
云南哈尼族地方水稻‘月亮谷’对褐飞虱取食危害的生理反应
2022-04-15 06:08肖关丽
中国农学通报 2022年9期
黄 钰,陈 斌,肖关丽
(1云南农业大学农学与生物技术学院,昆明 650201;2云南农业大学植物保护学院,昆明 650201)
0 引言
水稻(Oryza sativaL.)是全球极其重要的粮食作物[1]。然而,害虫是严重影响水稻产量和质量的重要因素,其中对水稻种植影响较大的属稻飞虱中的褐飞虱。褐飞虱(Nilaparvata lugensStål)是许多亚洲国家和中国水稻种植中最为首要的迁飞性害虫,其发生范围不断扩大,危害严重[2],虫量大时可对水稻稻株造成严重伤害,使水稻大面积减产甚至颗粒无收[3-4]。此外,褐飞虱还会传播水稻锯齿叶矮缩病(Ragged stunt virus,RSV)和草状矮缩病(Grassy stunt virus,GSV)等病毒病[5]。其中水稻抗性品种的严重缺乏是中国稻飞虱危害加重的重要原因之一[6]。‘月亮谷’(‘Acuce’)是云南省元阳哈尼梯田长期种植保留下来的优良地方水稻品种,种植历史悠久,有良好的病虫害抗性[7]。研究‘月亮谷’对褐飞虱取食的主要防御反应,探明其抗虫机理,深入挖掘该品种优良特性,为更好利用该地方品种提供理论依据。植物在与昆虫长期的协同进化过程中,进化形成了减轻昆虫取食对其伤害多种防御机制,如植株营养状况的变化、产生次生代谢有毒物质或防御蛋白等[8-9]。稻飞虱取食危害后水稻稻株内的可溶性糖含量下降[10-12],丙二醛(MDA)和过氧化氢(H2O2)含量上升[13-14]、多酚氧化酶(PPO)活性表现出先上升后下降的趋势变化[15],过氧化物酶(POD)活性在为害后显著升高[11]。然而,不同水稻品种对不同种类稻飞虱取食危害后的生理响应不尽一致[10,16-17]。稻飞虱对化学农药的抗药性的出现是制约稻飞虱有效防控的重要因素[18],因此,利用水稻抗性品种控制稻飞虱已成为稻飞虱持续控制的重要措施,而开展水稻对稻飞虱取食危害的生理反应格外重要。云南地处云贵高原,拥有十分丰富的地方稻种资源及抗性资源[19][20]。其水稻地方品种中蕴藏着新的抗性基因,具有丰富的遗传多样性,在改良水稻品种抗虫性方面具有极高的利用价值[21]。‘月亮谷’是云南元阳哈尼梯田种植面积最大的主栽地方品种,有上百年的种植历史,具有较强的抗病虫害特性和丰富的遗传多样性,是研究水稻遗传育种的理想材料[22-24]。研究褐飞虱取食胁迫对‘月亮谷’中可溶性糖、丙二醛(MDA)、过氧化氢(H2O2)含量,以及过氧化物酶(POD)、多酚氧化酶(PPO)主要防御酶活性的影响,探明地方品种‘月亮谷’对褐飞虱取食胁迫的生理响应。利用抗性品种控制稻飞虱的为害是最为经济有效的防治措施,探究水稻抗褐飞虱的机制是筛选和培育抗虫品种的基础工作。
1 材料与方法
1.1 试验材料
水稻品种为云南元阳哈尼族传统水稻品种‘月亮谷’,由云南农业大学生物资源利用与保护国家重点实验室提供。
抗、感褐飞虱水稻品种‘Rathu Heenati’(‘RHT’)和‘Taichung Native 1’(‘TN1’)均由中国水稻研究所提供。
褐飞虱在感虫水稻品种‘TN1’上,饲养于云南农业大学植物保护学院昆虫实验室的培养箱(28 ~30℃)内。
1.2 试验方法
1.2.1 试验处理 播种前浸种催芽,水稻种子露白后在育苗盘中分别种植‘月亮谷’及抗感对照‘TN1’与‘RHT’3个水稻品种,待水稻幼苗长至2叶1心期,挑选各品种健康壮苗5株,移栽至准备好的透明塑料杯(口径90 mm、高度175 mm、容量700 mL),雌雄比例1:1,每杯接入5对褐飞虱成虫,然后用纱布对塑料杯封口,并置于恒温培养箱中于16L:8D光周期,28 ~30℃,相对湿度70%±5%条件下饲养。试验设3次重复,以不接褐飞虱的水稻苗作为空白对照。分别取对照和接虫后3、6、12、24、48、72和96 h时水稻苗全部茎叶,分别装入自封袋(100 mm×70 mm)并置于液氮速冻1 min,取出放入-80℃冰箱保存待测。
1.2.2 稻株可溶性糖含量测定 称取样品0.1 g,采用可溶性糖试剂盒,由苏州格锐思生物科技有限公司生产(下同),分光光度计测定620 nm处的吸光值。利用测定数值计算可溶性糖含量。
1.2.3 稻株次生代谢产物含量测定
(1)稻株内MDA含量测定 称取样品0.1 g,采用MDA试剂盒,分光光度计分别测定532 nm和600 nm处吸光值。利用测定数值计算MDA含量
(2)稻株H2O2含量测定称取样品0.1 g,采用H2O2试剂盒,分光光度计测定415 nm处的吸光值。利用测定数值计算H2O2含量。
1.2.4 稻株保护酶活性测定
(1)稻株PPO活性测定 称取样品0.1 g,采用PPO试剂盒,利用分光光度计测定420 nm时的吸光值。利用测定数值计算PPO含量。
(2)稻株POD活性测定 称取样品0.1 g,采用POD试剂盒,分光光度计测定470 nm处的吸光值。利用测定数值计算POD含量。
1.3 统计分析
采用软件Excel 2010整理试验数据,利用单因素多重比较Duncan's新复极差法分析比较各测定数据间的差异显著性,所有数据分析均在DPS 2005软件中完成。
2 结果与分析
2.1 褐飞虱为害后水稻可溶性糖含量变化
结果表明(图1),褐飞虱取食3 h后,‘月亮谷’植株内可溶性糖的含量显著下降(P<0.05),降至谷值2.00 mg/gFW。而在取食胁迫6 h后,可溶性糖的含量又呈现上升,到48 h时达峰值8.98 mg/gFW,之后又缓慢下降;对于‘TN1’,褐飞虱取食3 h后,其体内可溶性糖含量达谷值5.06 mg/gFW,取食12 h后,其可溶性糖含量达峰值13.67 mg/gFW,取食72 h时达第二个峰值12.90 mg/gFW,取食96 h后与对照相近;褐飞虱取食3 h后,‘RHT’的可溶性糖含量显著下降,且在取食3 ~24 h后无显著变化,取食48 ~72 h后又显著增加,取食96 h后,‘RHT’可溶性糖含量又显著下降。褐飞虱取食胁迫后,‘月亮谷’可溶性糖含量的变化趋势与RHT可溶性糖含量的变化趋势相近。

图1 褐飞虱取食胁迫后水稻植株可溶性糖含量
从褐飞虱取食危害后供试水稻植株内可溶性糖含量增加率来看(表1),褐飞虱取食3 ~96 h后,‘月亮谷’、‘TN1’和‘RHT’可溶性糖含量相较于对照平均增加了-42.85%、-21.78%和-39.46%,‘月亮谷’植株体内可溶性糖含量增加率显著低于‘TN1’,而与‘RHT’间无显著差异。由此表明,褐飞虱取食危害后,‘月亮谷’和抗虫品种‘RHT’较感虫品种‘TN1’可溶性糖含量下降更为明显。
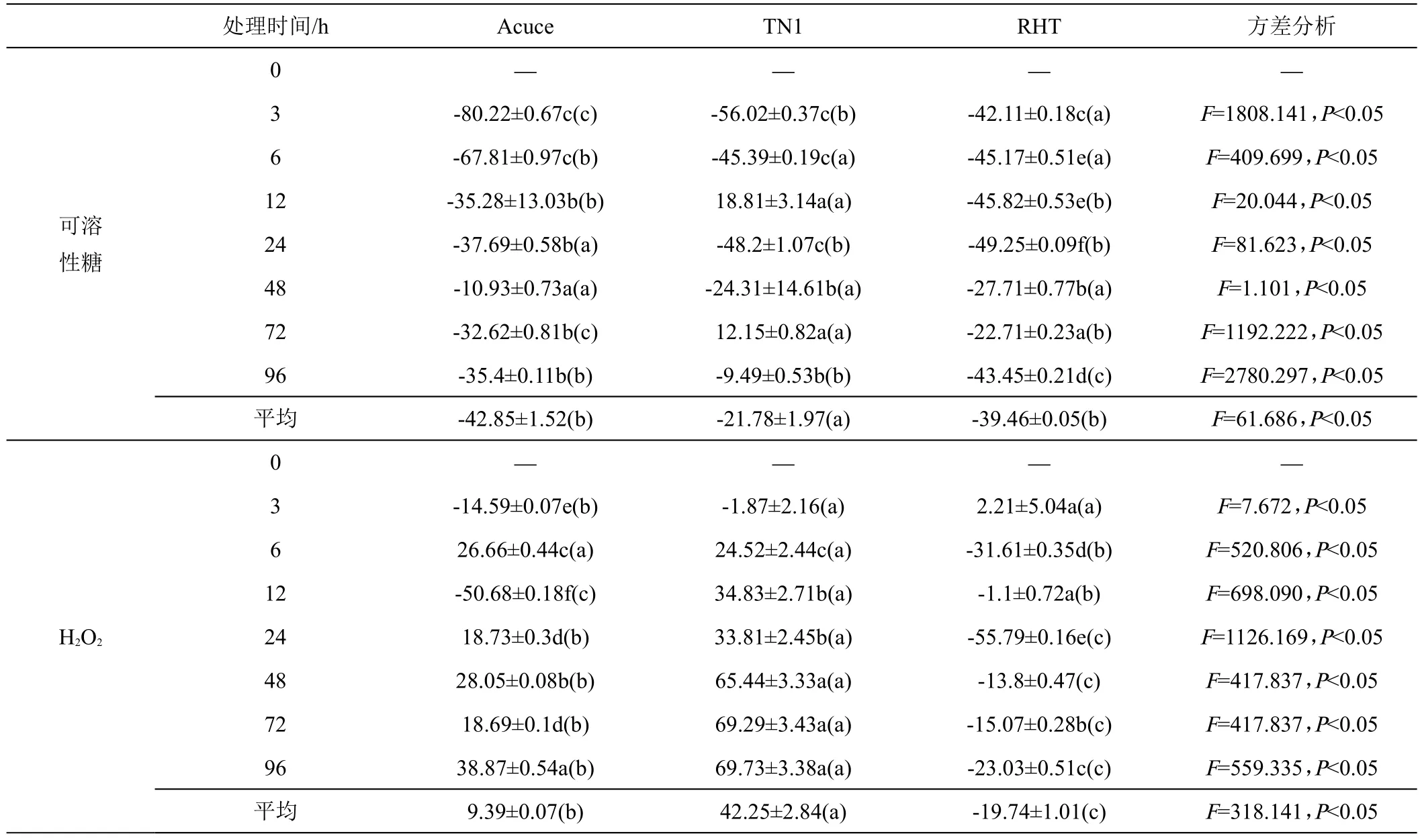
表1 褐飞虱为害后水稻体内可溶性糖、过氧化氢和丙二醛含量较对照的增降率%

续表1
2.2 褐飞虱为害后水稻过氧化氢含量的变化
由图2可看出,褐飞虱取食胁迫3 h后,‘月亮谷’植株内H2O2含量降低,取食6 h后,H2O2含量上升,取食12 h后,‘月亮谷’H2O2含量显著下降,取食24 ~96 h后上升至高于对照水平,取食96 h达峰值10.29 U/gFW;褐飞虱取食3 h后,‘TN1’的H2O2含量略下降,取食6 ~48 h上升,取食48 ~96 h后H2O2含量基本不变并高于对照水平,取食96 h为峰值10.04 U/gFW;褐飞虱取食胁迫6 h后,‘RHT’的H2O2含量显著下降,在取食12 h后显著上升且接近对照水平,取食24 h后显著下降,取食胁迫48 h后显著上升,至取食72 ~96 h后H2O2含量缓慢下降,取食6 h为峰值8.15 U/gFW,且与对照无显著差异。
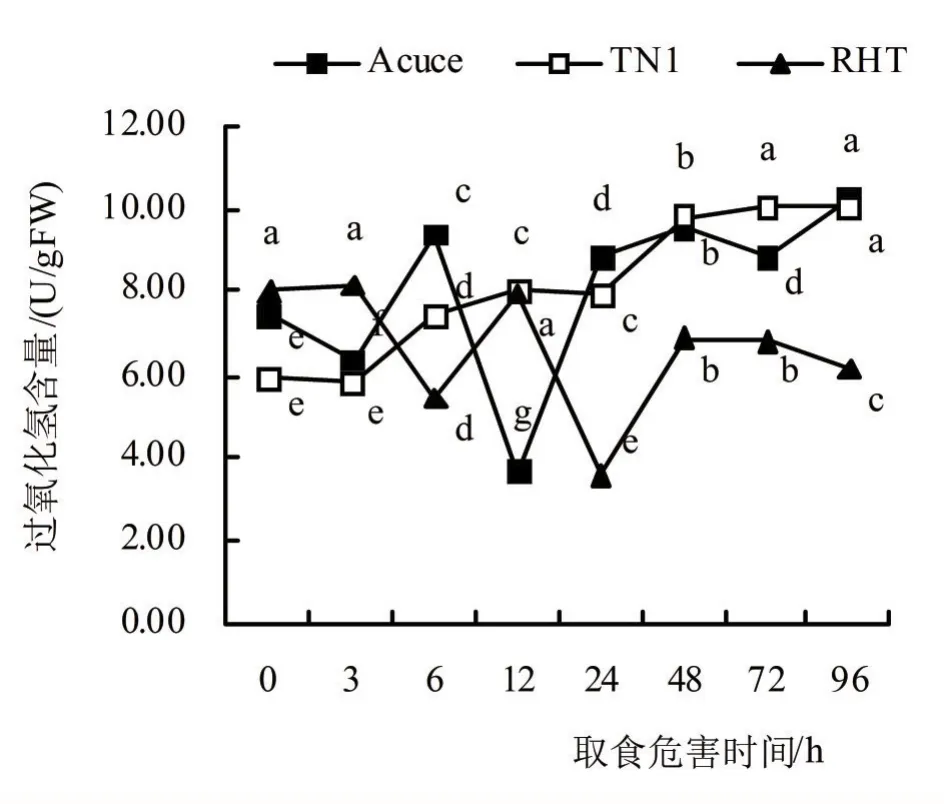
图2 褐飞虱取食胁迫后水稻体内过氧化氢含量
从褐飞虱取食危害后供试水稻植株内H2O2含量增加率来看(表1),取食3 ~96 h后,‘月亮谷’、‘TN1’和‘RHT’体内H2O2含量相较于对照分别增加9.39%、42.25%和-19.74%,由此看出‘月亮谷’体内的H2O2平均含量增加率高于抗虫品种‘RHT’,但显著低于感虫品种‘TN1’。
2.3 褐飞虱为害后水稻丙二醛含量的变化
由图3可知,褐飞虱取食胁迫3 h后,‘月亮谷’植株内MDA含量下降,在取食6 ~24 h后上升并保持相近水平,取食48 h后显著下降至最低值2.91 nmol/gFW,取食72h后上升并于取食96h上升至峰值6.34nmol/gFW;褐飞虱取食6 h后,‘TN1’的MDA含量显著下降至最低值0.89 nmol/gFW,取食12 ~72 h后上升至峰值5.90 nmol/gFW,取食96 h时下降;褐飞虱取食3 ~6 h后,‘RHT’的MDA含量迅速增加,取食24 h后,‘RHT’的MDA含量显著下降至最低值4.26 nmol/gFW,在取食48 h后上升,且取食胁迫72 ~96 h基本不变。
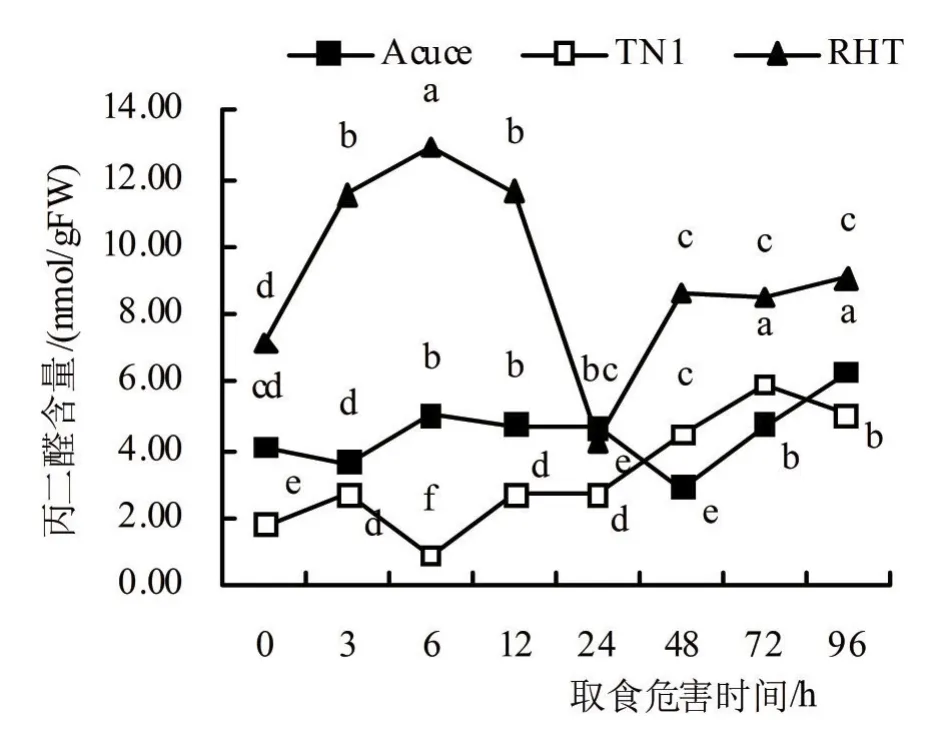
图3 褐飞虱取食胁迫后水稻体内丙二醛含量
从被褐飞虱取食胁迫后水稻植株内MDA含量的增降率来看(表1),被取食胁迫3 ~96 h后,‘月亮谷’、‘TN1’和‘RHT’植株内的MDA含量相较于对照分别增加了10.77%、87.81%和32.31%,‘月亮谷’植株体内MDA含量平均增加率显著低于‘TN1’和‘RHT’。由此表明,褐飞虱取食胁迫后,‘月亮谷’稻株内MDA含量较感虫品种‘TN1’和抗虫品种‘RHT’的更低,且抗虫品种‘RHT’的MDA含量平均增加率显著低于感虫品种‘TN1’。
2.4 褐飞虱为害后水稻多酚氧化酶活性的变化
由图4可知,褐飞虱取食胁迫3 ~12 h后,‘月亮谷’植株内PPO活性缓慢上升,取食24 h后时上升至第一峰值7.91 U/gFW,而在取食48 h后显著下降,取食72 h显著上升达峰值8.89 U/gFW后又下降;褐飞虱取食胁迫3 h后,‘TN1’的PPO活性迅速增加,并于取食6 h上升至峰值19.47 U/gFW,取食12 h后显著低于对照,取食12 ~48 h后上升达峰值19.47 U/gFW,取食72 h下降后又上升;褐飞虱取食胁迫3 h后,‘RHT’的PPO活性显著下降,在取食6 h后迅速降至接近对照水平,取食24 h后下降至最低值2.67 U/gFW,取食48 h后增加至峰值14.49 U/gFW,取食72 ~96 h后缓慢下降。
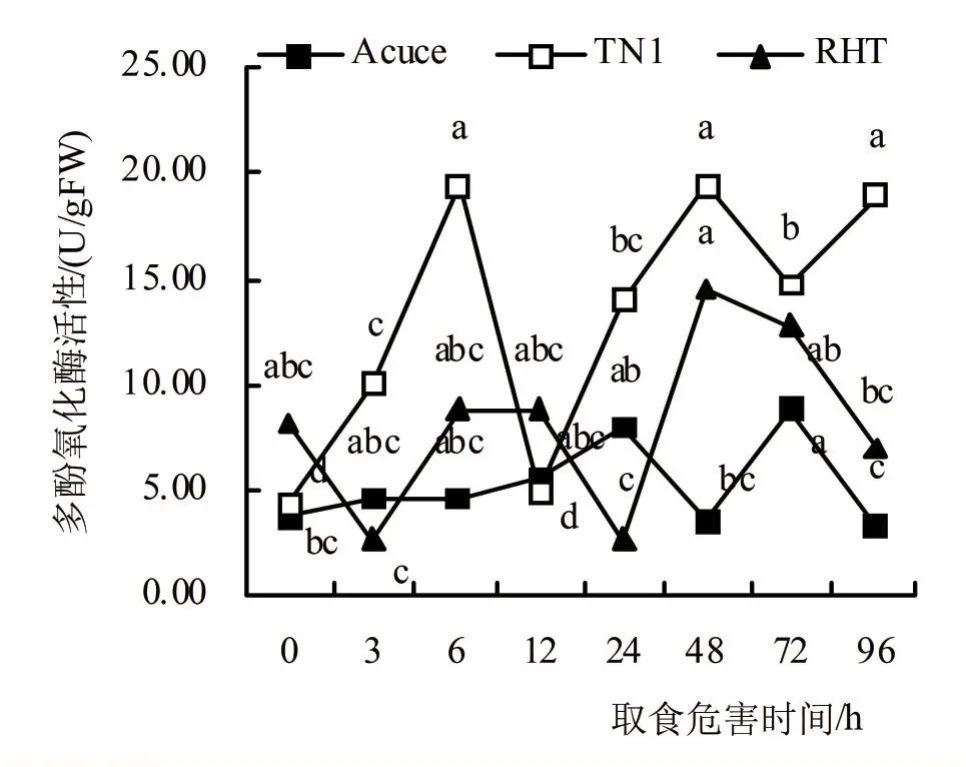
图4 褐飞虱取食胁迫后水稻多酚氧化酶活性
由表2可知,褐飞虱取食胁迫3 ~96 h后,‘月亮谷’、‘TN1’和‘RHT’的PPO活性相较于对照的平均增加率分别为49.96%、241.87%和0.89%,表明‘月亮谷’体内PPO活性平均增加率显著低于感虫品种‘TN1’,而与抗虫品种‘RHT’间无显著差异。

表2 褐飞虱为害后水稻体内PPO和POD活性较对照的增降率 %
2.5 褐飞虱为害后水稻过氧化物酶活性的变化
由图5可知,褐飞虱取食胁迫3 h后,‘月亮谷’POD活性显著下降,在取食6 h后又显著上升至峰值475.28 U/gFW,取食24 h后下降至最低值62.93 U/gFW,之后波动上升至高于对照水平;褐飞虱取食胁迫3 h后,‘TN1’的POD活性显著下降,在取食6 h后又显著上升至除对照外最高值220.80 U/gFW,取食12 h后下降,取食48 h下降至最低值23.70 U/gFW,取食72 ~96 h上升;褐飞虱取食胁迫3 h后,‘RHT’的POD活性显著下降,在取食6 h后又显著上升至峰值293.98 U/gFW,取食12 h后下降至最低值118.37 U/gFW,之后波动上升至高于对照水平。

图5 褐飞虱取食胁迫后水稻过氧化物酶活性
由表2可知,褐飞虱取食3 ~96 h后,‘月亮谷’、‘TN1’和‘RHT’的POD活性相较于对照的平均增加率分别为-14.75%、-45.01%和1.75%,表明‘月亮谷’体内POD活性平均增加率显著低于抗虫品种‘RHT’,高于感虫品种‘TN1’。
3 讨论
植食性昆虫从植物中获取生长的必要物质,昆虫的生长代谢直接受植物的营养物质含量影响,因此,植物通过降低营养物质含量,来达到抗虫目的[25]。段灿星等[26]研究报道,抗感水稻品种的可溶性糖含量在灰飞虱为害后均下降,且感虫品种下降幅度显著大于抗虫品种,而本研究中水稻被褐飞虱取食胁迫后,供试‘月亮谷’及抗虫和感虫品种稻株内可溶性糖含量均降低,但抗虫品种体内可溶性糖含量下降幅度显著大于感虫品种。虫害诱导后,植物中的可溶性糖转化为次生物质,降低了植物的适口性,植物内营养成分缺少,阻碍昆虫取食。本研究发现,褐飞虱取食后,水稻‘月亮谷’的可溶性糖含量较对照显著下降,且在取食为害后的多数时期可溶性糖含量低于对照,表明水稻植株通过减少稻株内的可溶性糖含量以响应稻飞虱的取食胁迫。此外,褐飞虱为害后,‘月亮谷’稻株内可溶性糖含量的下降趋势与抗虫品种‘RHT’相近,由此推测,‘月亮谷’与抗虫品种‘RHT’能通过降低可溶性糖含量来表现对褐飞虱的抗性。
害虫的取食危害胁迫能影响植物体内活性氧代谢系统的平衡,破坏膜结构[13]。MDA含量高低能够指示植物细胞膜发生细胞膜脂过氧化程度[27],也可用来表示植物抗逆境反应的强弱[28]。而H2O2是植物体内主要的活性氧,植物体内H2O2含量可作为昆虫对植物伤害强弱的生理指标[29]。本研究发现,褐飞虱取食胁迫后,‘月亮谷’植株内H2O2含量明显上升,显著高于抗虫品种‘RHT’体内H2O2含量,但却显著低于感虫品种‘TN1’。表明褐飞虱为害后,会诱导H2O2在水稻‘月亮谷’体内积累,增加至一定程度时引起水稻‘月亮谷’的防御反应;同时,水稻稻株内过量积累的H2O2使水稻过氧化而造成损伤,因此,感虫品种‘TN1’的MDA含量显著增加。
昆虫为害植物后,植物体内的防御酶就会被诱导或抑制,从而保护植物达到防御目的[30-31]。PPO是一类催化酚类化合物氧化为醌的酶,能够阻碍昆虫营养的利用,PPO的过量表达,一定程度上在植物虫害胁迫中起着重要的作用[33-34]。不同水稻品种的PPO活性均上升,且‘月亮谷’PPO活性增加率与抗虫品种‘RHT’无显著差异,表现出相似的防御反应。
过氧化物酶(POD)是一种在植物中广泛存在清除过量活性氧的氧化还原酶,同时也是植物抗逆反应过程中的关键酶之一,在一定程度上使植物抵御逆境胁迫伤害[32]。本研究中,云南哈尼族地方传统水稻‘月亮谷’和感虫品种‘TN1’受褐飞虱为害后,其稻株内防御酶POD活性反而下降,而抗虫品种‘RHT’的POD活性略增加,这与前人结果相反[16]。受褐飞虱取食为害后,不同水稻品种的不同防御酶协同作用,不同试验处理及试验方法对试验结果会产生较大的影响,产生不同甚至相反的结果,需要进一步的研究。
本研究中的云南地方品种‘月亮谷’仅在云南地区进行种植和试验,适用范围较窄,对全国范围内不同气候的适应性不明确,需要进行更大范围的试验点种植及完善的试验。而且本研究中仅仅进行了3个水稻品种的三叶期褐飞虱成虫取食胁迫试验,未进行大量水稻品种、不同水稻生长时期与褐飞虱不同时期虫态之间的取食试验;试验的水稻株数与单株水稻上的取食褐飞虱数量均较少,其试验结果存在偶然性,在今后应进行更加系统的试验。水稻‘月亮谷’对褐飞虱的抗性机制有待深层次的挖掘研究。
4 结论
本探究发现褐飞虱取食胁迫后,‘月亮谷’稻株内可溶性糖含量下降42.85%,与抗虫品种‘RHT’(-39.46%)无明显差异,而显著小于感虫品种‘TN1’(-21.78%);H2O2含量增加9.39%,低于感虫品种‘TN1’42.25%)而显著高于抗虫品种RHT(-19.74%);MDA平均含量增加10.77%,增加率显著低于‘TN1’(87.81%)和‘RHT’(32.31%);多酚氧化酶(PPO)平均活性增加49.96%,过氧化物酶(POD)活性下降,其中PPO平均活性增加率(49.96%)与‘RHT’(0.89%)无明显差异,而显著低于‘TN1’(241.87%);POD平均活性下降增加率(-14.75%)均显著低于‘RHT’(1.75%)但高于‘TN1’(-45.01%)。云南哈尼族地方水稻品种‘月亮谷’通过试验测定的各项生理指标均与抗虫品种‘RHT’反应较相似,具有良好的防御系统应对褐飞虱的取食为害,云南地方水稻品种‘月亮谷’可作为良好的抗褐飞虱育种材料应用于水稻生产。
猜你喜欢
中国农业科学(2022年12期)2022-06-28
热带作物学报(2022年5期)2022-06-06
热带农业科学(2022年5期)2022-06-02
江西水产科技(2022年2期)2022-05-17
农业工程学报(2022年1期)2022-03-25
河南科技(2022年1期)2022-03-11
载人航天(2021年5期)2021-11-20
农业与技术(2019年6期)2019-05-29
江苏农业科学(2017年19期)2017-11-22
农家顾问(2016年7期)2016-05-14
