
基于保护性农业的土壤固碳过程研究进展*
2022-04-14 00:16:20徐英德
中国生态农业学报(中英文) 2022年4期
徐英德
(沈阳农业大学土地与环境学院/土肥高效利用国家工程研究中心 沈阳 110866)
土壤有机碳(SOC)循环是驱动土壤肥力和生态服务功能实现的重要媒介, 识别SOC的固定路径及驱动机理属于土壤学核心理论范畴。然而, SOC的来源、固定过程和稳定机制仍存在较大争论。随着现代土壤分析技术的进步, SOC的研究方法逐渐趋向于原位、可视化和生物标识物等更真实反映碳赋存、周转动态的手段; 对于SOC固定的认识逐渐由腐殖化理论转变成以微生物代谢为核心驱动力的固碳过程; 有机碳分子组成主导SOC稳定性的作用减弱, 而物理、化学保护作用日益凸显; 通过微生物体内合成代谢产生的死亡残体在SOC截获中占据的重要份额被逐渐认可。新的理论发现极大地推动了SOC固定研究新一轮的认知革命, 同时也为构建SOC固定-土壤环境变化-土壤功能实现之间的关联带来新的挑战。
在所有陆地生态系统中, 农田生态系统因结构简化和人类活动制约而变得更为脆弱, SOC储量也极易受人为扰动影响。随着世界人口的不断增长,保障粮食安全成为全球范围内共同关注的焦点。据估算, 粮食产量在2050年必须增加60%才能满足人类需要。然而, 当前农业的发展在很大程度上还依赖于农资的投入而不是效率的增加。在人为垦殖的催化作用下, 大量SOC被氧化, 温室气体排放增加, SOC库和大气碳库之间的平衡被破坏; 同时,长年翻耕、过量施肥和农机使用等不合理的农作措施很容易导致土壤侵蚀、退化和结构破坏, 并将土壤由碳汇转为碳源。目前, 世界范围内已有多达25%的农田土壤严重退化, 这迫使我们重新审视耕作在农业可持续发展中的意义, 其中的核心任务就是SOC储量的稳定与提升。《联合国气候变化框架公约》第21次缔约方大会宣布启动“千分之四全球土壤增碳计划”, 旨在通过增加SOC含量实现保障粮食安全与减缓气候变化双重目标。无疑, 破解当前困局并将“千分之四计划”落地的关键就是寻求生产高效且使农业与环境充分兼容的农田管理方式。
近年来, 保护性农业(conservation agriculture)措施为世界粮食生产带来一场“耕作革命”, 并被越来越多的国家所采用。保护性农业通过减少土壤扰动,增加地表覆盖以及综合的养分管理等措施减少土壤侵蚀的风险, 提高土壤固碳速率, 保持土壤水分, 改善土壤健康状况, 增强土壤应对气候变化的能力并保障农田可持续发展。保护性农业措施可以通过物理、化学和生物等不同过程调控SOC的固定水平, 最终影响到不同生态系统中碳收支的权衡。在当前SOC固定理论发生重大变革的机遇下, 有必要在厘清保护性农业措施对SOC含量影响的基础上,进一步探讨其介导土壤固碳过程的机理, 这对于明确农田土壤碳增汇过程, 完善农田生态系统碳循环模型意义重大。基于以上背景, 本文对近年来SOC固定这一关键生物地球化学过程及保护性农业对SOC固定的调控等研究进行了综述: 总结了SOC固定途径和稳定过程新的进展与面临的挑战, 分析了传统耕作与保护性农业措施对SOC固定的影响, 剖析了保护性农业驱动SOC固定的机制, 最后对相关研究领域的发展进行了展望。
1 土壤有机碳的固定途径
SOC库是陆地表面最大的有机碳库, 全球土壤数据整合计量表明, 深度为1 m内的SOC储量为1505 Pg, 约是植被有机碳储量的2.43倍, 大气碳储量的1.71倍; 深度为2 m内的SOC储量超过2000 Pg(图1)。鉴于以往对SOC储量的评估缺乏地形和山坡斜率的考虑, SOC的储量甚至被低估。因此, SOC储量的微小变化就会对大气CO浓度产生强烈影响(1 Pg SOC=0.47 mol·LCO)。同时, SOC的流动能够占到全球陆地碳循环总量的80%。因此, SOC的固定及其人为管理在调节全球气候变化和农田生产力上具有极为重要的意义, 其固定过程与调控机理也一直备受关注, 并出现百家争鸣的局面。
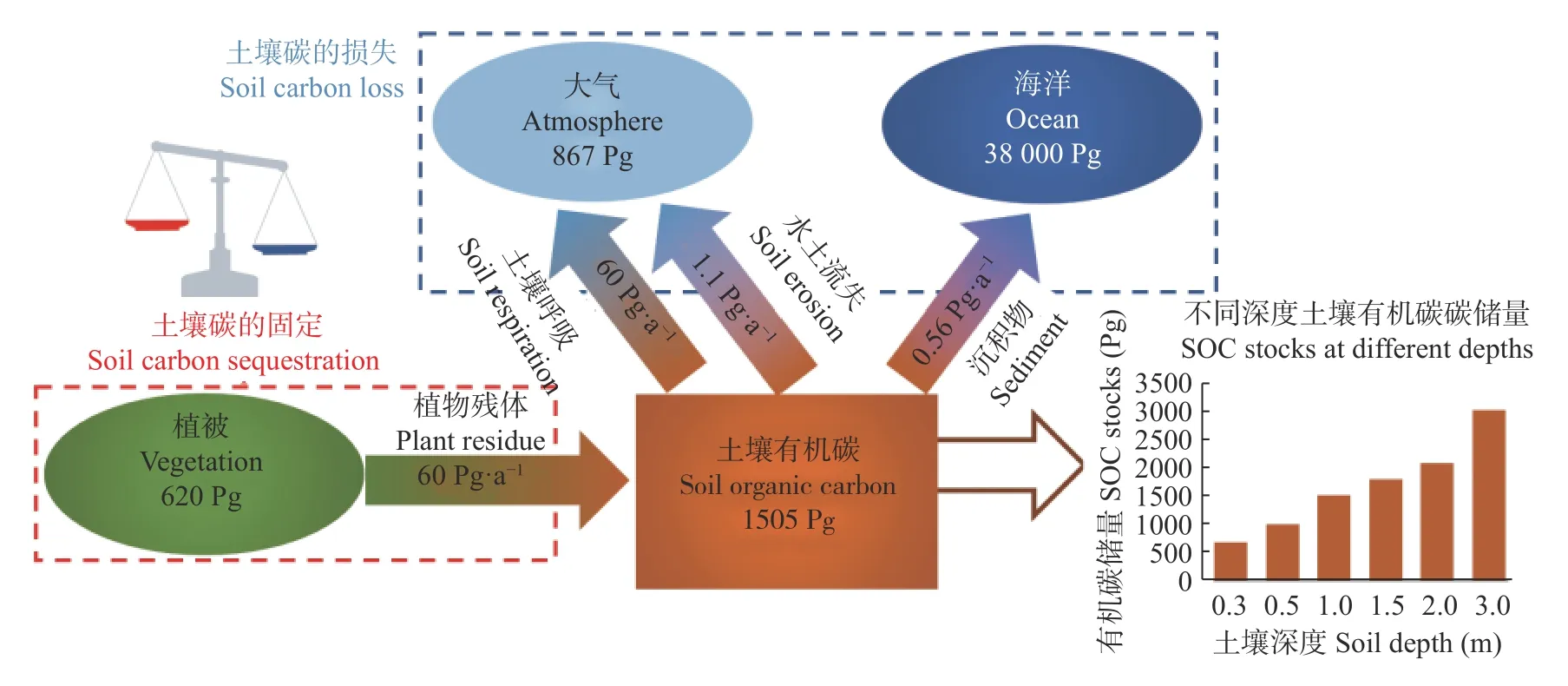
图1 全球碳收支模型[16-18]Fig.1 Contemporary global carbon budget[16-18]
在全球范围内, 面对愈发严峻的碳中和及粮食安全的压力, 土壤固碳研究在近几十年发展迅速(图2)。一方面, 由传统的化学提取等研究方法所得到的“盲人摸象”结果演变为原位高分辨率可视化和微生物作用机理趋于清晰的有机碳固定、转化过程; 另一方面, 有机碳作为土壤乃至整个地球生态系统中的基本属性与驱动力, 其在全球治理服务及政策管理中扮演的角色越发重要。目前从SOC的赋存形态到形成途径均产生了较大变革, 逐渐由酸碱浸提为基础的腐殖质相关学说向生物标志物为基础的有机质分子组学转变, 其中微生物对土壤固碳起到的推动作用逐渐成为共识。
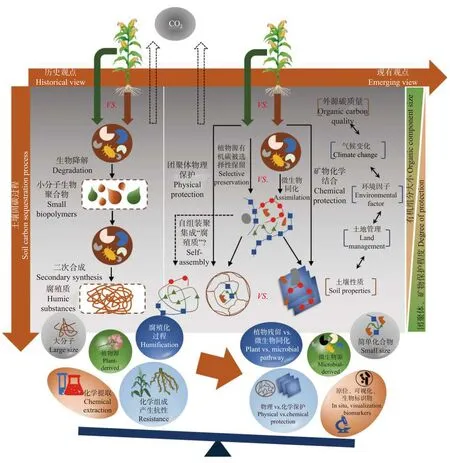
图2 土壤固碳机制示意图Fig.2 Schematic diagram of soil carbon sequestration mechanism
在经典土壤学中, 输入到土壤中的未分解和半分解的动植物残体和腐殖质被认为是SOC的主要赋存形态。由新鲜有机质矿化-残余物二次合成-聚合与缩合等过程(即腐殖化过程)形成的分子量大、具有高度异质性的长链物质——腐殖质是SOC库的主体。其中, 有机质分解后的中间产物与含氧芳香烃的聚合过程是腐殖质学说的根本。事实上, 至今尚未有充足的直接证据证明“二次合成”过程的真实存在性, 腐殖质的真实分子结构也存在模糊性。但最近也有研究通过C核磁共振(NMR)和高分辨质谱得到了腐殖质存在和木质素起源学说的新证据,这为SOC赋存形态的争论带来了新的视点。除土壤腐殖化过程外, 选择性残留学说也是SOC固定途径的重要理论; 该理论认为SOC是微生物对外源有机质完成分解后选择性保留在土壤中的结果, 其中易分解组分会快速被微生物降解, 而那些顽固组分则会残留在土壤中构成稳定有机碳库。在此基础上,一些新的研究认为土壤稳定有机碳库多是由相对较小的短链小分子化合物构成, 如多糖、多肽和木质素碎片等; 而整个SOC库则是从大的植物生物聚合物到小分子化合物的连续体。
在历史观点中, 腐殖质的形成与木质素密切相关, 加之选择性残留学说也认为植物源有机组份的选择性保留形成SOC, 因而植物源有机碳是构成SOC库的主力军。但是, SOC的固定离不开微生物的驱动, 土壤微生物可以通过分解代谢和合成代谢这2个相反的途径分别将SOC释放到大气或将微生物来源碳贡献于SOC库。但在传统的土壤固碳理论中, 大多数研究只关注到了前者。越来越多的研究证实微生物通过周而复始的同化过程使其死亡残体和代谢产物不断地在土壤中累积并趋向稳定化(即续埋效应)。因此, 微生物源有机碳在SOC库中的权重可能被远远低估; 同时, 以微生物合成代谢为核心的土壤固碳过程成为了近期关注的焦点。综合上述观点, 从微生物驱动的角度, 土壤的固碳途径可能同时存在被微生物选择性保留植物源有机碳组分的过程, 以及被微生物同化合成微生物源有机碳组分的过程。但需要强调的是, 随着SOC库主要由小分子构成的观点被逐渐接受, 很多研究同样发现腐殖质是通过小分子物质通过疏水力和氢键等弱相互作用自组装聚集而成。这为SOC的形成机理带来新的挑战: 植物残体和微生物残体的分解产物能否进一步构成具有新内涵的“腐殖质”?这种“腐殖质”中微生物源和植物源的占比如何?
伴随着土壤固碳途径观点的演变, 土壤的储碳机理也逐渐趋于清晰, 但也出现了新的争议点。SOC的稳定性取决于有机碳和土壤环境因子的相互作用。在传统腐殖质学说中, 腐殖质复杂的化学结构和疏水性使其具有了较高的化学稳定性, 并与土壤矿质胶体通过金属离子桥键结合的方式进一步提高了其抗分解的能力。但腐殖质的化学稳定性只是相对的,假设有简单有机分子穿插在腐殖质的疏水缔合结构中, 其稳定性便会被破坏; 在适宜的条件下, 微生物对大分子有机物的分解速度也可以很快; 并且, 其与矿物结合的过程并不符合热力学降低的规律。由于传统提取腐殖质方法的弊端, 提取出的大分子有机质可能存在假象缩合的现象, 从而掩饰了SOC的真实形态。此外, 伴随着土壤微生物固碳学说的发展, 有机质自身内源性属性的差异(如植物残体质量)对土壤固碳过程的影响受到了争议。例如, 由木质素和软木脂等组成的植物根茬不易被微生物降解,被认为是稳定SOC库的重要前体; 然而, 微生物对植物茎叶有更高的碳利用效率, 从而使其更高效地以微生物残体的形式贡献于SOC库。新的证据表明植物残体腐解初期主要通过可溶性有机碳-微生物同化途径形成矿物结合态有机碳, 而在腐解后期植物残体则通过物理转运途径形成颗粒有机碳。这说明有机质的质量或分子组成仍是介导SOC固定的重要因素, 尤其是通过影响微生物的同化/异化过程调控SOC的形成路径, 但不同分子组成有机质在微生物同化和选择性保留途径的中的分配和权衡尚待商榷。尤其是随着时间的推移, 那些被选择性保留的植物源有机碳仍然可能被微生物利用, 这进一步增加了相关科学问题的复杂性和厘清的必要性。
至今为止, 虽然众多研究试图从有机碳分子组成角度摸索SOC的稳定机制, 胡敏酸、木质素、芳香碳或烷基碳等组分或分子的多寡常被用来预测SOC的稳定性。但近期也有很多研究更倾向于认为有机碳化学结构本身对SOC稳定性的作用被高估, 起码结构或分子的差异不起决定性作用。例如糖类和氨基酸等碳组分也可以作为稳定SOC储存,而木质素在土壤中的周转时间却可能很短。而SOC的真实稳定性取决于其被团聚体和矿物保护的情况(即空间上的隔离和生物化学上的难分解), 或者说取决于土壤生物的可及性。这些理论为揭示SOC固定机理提供了新的视角和思路。首先, 土壤中的水溶性碳并非意味着易分解碳, 这些碳一方面容易被吸附于矿物表面, 还可以被微生物同化固定, 从而通过不同的化学-生物途径组成土壤中的“老”碳。其次, 团聚体的物理保护与矿物的化学保护之间同样存在权衡关系, 例如颗粒有机碳虽然没有受到矿物的结合, 但同样可以通过团聚体的物理隔离而成为持久固存的“老”碳。潘根兴等主张之前所认同的化学结构复杂有机碳的抗降解性, 实际上也来源于团聚体、矿物和微生物构成的微结构对这些有机碳的保护。但值得注意的是, 有机碳自身化学组成对土壤固碳的影响不应被全盘否定, 尤其是在外源有机碳输入土壤的初期, 这些有机碳自身性质深刻影响着不同组分在微生物-团聚体-矿物中的归趋, 进而调控土壤的储碳过程。
总体来看, 有机碳化学组成、微生物、有机分子间的凝聚、团聚体和矿物在土壤固碳及稳定过程中扮演的角色不能被割裂。微生物的同化/异化、团聚体的周转和有机无机复合过程均是同步进行的,他们之间既相互协调, 又相互制约, 最终不同分子组成的外源有机碳在各条土壤固碳途径之间的权衡造就了不同区域土壤微结构、微生物区系和SOC固存模式的多样性。此外, 除了SOC周转的自身过程、外源有机碳的质量和土壤因子外, 日益严峻的全球气候变化和环境因子也成为深刻影响SOC固定的重要推手, 这些因素不仅能改变外源有机碳输入的数量、质量和地理分布格局, 还能够调控微生物的活性与群落组成。尤其是在农田土壤中, 还要将人为扰动纳入到SOC固定模型中; 土地利用方式、耕作模式和农田管理措施等对SOC固定的影响会远远超过自然变化影响的速率和程度。
2 传统农作方式造成土壤有机碳的损失
在人类社会发展过程中, 由自然生态系统向人为管理生态系统的转化会消耗SOC存量。人类活动影响了约40%的地球表面, 将近92%的天然草地/草原已转为人类使用, 包括放牧和农田。平均而言,将天然草地转化为作物生产导致了约50%的有机碳损失。利用这些土地利用变化和有机碳消耗量的数据, 估算出历史上因土地利用方式变化而造成的SOC损失估计为115~154 Pg。将泥炭地转换土壤利用方式后, SOC储量的相对损失比矿质土壤更严重。农业耕作能加剧土壤侵蚀, 造成SOC的氧化以及土壤结构质量的下降。早在19世纪30年代, 美国因沙尘暴的爆发对耕作的意义进行了大讨论。特别是进入20世纪60年代以来, 人类通过改良作物品种、施用化肥和除草剂、灌溉和耕作机械化等农业管理方式的改良, 推动了世界范围内粮食产量的飞跃。但这背后所隐藏的是环境污染、土壤侵蚀和养分耗竭、水资源枯竭等问题。政府间气候变化专门委员会(IPCC)统计数据显示, 近几十年间, 大气CO浓度已经从原来的316 mol·L上升到400 mol·L以上, 这很大程度上归咎于SOC的释放。Lal估算表明, 在2007-2016期间, 由于人为的土地利用方式转换产生的碳排放量年均约1.3 Pg。在我国, 耕作土壤的有机碳损失量估计为15 Mg·hm; 在北方半干旱的坝上地区, Zhao等报道, 经过50年的草地开垦, SOC减少幅度为73%~79%; 东北黑土区是我国重要的商品粮基地, 由于长期高强度重用轻养等原因, SOC含量在开垦最初20年下降了约30%, 40年后下降约50%, 70~80年后下降约65%, SOC储量的降低导致黑土越来越瘦。
以犁耕为代表的传统耕作方式主要通过引发土壤侵蚀和水土流失而影响SOC固定过程。集约化耕作使土壤疏松并裸露, 为土壤结构和有机质被风、雨和流水等外力破坏创造了有利条件。在这些外力的作用下, 土壤团聚体发生破碎, 团聚化过程受到阻碍, 从而使包被在团聚体中的有机碳更易接触到微生物, 进而被分解。因此, 加速的土壤侵蚀是大气CO的重要来源。在全球范围内, 因水蚀产生的SOC排放量约为1.1 Pg·a。侵蚀在加速碳损失时, 对不同SOC组分的影响是不一致的; 例如, 颗粒有机碳更容易通过水蚀作用而分解, 但矿物结合碳则保留时间较长; 土壤活性有机碳在好氧和厌氧条件下会分别更倾向于通过地表径流迁移和分解释放为CO。因此, 传统耕作模式-侵蚀-SOC损失途径会受到土壤特性、微气候和管理措施等多种因素的影响。对于我国而言, 干旱、半干旱耕地所占比例较大, 尤其是在北方地区, 这在一定程度上加速了土壤的退化和SOC的矿化。此外, 长期秸秆焚烧导致有机物料不能及时归还土壤; 长期耕地粗放管理导致土壤固碳能力不足; 以小四轮拖拉机为主要动力的犁耕方式导致耕作层越来越薄; 不合理、过量施肥导致土壤板结、酸化; 人口增长导致粮食需求量越来越大, 这些都成为SOC流失的重要推动因素。因此, 建立合理的农业耕作措施刻不容缓, 对提升SOC储量和肥力水平意义重大。
3 保护性农业的内涵
3.1 保护性农业的发展及其概念的标准化
保护性农业是美国在20世纪30年代由于大规模开垦和过度耕作所引发的黑风暴、干旱、土壤退化、疟疾泛滥等资源环境约束背景下发展起来的一种现代农业技术, 目前已成为很多发达国家主推的可持续性耕作制度。在黑风暴之后, 引起美国政府和民众对传统耕作方法的反思和对水土保持新方法的探索。20世纪40年代以来, 人们逐渐意识到有必要取消传统耕作, 并在地表保留作物残茬覆盖物, 由此产生了一系列新的相关概念, 如免耕(no-till)、零耕作(zero tillage)、直接播种(direct seeding)、保护性耕作(conservation tillage)、少耕(minimum tillage)、覆盖耕作(mulch tillage)和条带耕作(strip tillage)等,这些概念的发展也伴随着对保护性农业定义的演变(图1)。如果不加区别地使用这些概念, 会加剧该农业管理实践的混淆和争议。因此, 对保护性农业的概念进行标准化是非常重要的。
保护性农业是在免耕的基础上发展起来的, 但也有很多研究认为免耕技术在减少水土流失的同时,也存在很多弊端, 如提高杂草和病虫害数量、提升土壤压实的风险、NO排放增加和肥料利用率降低等。由于这些局限性和不确定性, 重点应将保护性农业作为一个在免耕的基础上发展起来的综合系统进行考量, 而保护性农业产生的最终效果要取决于系统中不同组件之间的互联性及协调性。基于这个理念, Lal认为保护性农业的定义是在一个轮作周期内由作物残体覆盖(residue mulch)、种植覆盖作物(cover crop)、免耕(no-till)和养分综合管理(integrated nutrient management)等技术组成的耕作系统,其目的是有效地保持水土、固碳、适应和缓解气候变化并实现农业的可持续发展(图3)。保护性农业的目标并非是谋求最高的产量, 而是可持续的最佳产量, 尤其是在作物生长状况较差的季节是否能保持稳定可观的产量, 这也体现了其气候适应性。
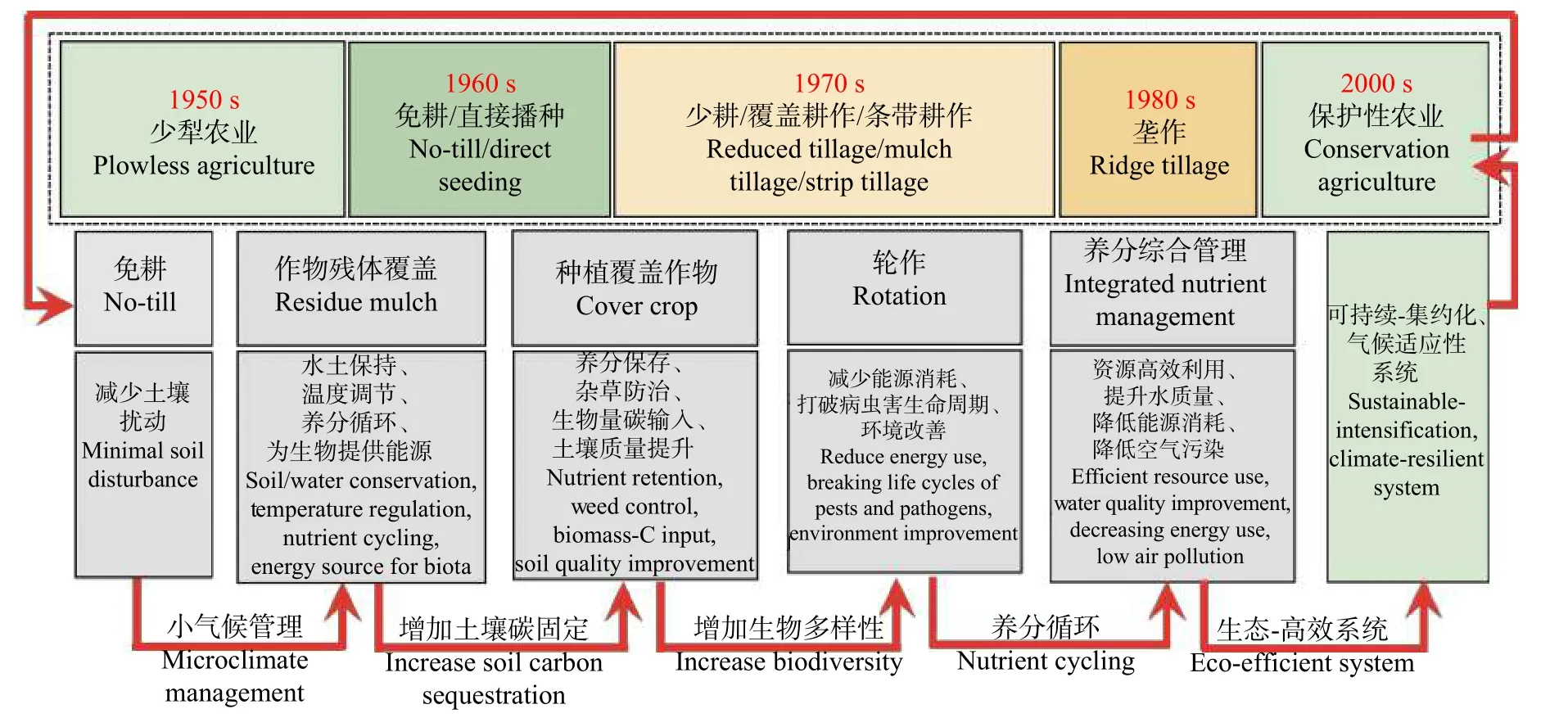
图3 保护性农业的历史演变与主要内容Fig.3 Historical evolution and main contents of conservation agriculture
3.2 保护性农业主要内容
免耕: 在收获后至播种前不搅动土壤, 以减轻风蚀和水蚀, 并采用联合作业的免耕播种机播种。播种机前部装有切刀, 在不拖移地表残留物的前提下,开沟播种、覆土和镇压, 一次完成作业。施肥可与播种同时进行或分期进行。
作物残体覆盖: 移除作物残体后对土壤性质及粮食生产力的不利影响已被广泛报道。而对半干旱地区土壤实行免耕措施之后, 如果土壤没有作物残体覆盖层, 将会大大降低农作物的产量和固碳潜力。作物秸秆盖土、根茬固土对于保护土壤和水分, 加强团聚化过程, 创造正的SOC收支, 改善土壤动物和微生物的活性和多样性, 最终促进养分循环利用以及维持粮食产量至关重要。
种植覆盖作物和轮作。种植覆盖作物的农业管理措施可以追溯到我国周朝时期, 但自1960年以来,由于化学肥料的廉价高效, 覆盖作物被逐渐舍弃。进入21世纪, 由于覆盖作物在环境和农业可持续发展方面的重要作用而被纳入到保护性农业系统中。种植覆盖作物可为作物保存养分, 减少水土流失, 缓解免耕造成的土壤压实, 并抑制杂草, 从而提高作物产量。此外, 轮作能够通过在季节间或年度间轮换种植不同作物达到均衡利用土壤养分, 减少病虫害并改善土壤理化性质的目的。
养分综合管理。充足的养分是维持土壤肥力与作物生长的必备要素。为保持农田生态系统的最佳生产力和可持续发展, 应采取必要的人为施肥调控策略, 建立有利元素(如氮、磷、钾和微量元素等)之间的平衡。同时, 养分综合管理措施也可以有效解决免耕对植物营养吸收带来的不利因素。需要强调的是, 土壤养分还有很大一部分来自植物、环境和自身有机质分解, 因而施肥措施必须建立在遵循不同体系之间养分转化、迁移和利用过程内在联系的基础上, 并将施肥同其他保护性农业措施以及环境因素(降水、温度等)融合考量, 以实现养分高效利用、环境友好与作物高产的目标。
4 保护性农业介导下的土壤固碳过程
4.1 保护性农业对土壤碳固定的影响
南美洲的保护性农业采用率在全球范围内占领先地位, 尤其是通过轮作与种植覆盖作物的结合普遍促进了SOC的固定。Sá等在巴西通过22年田间试验表明, 处于保护性农业措施下的土壤固碳速率为806 kg(C)·hm·a(0~20 cm)和994 kg(C)·hm·a(0~40 cm); Bayer等报道显示在砂质壤土和黏土中实行保护性农业措施, 土壤固碳速率分别提升0.30 Mg(C)·hm·a和0.60 Mg(C)·hm·a。此外, Sisti等观察到免耕只有与覆盖作物结合时, 才能提高土壤的固碳效率, 这体现出了保护性农业措施的系统协同特征。美国也对保护性农业进行了大范围的推广和研究。Allmaras等在分析美国长期田间试验数据的基础上, 报告了SOC储量的大小顺序为免耕>非犁耕>犁耕。总体上, 世界范围内对保护性农业提升土壤固碳速率的报道不胜枚举, 即使在季节性干旱的农业生态系统中, 保护性农业也起到了改善SOC储量的效果。我国虽然在保护性农业方面的实践起步较晚, 但关于其对SOC固定影响的研究也不断增加。李景等研究发现, 免耕秸秆覆盖和深松秸秆覆盖均提高了大团聚体有机碳含量, 且有机碳含量随耕作年限增加; 武均等在陇中黄土高原旱作农田不同耕作试验发现, 免耕和秸秆还田均较传统翻耕提高了SOC含量, 且以免耕秸秆覆盖处理效果最优。王成已等整合分析我国长期定位试验表明, 保护性农业处理下旱田和水田SOC平均增幅达0.21 g·kg·a和0.21 g·kg·a。
虽然由传统耕作方式向保护性农业转变后有利于表层土壤固定更多有机碳的规律已被广泛证实, 但对于底层土壤(20~50 cm)的固碳效果还存在争议。一些研究发现采用保护性耕作措施后, SOC分布的层化现象更为明显, 即有机碳主要在表层富集, 而对底层SOC含量影响不大。这主要是保护性农业措施对SOC固定的滞后效应导致的。随着土壤环境的改善及深根作物的种植, 深层SOC可能会不断累积, 并形成相较于表层土壤更稳定的碳库。例如,Walia等发现采用免耕等保护性农业措施44年后1 m深度的SOC储量明显提升, 甚至超过了森林土壤。此外, 保护性农业对SOC固定的时间尺度效应仍无定论。有研究发现保护性农业措施在短期内对SOC影响不大或存在波动, 但也有研究计算得出, 任何一个农业生态系统的耕作方式改变, 在前期SOC都以稳定的速率积累, 后期碳积累速率逐渐下降, 最终达到碳积累速率为零的稳定状态。
值得关注的是, 保护性农业在一些情况下并不会产生最大的粮食产量, 也不会使土壤固定足够的碳以缓解气候变化。在保护性农业实施的初期, 由于杂草、害虫和病原体增加、还田秸秆C/N较高、养分失衡以及播种设备不适宜等问题, 粮食减产的现象时有发生。此外, 土壤固碳过程受到气候和土壤环境等多种因素的影响, 可能会制约保护性农业对固碳的提升效果。例如, Ludwig等并未观察到不同耕作条件下SOC库的明显差异, 但保护性农业措施可能会促进SOC库的再分配, 即增加表层SOC含量而降低犁底层SOC含量。同时, 在一些地区, 夏季休耕也对SOC储量产生了不利影响, 可以通过将休耕转换为种植豆科植物以达到最大的固碳效益。总体来看, 保护性农业有时需要通过牺牲短期的粮食产量来谋求农业生态系统长期的可持续性,特别是对于一些生态脆弱区。因此, 确定不同特定地区限制保护性农业增产、固碳壁垒的原因极为重要; 同时, 通过合理的布局、组合和优化保护性农业的各项措施, 以使其达到最优的粮食产量与固碳效应是当前的主要挑战之一。
4.2 保护性农业驱动下的土壤固碳机制
改善水土保持状况是保护性农业提升土壤固碳能力的主要优势。保护性农业减少了土壤耕作的频率和强度, 并将作物残体作为覆盖物保留在土壤表面, 可保护土壤免受雨滴的冲击而结块或结皮, 减少了径流、土壤侵蚀和SOC损失的风险, 增强了雨水渗透性, 使土壤结构稳定性得到恢复, 孔隙比例更加协调。研究表明覆盖作物使土壤保水性提高10%~11%。在此基础上, 保护性农业能进一步通过影响团聚体的形成与瓦解更加精细地调节SOC储量。土壤颗粒的团聚化过程通过固定有机碳来增加SOC库的数量和质量。Li等通过meta分析表明, 各种保护性农业措施均能有效地增加土壤团聚体的稳定特征, 从而为有机碳的保护奠定根基。秸秆还田和种植覆盖作物所引入的植物源有机质可以作为胶结物质参与到团聚体的形成过程; 同时, 土壤团聚体通过复杂的物理/化学空间隔离作用(水分、氧气和微生物等)增加对新输入有机碳的保护。此外, 丛枝菌根真菌菌丝及其产生的球囊霉素蛋白作为结合剂是提高团聚化过程的重要因素, 免耕和少耕则能显著增加丛枝菌根真菌浓度, 从而有利于团聚体和有机碳稳定性的提高。土壤团聚体对有机碳的固定主要取决于其粒级大小, 其中大团聚体中的有机碳更新较快且更易受到耕作措施的影响, 而微团聚体由于较强的化学吸附能力更有利于有机碳的长期累积。因此, 合理的耕作措施所构建的团聚体-有机碳良性周转关系有利于土壤固碳能力的提升。
土壤微生物是外源有机碳向稳定SOC转化的重要通道。大量研究表明, 保护性农业措施能够为微生物生存创造适宜的温度和水分条件, 并为微生物代谢提供丰富的碳源, 促进微生物量的生成, 进而实现活体微生物-微生物残体-稳定有机质这一迭代过程的持续运转。保护性农业措施在促进微生物繁殖的基础上, 还能通过影响微生物群落组成调控土壤固碳水平。真菌较细菌具有较高的碳氮比,对碳需求更高, 对新输入碳源的响应也更强。因此, 在秸秆还田或种植覆盖作物的条件下, 更有利于真菌的繁殖和累积; 同时, 保护性农业措施还可以通过改变土壤pH和团聚体通气性的方式促进真菌菌丝网络的形成, 而免耕减少了土壤扰动, 也有助于真菌菌丝的建立和稳定性的增加。一般而言,真菌残体在土壤中比细菌残体更稳定, 加之其较强的底物同化能力, 因而保护性农业通过真菌和细菌群落之间的权衡使土壤固碳进一步朝有利的方向发展。在真菌门类水平, 有报道显示免耕和秸秆覆盖降低了担子菌门(Basidiomycota)丰度, 增加了子囊菌门(Ascomycota)丰度, 其中子囊菌门在外源有机碳的转化过程中作用很大。此外, 微生物群落多样性较高的土壤中不同质量外源有机碳参与到微生物周转过程的程度更大。由于微生物群落多样性对土壤环境变化较为敏感, 有部分研究指出免耕等措施会降低土壤微生物群落多样性, 但更多研究表明轮作和覆盖作物等措施均有助于提高农田土壤微生物群落多样性, 秸秆还田则能提高土壤自养固碳微生物多样性。在改善微生物代谢能力的基础上, 保护性农业还对整个土壤食物网及蚯蚓等动物数量和物种多样性具有积极影响, 土壤动物的穴居活动促进了SOC向底层土壤的迁移, 提高了SOC的驻留时间。
除微生物量及其群落组成之外, 保护性农业还能够通过增加微生物可利用资源调控微生物代谢活性及其衍生的微生物源碳累积过程。土壤酶直接参与到SOC循环过程, 其活性能够反映微生物活性。相对于传统耕作, 免耕、秸秆还田和作物覆盖等保护性农业措施能提高与各类群微生物量密切相关的脲酶、蔗糖酶、碱性磷酸酶、过氧化氢酶和纤维二糖酶的活性。另外, 秸秆还田有助于提高微生物底物利用率、代谢功能和与之相关的CO固定基因丰度, 从而促进了SOC的有效累积。从微生物残体累积情况来看, 免耕、施有机肥和秸秆还田会有效促进土壤表层及各级团聚体中微生物死亡残体的数量, 提高土壤SOC库的质量与整体稳定性, 其中真菌残体提高的幅度更大, 但研究结果尚未达成一致。Ding等报道显示, 微生物残体的累积能够有效地参与到团聚化过程, 且优先在微团聚体中累积并达到饱和, 这有利于土壤SOC的形成和稳定。还有研究发现免耕也能增加深层土壤中真菌源氨基葡萄糖在黏粒中的富集, 这对于深层SOC库的稳定性提升具有重要意义。
植物生产力同样与SOC储量关系密切, 地上和地下生物量的产生均可以作为SOC形成的潜在来源。由于保护性农业土壤中较好的结构与养分供应关系,植物净初级生产力及其衍生的净生态系统生产力、净生物群系生产力和作物产量等均会有所提高, 为SOC固定创造重要前期基础。同时, 种植深根作物及秸秆深还田等措施有助于将根系分泌物和秸秆源碳输入到底层土壤, 这也是提高SOC储量的有效策略。鉴于底层土壤中较高的微团聚体、黏粒含量以及较低的氧气比例, 底层土壤中的SOC更易于与土壤矿物颗粒结合, 形成稳定的化学保护结构。
总体来看, 保护性农业措施可以通过减少SOC的矿化, 增加SOC来源, 调控土壤团聚化过程, 促进微生物量及其残体的累积, 改善微生物群落组成及代谢活性等多种方式协同作用于SOC固定过程(图4)。但值得注意的是, 保护性农业应该是集多种措施的互补性和整体性农业管理方案, 而长期使用单一的耕作措施会造成土壤的单向演变, 如长期免耕可能会增加容重, 不利于SOC的良性循环。因此, 恰当的保护性农业措施选择与组合是实现土壤最大固碳能力的关键所在。

图4 保护性农业驱动下的土壤固碳机制Fig.4 Soil carbon sequestration mechanism driven by conservation agriculture
5 总结与展望
正确认识并理解SOC的形成稳定机制及其与农田管理措施的关系是解决土壤退化以及应对全球气候变化的关键。尤其是在面临碳中和目标的压力下,提高土壤的碳汇功能至关重要。长期实践证明, 保护性农业有望成为破解土壤肥力下降与粮食需求增长矛盾的利刃。然而, SOC具有复杂性、易变性和相关理论体系多元化的特征; 保护性农业也是众多农田管理措施的综合体, 错综复杂且相互影响。在这种背景下, 保护性农业如何精确调控SOC循环过程仍然扑朔迷离、充满疑问。未来研究应在以下几方面加以深入:
1)明确保护性农业介导下SOC固定的微生物-团聚体-矿物协同作用机制。探索SOC的真实赋存形态是明确土壤固碳机制的根本。随着有机碳分组和分离技术的提升, 使准确地解析SOC组成、结构和丰度, 建立起SOC分子组学研究体系成为可能。此外, 微生物是SOC截获过程的关键驱动者。在耦合运用同位素技术和组学方法的基础上, 如何抽丝剥茧地精准识别出保护性农业介导下的微生物群落格局及其碳周转功能的关联是破解土壤固碳黑箱的重中之重。同时, 将活体微生物的代谢过程同微生物死亡残体累积的动态进行结合有助于解析SOC形成的微生物连续性参与机制。需要注意的是, 微生物虽然主导着SOC的形成与损失, 但有机碳真实稳定性的产生应归属于团聚体与矿物对其的物理闭蓄和化学吸附。因此, 明确不同空间格局下土壤颗粒对微生物来源碳和植物来源碳的保护机制以及微生物群落响应规律, 综合构建起SOC固定的微生物-团聚体-矿物协同作用框架将成为该研究领域的前沿。
2)建立保护性农业长效研究体系。SOC的演变是一个较为缓慢的过程, 其短期含量的变化并不能真实反映土壤质量的增减, 因而迫切需要在不同农田类型区建立保护性农业长期定位试验基地, 动态持续监测SOC收支状况、团聚体周转及微生物代谢变化、水分和养分循环特征、病虫害变异规律等与土壤固碳过程密切相关的因素。在考虑气候类型、土壤状况、种植制度等基础上, 明确不同保护性农业措施单独或耦合调控下SOC固定的潜力与实质,从而制定出因地制宜的保护性农业推广方案。
3)关注保护性农业下深层土壤固碳过程。深层土壤是潜在的巨大“碳汇”, 但也存在很多限制微生物活动和根系生长的因素, 如能量来源和养分的缺乏。保护性农业措施如何影响深层土壤微生物的适应性,深根作物根系分泌物如何参与到微生物同化及矿物吸附途径仍有待进一步考证。同时, 探索缓解深层土壤中根系限制因子的有效方法对于增加深层SOC库和作物产量至关重要。
4)构建基于农田实践的SOC管理框架。土壤固碳是恢复退化农田生态系统的必然选择, 但明确特定条件下土壤固碳的阈值水平, 理解SOC与农田生产力及土壤过程、功能的影响不仅仅是一个学术问题, 还需要进一步加强SOC管理实践运动。将土地与农田的管理同碳的管理相结合, 建立起SOC管理框架, 并纳入经济政策、环境可持续方案等要素, 这些体系可以帮助土地管理者和农户制定科学合理的可持续土地利用战略。
猜你喜欢
中国肿瘤外科杂志(2023年5期)2023-11-04 05:50:14
土壤学报(2021年4期)2021-09-10 03:45:40
中国工程咨询(2017年7期)2017-01-31 03:01:26
中国工程咨询(2017年1期)2017-01-31 02:55:32
水土保持研究(2016年4期)2016-10-21 02:28:59
新农业(2016年15期)2016-08-16 03:40:48
西藏大学学报(自然科学版)(2015年1期)2015-12-25 11:43:44
土壤与作物(2015年3期)2015-12-08 00:46:58
现代农业(2015年5期)2015-02-28 18:40:57
作物研究(2014年6期)2014-03-01 03:39:15
