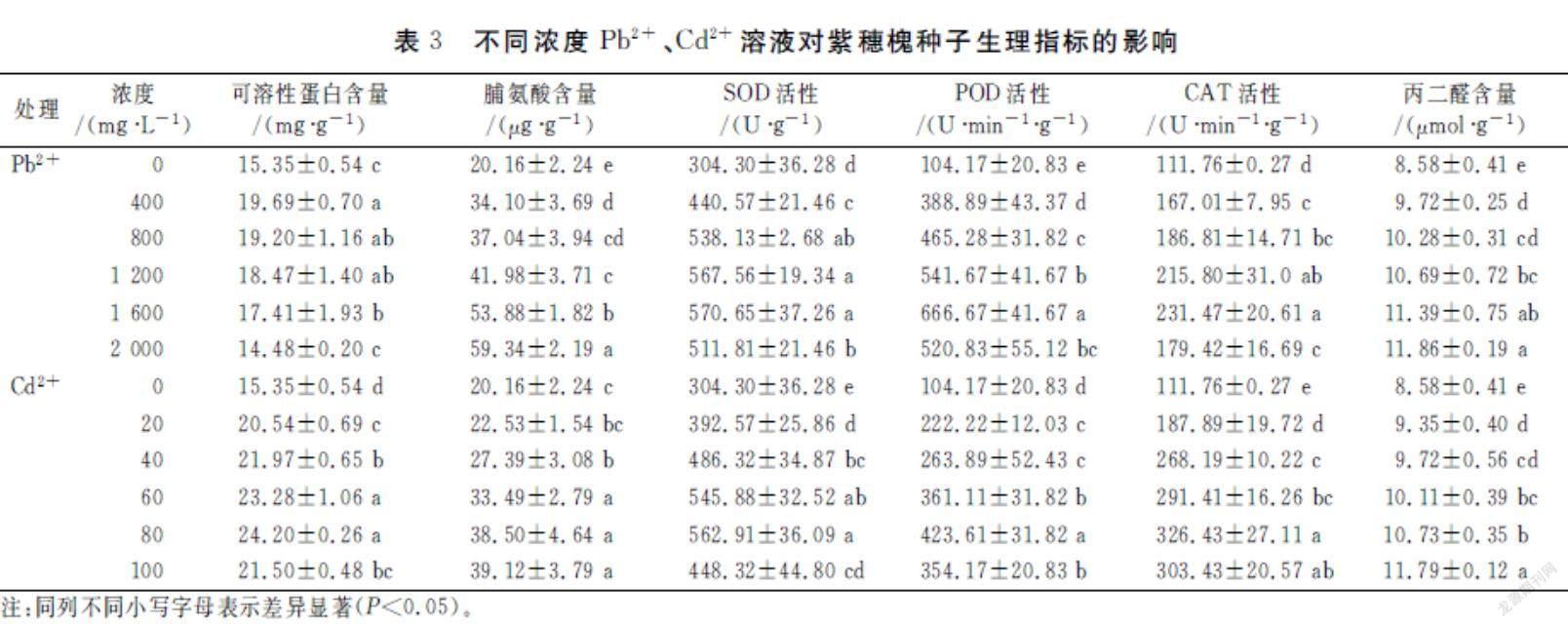
由表3可知,紫穗槐幼苗可溶性蛋白含量随着Pb2+、Cd2+浓度的升高而先升高后降低,各处理组均高于对照组,且差异显著,当Pb2+浓度为400 mg·L-1和Cd2+浓度为80 mg·L-1时,可溶性蛋白含量最高,比CK增大了28.27%和57.65%。脯氨酸含量与Pb2+、Cd2+浓度显著正相关,当Pb2+浓度为2000 mg·L-1和Cd2+浓度为100 mg·L-1时,脯氨酸含量最高,比CK增大了194.35%和94.05%。
2.4不同浓度Pb2+、Cd2+溶液对紫穗槐抗氧化酶的影响
由表3可知,SOD、POD、CAT活性随Pb2+、Cd2+浓度增加而先上升后下降,各处理组均高于对照组,且差异显著。当Pb2+浓度为1600 mg·L-1和Cd2+浓度为80 mg·L-1时,SOD活性最高,比CK上升了87.53%和84.99%;当Pb2+浓度为1600 mg·L-1和Cd2+浓度为80 mg·L-1时,POD活性最高,比CK上升了539.98%和306.65%;当Pb2+浓度为1600 mg·L-1和Cd2+浓度为80 mg·L-1时,CAT活性最高,比CK上升了107.11%和192.08%。
2.5不同浓度Pb2+、Cd2+溶液对紫穗槐幼苗脂质过氧化的影响
丙二醛(MDA)是脂质过氧化的代谢产物,其含量的多少可以表明脂质过氧化程度的高低[19]。由表3可知,紫穗槐幼苗MDA含量与Pb2+、Cd2+浓度显著正相关,当Pb2+浓度为2000mg/L和Cd2+浓度为100mg/L时,MDA含量最高,比CK增高了38.23%和37.41%,表明此时其脂质过氧化程度最严重。
3结论与讨论
3.1Pb2+、Cd2+对种子萌发及幼苗生长的影响
重金属Pb2+、Cd2+是植物生长发育过程中不需要的元素,植物从种子萌发到幼苗生长阶段都会受到其深远的毒害作用,可能会出现植物生长变慢、生物量减小、植株矮小、根系生长受到抑制等现象,严重时植株死亡[20]。本试验表明,重金属Pb2+、Cd2+对紫穗槐种子发芽率、发芽势的影响不同,Cd2+显著抑制种子发芽率和发芽势,而Pb2+对发芽率和发芽势影响不显著。这可能是种子通过吸涨作用完成内部储藏物质的代谢分解,因此铅对种子的毒害作用会被部分掩盖[21-23];而镉的毒性远强于铅,即使少量也会使种子内部的蛋白质酶和淀粉酶活性受到抑制,导致萌发所需的物质与能量匮乏,使发芽率降低[24, 25]。
本实验中,胚根长度、胚轴长度等在Pb2+、Cd2+低浓度时即有抑制效果,浓度越高抑制效果越强;Pb2+、Cd2+对紫穗槐幼苗胚根长度的抑制作用比胚轴更强,说明Pb2+、Cd2+对幼苗胚根的毒害作用比胚轴更显著,这可能是因为Pb2+、Cd2+被根系细胞壁的脂质和蛋白质大量固定,影响了Ca2+跨膜运输,导致Ca-ATP酶与钙调素CAM无法激活,抑制了根部细胞的有丝分裂,抑制植物根系对外界物质的吸收,最终抑制胚轴的生长发育[26]。
3.2 Pb2+、Cd2+对生理指标的影响
逆境因子会引发渗透胁迫,植物为了适应外界条件抵抗胁迫压力从而进行渗透调节,渗透调节是植物抵抗逆境胁迫的有效方式[27]。在重金属胁迫下,为了维持植物细胞和组织的渗透平衡,大量的渗透调节物质,如可溶性蛋白和游离脯氨酸,在植物体内合成和积累[28],其含量是评价植物抗重金属胁迫能力的生理指标。本试验中,当Pb<400 mg·L-1,Cd<80 mg·L-1时紫穗槐幼苗的可溶性蛋白含量随浓度升高而高,当Pb>400 mg·L-1,Cd>80 mg·L-1时可溶性蛋白浓度开始降低;游离脯氨酸含量随着Pb2+、Cd2+浓度升高而显著增加;这表明Pb2+、Cd2+脅迫能够诱导紫穗槐产生更多的可溶性蛋白来维持细胞渗透压,从而提高对Pb2+、Cd2+胁迫的抗逆性。
Pb2+、Cd2+胁迫会刺激植物产生大量的活性氧自由基,而植物体内的抗氧酶防卫系统能清除自由基,保护植物细胞免受氧化损伤[29]。植物的抗氧化酶系统主要由过氧化氢酶(CAT)、过氧化物酶(POD)和超氧化物歧化酶(SOD)组成[30]。SOD将超氧阴离子自由基(-O2-)通过歧化反应歧化为养活性较弱的H2O2和羟基自由基(-OH),再由POD、CAT将H2O2分解成H2O,从而减少自由基的损伤,在低浓度重金属胁迫下,会促使植物抗逆基因表达,激活植物抗氧化系统,随着植物体内-O2-和H2O2过量积累,植物体内活性氧自由基的动态平衡被打破,酶系统开始被破坏,酶活性降低[31-33]。当Pb2+<1600 mg·L-1、Cd2+<80 mg·L-1时紫穗槐幼苗的抗逆基因表达CAT、POD、SOD三种酶活性均随浓度升高而增大,当Pb2+>1 600 mg·L-1、Cd2+>80mg/L时抗氧化酶系统被破坏,CAT、SOD、POD酶活性开始显著下降。
丙二醛(MDA)是植物中活性氧自由基和脂质相互作用进而导致脂质过氧化的产物,逆境胁迫会引起丙二醛的累积,造成蛋白质、核酸等大分子的交联聚合,降低植物细胞中蛋白质代谢效果,也会对细胞膜系统、光合作用等产生较大的危害,丙二醛含量间接反映了细胞水平由于自由基导致的氧化损伤程度[19, 34]。紫穗槐幼苗的丙二醛含量随着Pb2+、Cd2+浓度的增加而显著上升,说明Pb2+、Cd2+胁迫使紫穗槐细胞膜受到破坏。
综上所述,Pb2+、Cd2+紫穗槐幼苗的发育虽然受到了抑制,但通过抗氧化酶和渗透调节系统的作用,使紫穗槐种子仍能够保持较高的萌发率,说明紫穗槐对Pb2+、Cd2+单一胁迫具有较好的抗逆性。在自然环境中,重金属胁迫往往是以复合胁迫的方式存在,因此,研究重金属之间的复合胁迫及野外试验,对进一步了解紫穗槐对重金属的抗逆性具有更重要的意义。
参考文献
[1] World Health Organization. Inorganic lead [J]. 1995.
[2] 中华人民共和国环境保护部, 中华人民共和国国土资源部. 全国土壤污染状况调查公报 [J]. 环境教育, 2014(6): 7-10.
[3] 王泓博, 苟文贤, 吴玉清, 等. 重金属污染土壤修复研究进展:原理与技术 [J]. 生态学杂志, 2021, 40(8): 2277-2288.
[4] MAHMOOD Q, MIRZA N, SHAHEEN S. Phytoremediation Using Algae and Macrophytes: I [J]. 2015.
[5] 尚宏芹, 高昌勇. 镉、汞胁迫对桔梗种子萌发、幼苗生理生化特性及镉、汞含量的影响 [J]. 核农学报, 2018, 32(6): 1211-1219.
[6] KEREN S, DIACI J, MOTTA R, et al. Stand structural complexity of mixed old-growth and adjacent selection forests in the Dinaric Mountains of Bosnia and Herzegovina [J]. Forest Ecology and Management, 2017, 400.
[7] LJILJANA K, ZORANA K, TANJA Ž P, et al. Phytotoxic effect of invasive species Amorpha fruticosa L. on germination and the early growth of forage and agricultural crop plants [J]. Ecological Research, 2020, 36(1).
[8] 管培军. 山东省砂石山地6个造林树种根系形态的比较 [D].泰安:山东农业大学, 2014.
[9] 刘刚, 张伟龙. 野樱桃和紫穗槐对矿区土壤重金属耐受特征分析 [J]. 世界热带农业信息, 2021, (10): 1-5.
[10] XINZHU S, SONGMIAO H, XIN W, et al. De novo assembly of Amorpha fruticosa L. transcriptome in response to drought stress provides insight into the tolerance mechanisms [J]. PeerJ, 2021, 9.
[11] 王竞红, 张秀梅, 陈艾, 等. 紫穗槐幼苗根系生理特性和解剖结构对PEG-6000模拟干旱的响应 [J]. 生态学报, 2018, 38(2): 511-517.
[12] 王涛, 蒙仲举, 张佳鹏, 等. NaCl胁迫对紫穗槐幼苗生长及生理特性的影响 [J]. 西北林学院学报, 2021, 36(1): 25-30.
[13] 邹丽娜, 周志宇, 颜淑云, 等. 盐分胁迫对紫穗槐幼苗生理生化特性的影响 [J]. 草业学报, 2011, 20(3): 84-90.
[14] 施翔, 陈益泰, 王树凤, 等. 废弃尾矿库15种植物对重金属Pb、Zn的积累和养分吸收 [J]. 环境科学, 2012, 33(6): 2021-2027.
[15] 赵鑫, 辛一凡, 张应龙,等. 不同植被类型对毛乌素沙地背风坡土壤水分时空变化的影响 [J]. 西北林学院学报, 2021, 36(3): 36-43.
[16] 宋清玉. 螯合剂对紫穗槐和栾树幼苗修复铅污染土壤的影响 [D].杨凌:西北农林科技大学, 2017.
[17] WEI B G, YANG L S. A review of heavy metal contaminations in urban soils, urban road dusts and agricultural soils from China [J]. Microchemical Journal, 2010, 94(2): 99-107.
[18] 王学奎. 植物生理生化实验原理和技术.第2版 [M]. 2版. 2006.
[19] MAHMOUDI H, SALAH I B, ZAOUALI W, et al. Priming-induced changes in germination, morpho-physiological and leaf biochemical responses of fenugreek ( Trigonella foenum-graecum ) under salt stress [J]. Plant Biosystems - An International Journal Dealing with all Aspects of Plant Biology, 2020, 154(5).
[20] 孫金金, 鱼小军, 王金辉, 等. 重金属Cu2+、Cd2+和Pb2+对8种禾草种子萌发和幼苗生长的影响 [J]. 草地学报, 2018, 26(3): 673-683.
[21] 段代祥, 刘俊华. 重金属铅胁迫对绿豆种子萌发及幼苗生长的抑制效应 [J]. 种子, 2021, 40(01): 84-87+98.
[22] 杨楠. 干旱及铅胁迫对主要造林树种种子萌发与幼苗生长的影响 [D].杨凌:西北农林科技大学, 2012.
[23] 齐文靖, 于晗, 张佳慧, 等. 不同重金属胁迫对绿豆种子萌发和幼苗部分生理指标的影响 [J]. 北方园艺, 2018(21): 1-5.
[24] 陈丽. 镉胁迫下绿豆种子萌发过程中代谢组学的响应 [D].南昌:南昌大学, 2017.
[25] 张义贤. 汞、镉、铅胁迫对油菜的毒害效应 [J]. 山西大学学报:自然科学版, 2004 (4): 410-413.
[26] KOPITTKE P M, ASHER C J, MENZIES N W. Toxic effects of Ni2+ on growth of cowpea (Vigna unguiculata) [J]. Plant and Soil, 2007, 292(1-2): 283-289.
[27] 周紫鹃, 苏培玺, 解婷婷, 等. 不同生境下红砂(Reaumuria soongorica)的生理生化特征及适应性 [J]. 中国沙漠, 2014, 34(4): 1007-1014.
[28] 徐维杰, 王庆亚, 杨文嘉, 等. 铬(Cr6+)胁迫对不同基因型小麦种子萌发及幼苗抗氧化和渗透调节的影响 [J]. 麦类作物学报, 2017, 37(8): 1112-1119.
[29] 黄玉山, 罗广华, 关棨文. 镉诱导植物的自由基过氧化损伤 [J]. Acta Botanica Sinica, 1997 (6): 522-526.
[30] 姜艺, 黄琳丽. 镉胁迫对植物的影响探究 [J]. 南方农业, 2020,14(30): 138-139.
[31] SRIVASTAVA R K, PANDEY P, RAJPOOT R, et al. Cadmium and lead interactive effects on oxidative stress and antioxidative responses in rice seedlings [J]. Protoplasma, 2014, 251(5).
[32] TRIANTAPHYLIDèS C, HAVAUX M. Singlet oxygen in plants: production, detoxification and signaling [J]. Trends in Plant Science, 2009, 14(4).
[33] ZHANG Y, DENG B, LI Z. Inhibition of NADPH oxidase increases defense enzyme activities and improves maize seed germination under Pb stress [J]. Ecotoxicology and Environmental Safety, 2018, 158.
[34] 杨国远, 万凌琳, 雷学青, 等. 重金属铅、铬胁迫对斜生栅藻的生长、光合性能及抗氧化系统的影响 [J]. 环境科学学报, 2014, 34(6): 1606-1614.
猜你喜欢
疯狂英语·新阅版(2021年8期)2021-09-10
时代英语·高二(2021年4期)2021-07-29
医学概论(2021年19期)2021-01-21
疯狂英语·读写版(2020年12期)2020-12-28
保健与生活(2019年13期)2019-08-27
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
时尚北京(2016年12期)2017-01-16
天津农业科学(2016年12期)2017-01-11
现代农业科技(2016年20期)2016-12-20
