
基于鸟类多样性提升的社区公园生境营造策略探析
2022-04-13 02:58张颖
中国园林 2022年3期
张 颖
黄婷婷
胡 骞
朱建宁*
1 研究背景
1.1 社区公园在生物多样性保护中的作用
社区公园被定义为用地独立,具有基本的游憩和服务设施,主要为一定范围内居民就近开展日常休闲活动服务的绿地[1],规模宜大于1hm2。陶晓丽等[2]的研究表明,其面积之和占北京公园总面积的40%,为众公园类型之首。作为绿地的组成部分,社区公园是城市居民最常接触到自然和使用频率最高的开放空间。
社区公园面积小、分布广、布局灵活,是许多生物迁移的踏脚石(Stepping-stone)和重要庇护场所(Shelter)[3]。在生态网络中,社区公园是外围保护区的重要节点,可提高区域整体的景观连通程度[4],协同保护生物多样性。据Strohbach等的研究,150m2的绿地斑块即可为多一种鸟类提供生存环境[5]。在伦敦,这类小微绿地的设计也要求在提供户外活动场所的同时,通过绿化促进生物多样性和环境绩效,为野生物种提供栖息环境[6]。而在中国,小微绿地的设计却极少考虑生物多样性。
1.2 鸟类在居民日常生活中的积极影响
社区公园生物多样性的提升可以有效促进居民身体健康[6]。作为居民利用率最高的绿地,在其所提供的环境中,自然性因子所产生的恢复性效应最高[7]。植物、动物、景观都属于重要的自然性因子。“鸟鸣声” 是自然声音中对居民恢复效果最显著的一种,吸引感、趣味感等4种感知维度均表现出显著正向恢复效应[8]。鸟类作为城市居民日常最常接触到的野生动物,对人体的身心健康有显著正向影响,有利于疲劳恢复、压力缓解[9]。因此,在社区公园中有意识地提供鸟类等小动物栖息的场所资源,营造更好的生物多样性环境十分有必要[10]。
1.3 公园生境营造的研究现状
基于绿地空间设计对鸟类生境进行探讨的研究目前集中于对某类特定物种进行保护的实践总结。冯沁薇等总结了58个国家湿地公园的营建案例,提出城市湿地公园应针对不同生态类型的鸟类需求进行设计[11]。在对公园整体生境进行探讨的研究中,大型绿地的相关研究较多。如杨刚在对上海4个大型公园的研究中,提出在大型公园设计时,应该通过配置多种景观类型产生丰富的边缘效应[12]。小微绿地的研究较少,干靓等对上海世纪大道沿线街旁绿地研究,提出“提高生物基层质量”和“减缓人工环境干扰”2个维度的提升建议[13]。社区公园的鸟类生境营造研究还尚未有人提及。本文尝试以北京市4个小型社区公园为例,探索鸟类适宜性的社区公园环境营造方法。生境基底与干扰控制通常被看作是影响公园鸟类的两大类因素,本文研究也主要从这2个方面展开。
2 研究方法
2.1 研究区域
北京市公园分布符合规模递减原则,小型社区公园占社区公园的85%[2]8。小型社区公园更具有生态网络中“踏脚石”特点的代表性。参照《城市绿地鸟类栖息地营造及恢复技术规范》[14],以栖息地斑块不得小于2hm2为基准,综合过往对北京市社区公园情况[15]文献的研究,选择南馆公园(2.6hm2)、东单公园(4.3hm2)、双秀公园(6.2hm2)、马甸公园(8.5hm2)作为研究对象。
4个样点公园面积成近似等差数列递增,在对面积效应的研究中互为对照,又方便通过数学运算,弱化面积效应的影响。周边环境方面,4个公园都处于城市核心区域,受到较强城市环境干扰,周边用地情况类似,主要为居住、办公、道路用地,服务对象主要为周边居民。双秀公园与马甸公园位于北三环南北两侧,隔路相对。南馆公园与东单公园则都毗邻城市主要道路,干扰程度类似。公园内部方面,南馆公园、东单公园都集中布置了一个主要铺装场地,放置健身器材。游人活动主要以聚集在场地运动及环绕园路散步为主。其余2个公园为游人提供了更多设施。马甸公园设置了小型篮球场等;双秀公园则设置了小型游乐场。公园景观上,4个公园都由一圈主要园路串联全园。南馆公园设置中央水面,结合湖心岛、拱桥突出水景;东单公园则在北部人工堆叠一座山体,突出山景;双秀公园景观塑造细致丰富,有不同风格主题的园中园。
2.2 调查与分析方法
1)鸟类及植物调查。
调查时间为2018年9月—2019年9月。调研时间在晴朗无风的早晨,调研团队匀速地在公园主路上行进,使用双筒望远镜观察并记录沿途出现的鸟类种类、数量,分布生境特征及栖息模式[16]。在鸟类调查的同时,对植被垂直结构进行记录,并在各公园中设置植物样方,对样方内的植物种类及数量进行统计,样方面积为20m×20m。样方抽样数量按照样点公园面积分配,南馆公园为N1—N2;东单D1—D4;双秀S1—S7;马甸M1—M8。在选择时尽可能多地覆盖不同微生境种类。
2)噪声调查。
使用2台数字声级计对各样点公园各处进行噪声测量,在测量前进行校准,为保证测量的准确性,2台设备同时进行测量,计权方式为A级计权。测量时间为2020年11月中旬,测量时段为游人活动较频繁的周末15:00—18:00[17]。
3)分析方法。
生境基底方面,对各公园生境构成的种类、结构、质量等信息进行详细记录和剖析,笔者采用生境单元制图法,对各公园的生境情况进行空间图示化和数据量化[18],首先通过eCongniton对公园地表覆盖情况进行粗分类,获取公园水平生境结构。分类数据为高分二号(GF-2)高分遥感影像,分辨率为0.95m。后辅以实际调查中获取的植被被垂直结构数据、公园设计图纸等资料,导入ArcGIS map得到4个样点公园的生境单元制图。
最终生境单元制图结果导入Fragstas 4.2中计算并获取每个公园的景观结构数据。这些信息主要包括公园整体生境与各类别微生境类型2个层面。整体层面主要包括:公园的生境总丰富度(PR)、丰富度密度(PRD)、多样性指数(SHDI)等;各类别微生境层面主要包含每一种生境类型的面积(CA)、占比(PLAND)等。这些结构数据及植物样方调查中得到的植物丰富度(RIVv-P)等数据将与通过鸟类调查得到的公园鸟类物种丰富度(RIVb)与鸟类多样性指数(DIVb)进行相关性分析,分析平台为SPSS 23.0。
干扰控制方面,社区公园处于稳定的城市内部环境中,且游人活动类型较单一,无城市边缘区栖息地遭到开发等形式的干扰,也不似大型公园中可能发生诱鸟捕鸟等人类干扰行为。社区公园的干扰源主要为外部城市道路和园内游人活动。这2类干扰的强度都与其产生的噪声具有明显线性关系。谢世林等的研究表明,城市环境通过噪声尤其是低频音,对鸟类的鸣唱、信息传递产生干扰,从而影响其种群密度和群落结构[19]。车流量越大,游人活动强度越大,产生的环境噪声就越强。Reijene等的研究表明,城市道路交通与人类活动所产生的环境噪声越大,鸟类分布密度就越低[20]。因此,本文主要通过对公园中各类风景园林要素对外部交通和内部游人产生的噪声削减和分区控制来进行分析。
分析时利用ArcGIS对收集到的噪声数据进行计算,生成公园声压值等值线,并将其划分为低、中、强3个区域,形成公园噪声强度分布图,叠加鸟类空间分布情况,进行空间耦合。计算方法为样条函数法(Spline)。
3 结果与分析
3.1 各公园微生境组成及结构
各公园微生境构成方式如表1所示,景观结构指数计算结果如表2所示。4个公园中,双秀公园的一到四级微生境丰富度都最高。东单公园是唯一缺少水系的公园,丰富度最低,这一指标与生境多样性的变化规律类似。微生境丰富密度方面,南馆公园最高,说明在极小的空间内做了精细的设计以求植物景观变化丰富,马甸公园则最低。
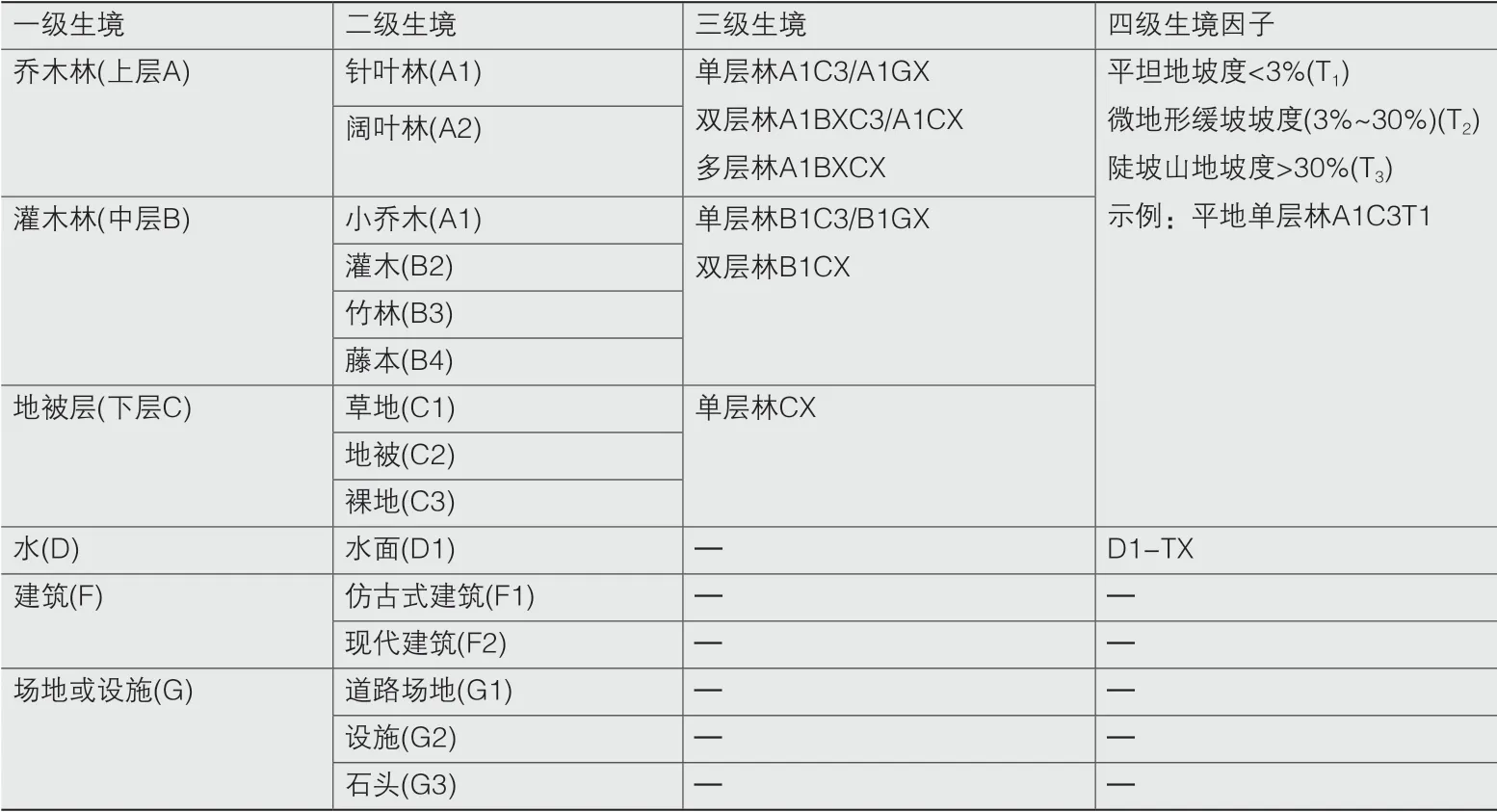
表1 公园各级生境单元组成
3.2 植物调查情况
在对21个植物样方的植物抽样调查中,共统计到植物54种,其中乔木26种,灌木15种,藤本植物3种,地被植物10种。各样方植物丰富度数据如表2所示。
3.3 鸟类空间分布与多样性
本次调查共记录到32种鸟类。各公园鸟类在园内生境空间上的分布情况如图1所示,树麻雀没有在图中进行标注。其中双秀公园鸟类丰富度最高。其次为南馆公园,统计结果如表2所示。由于社区公园鸟类组成较为简单,本研究使用Gleason指数对鸟类多样性进行计算,计算公式为:DIVb=S/lnA[21]。式中,S为样点公园鸟类种数;A为该公园总面积。运算结果如表2所示。
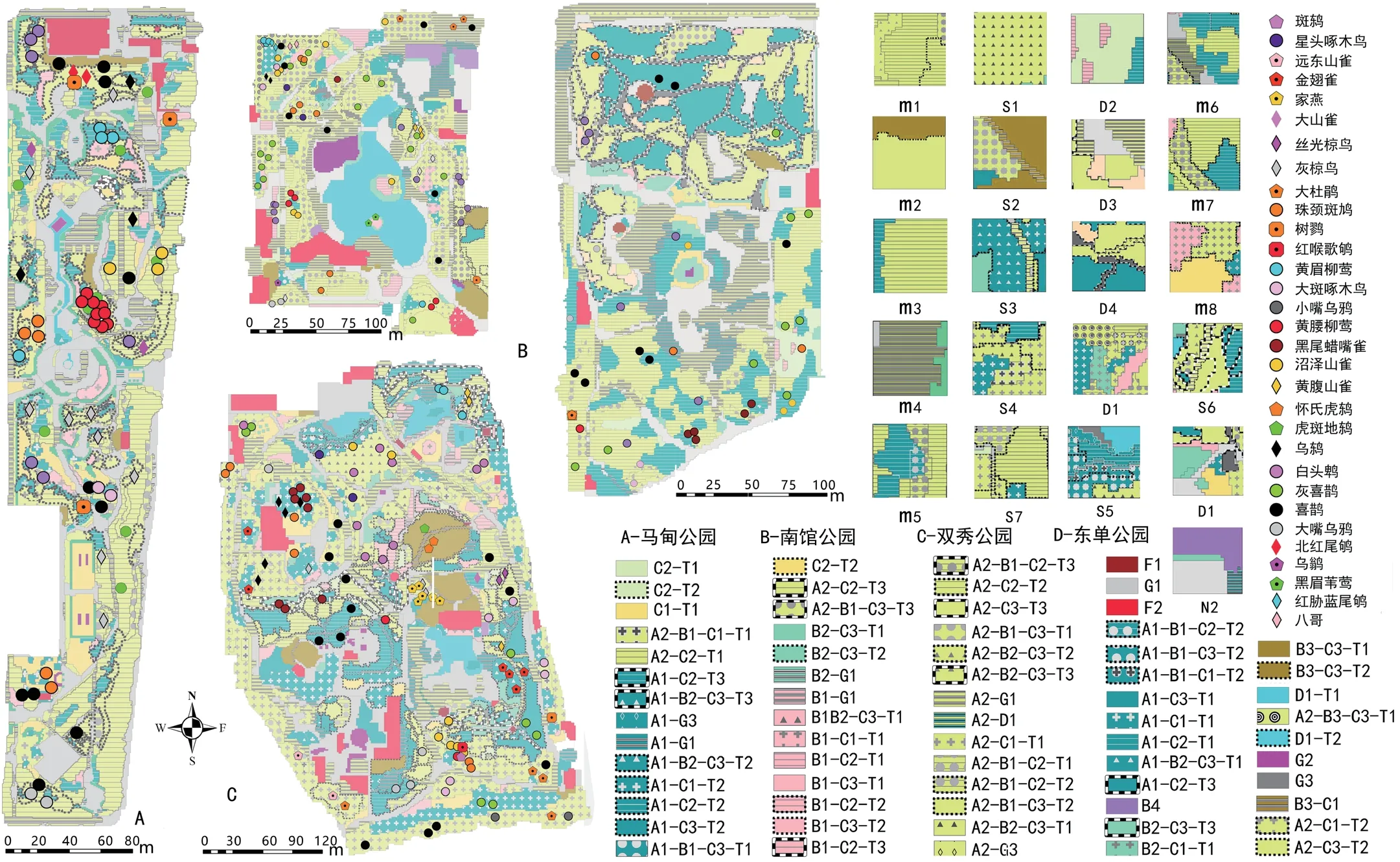
图1 公园生境单元叠加鸟类空间分布制图及植物样方编号图(作者绘)
所记录到的鸟类多为城市常见陆生鸟类。适应城市环境的杂食性鸟类如灰喜鹊等占有较大数量优势。食虫类鸟类如黄腰柳莺、大斑啄木鸟等在种类丰富度上占有优势。被观察到的鸟类多在乔木树冠顶层、远离地面20m以上的位置活动。面积越小的公园中,这一现象更为明显(图2)。在南馆公园中只观察到2只朱颈斑鸠在游人不可进入的草地中觅食,其他鸟类均位于树冠高层。东单公园南侧树木高大处有较多鸟类在树冠高处活动,北侧山体树木低矮,鸟类较少,仅有喜鹊等适应性强的鸟类在离地面12.5m左右的树冠中出现。在面积稍大的双秀和马甸公园中,则观察到沼泽山雀等极少几种鸟类有在灌丛,或游人较少处的地面活动。
3.4 公园生境特点与鸟类多样性的相关性检验
将表2运算结果与鸟类丰富度和多样性进行了相关性分析。分析之前所有数据通过正态性检验。

表2 各公园景观结构及鸟类多样性相关情况
结果显示,PR、SHDI、灌木盖度、植被层次丰富度、样方内的植物种类丰富度5个指标与公园鸟类丰富度显著正相关。PRD、水域面积则与鸟类多样性指数间呈明显的正相关。其余指数则没有与鸟类丰富度和多样性产生明显的相关性。由分析结果可知,在面积有限的社区公园中,微生境越丰富,吸引到的鸟类种类越多。鸟类多样性仅与微生境种类的总数量有关,而与分布是否均匀没有明显关系。灌木覆盖尤其重要,灌木茂密的区域往往植被总层次越丰富,而植物层次越丰富的区域能吸引到更多鸟类。本次实验中的N2、S4样方中鸟类丰富度较高,均为灌木发达的多层植被生境。乔木盖度、公园面积这些指标在本次实验中没有与鸟类丰富度产生明显正相关关系。可能是由于社区公园面积较小,4个公园的面积差不大,并不足以对鸟类多样性产生明显影响。公园总面积、植被总盖度与乔木盖度需要在更大差异梯度范围上对鸟类多样性产生影响。侧面反映了在面积有限的情况下,增加植被层次可能是比增加林地面积更加有效的做法。水面占比仅与多样性数据相关,印证了鸟类对水源有距离要求。水面服务半径越大,越能在单位面积内吸引更多种类的鸟。
3.5 公园空间设计与干扰控制分析
样点公园中鸟类受到的干扰主要为外部道路交通干扰和内部游人游憩干扰。各公园的噪声干扰强度与鸟类空间耦合情况如图2-1所示。通过运算结果可知,南馆公园整体环境噪声干扰较低,而马甸公园则相对受到较高程度的干扰。双秀公园和东单公园内的情况则出现了较大变化,如图2-2的分析结果。从各个样点公园来看,大多数鸟类都选择分布在干扰强度低的区域,双秀公园与南馆公园这一现象突出,马甸公园则各干扰区差值相对较小,说明马甸公园中的鸟类对噪声干扰的敏感度可能相对较低。随着干扰强度升高,鸟类丰富度与多样性逐级递减。
由图2-1中声压等值线变化规律来看,集中的地形、建筑物对噪声削减幅度较大,使得削减距离减小。位于城市快速路两侧的双秀公园与马甸公园在周边交通噪声干扰强度上类似。马甸公园距离边界30m左右的噪声消减情况为5dB左右,双秀公园则为9dB。60m处,消减绝对值为7.5和12.1dB。双秀公园的边界空间对环境噪声的消减效率明显大于马甸公园。东单公园北侧没有明显的边界空间,紧挨护栏的环路环绕山体。以山脊为界线,南侧噪声明显降低。紧邻东侧道路的绿地对噪声的消减情况很微弱。公园南侧的东南方向因有大量外部建筑物阻挡,减噪作用也非常明显。

图2-1 公园噪声干扰强度分级图(作者绘)图2-2 各分级区域鸟类多样性变化趋势(作者绘)
从对外部道路干扰削减的角度看,公园边界的空间设计非常重要。双秀公园边界由相对较高陡的地形将公园内外隔离开来,并在边界上密植排列圆柏,圆柏上层为高大的毛白杨等乔木。边界向内地形起伏丰富,灌木覆盖度高。相较之下马甸公园边界地形平坦,且缺乏中层植物,对干扰的削减能力较差。东单公园的植被层次也较单一,但北侧山体体量大、坡度陡,东南侧场外建筑都对噪声消减起到了很强的作用。
对比4个公园的数据结果可看出,从公园内部布局对干扰的控制来看,双秀公园用地形加郁闭度高的植物将干扰度大的活动场地下沉围合的做法,在削减控制干扰上效果比较明显。在干扰源与植被状况较好的林地间增加建筑作为屏障,也能使得噪声大幅度衰减。另外,马甸和东单公园活动场地布置在公园中心,活动产生的噪声由中心向全园扩散。相比之下,南馆和双秀公园将强干扰场地布置在公园角落的布局对公园整体的干扰影响更小。
4 结论与讨论
4.1 生境营造策略
由上文分析可知,社区公园总面积较小,小梯度范围增加公园面积或通过增加绿地率来扩大少量绿地面积对鸟类多样性的影响并不明显。微生境的多样性和丰富度、植被层次与多样化、灌木的盖度、水系的分布等才是影响鸟类种类的显性因子。社区公园的内部生境营造提升可以总结为以下几条设计策略。
1)增加微生境丰富度。在满足公园基本功能的前提下,在植物、地形、建筑等园林要素上进行丰富的变化和组合,尽量塑造出多样化的生境空间。如地被植物能给一些鸟类提供隐蔽空间,却不利于地栖鸟类在地面取食。不同鸟类因体型、习性不同,对微生境的偏差异较大,多样化的生境才能使更多鸟类栖息。
2)适当增加竖向层次丰富的植物空间。小面积公园中,鸟类受到高强度的人类干扰,植被层次更丰富的生境能给它们起到提供隐蔽场所、屏障削减干扰的作用,且层次丰富的生境本身能同时满足更多鸟类喜好。但应注意的是,过度郁闭的植物和单一化的复杂生境可能会适得其反。本研究中双秀公园的植被竖向层次平均为1.75左右,效果在样本公园中最好。
3)植物类型多样化。多样化的植物能为不同的植食性鸟类提供食源,不同植物的树冠、枝干所形成的空间能给予不同体型的鸟类隐蔽空间。如在双秀公园,虎斑地鸫出现在缓坡竹林底部,而红喉歌鸲则被发现位于黄刺玫灌丛的底部空间。同为对干扰敏感度稍高的鸟类,不同体型的鸟类会选择人类无法进入、空间尺度差异化的植物生境栖息。
4)提高灌木盖度。增加灌木覆盖率,种植多样化的灌木是社区公园鸟类多样性提升的关键。小型公园景观一般比较精致,地被覆盖率高,灌木盖度一定程度上决定了植被层次丰富度。典型的灌木发达生境,如双秀公园中的S4样方中种植了黄刺玫、珍珠梅等5种灌木,除适应程度较高的鸟类外,还吸引了对生境要求稍高的鸟类。灌木底部林下带为小型鸟类最喜爱的活动空间。在灌木种类的选择上可以多选择有一定高度、可形成枝下活动空间、有果实的植物,如黄刺玫、金银木等[22]。
5)提高植被高度。小公园中的鸟类往往在狭小的区域内与人类共处,在水平上拉开距离的空间较小。据观察,除部分城市化适应力强的鸟类如珠颈斑鸠、麻雀外,大多数鸟类都在高大树木的树冠顶层活动,面积越小的公园中,这一现象更为明显,在鸟类行为学研究中也有类似发现[23](图3)。南馆公园虽面积极小,但乔木高度很高,地面人类活动与乔木顶层的鸟类活动相互干扰较小。东单公园北侧山体植被矮小,少有鸟类出现。对干扰敏感度较高的鸟类来说,更高的树高意味着更长的缓冲与警戒距离,中层植被覆盖意味着更高的隔离度。二者为其创造了有效隔离人类干扰的栖息环境。
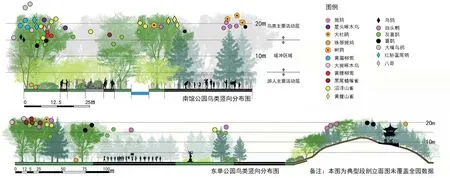
图3 公园鸟类竖向空间分布图(作者绘)

表3 Pearson相关性分析结果
6)水面的设置。绿地能否为鸟类提供水源十分关键,从支持鸟类多样性的角度来看,再小面积的公园也应当设置一定面积的水面。没有条件的公园可以通过微地形形成小洼坑,汇集降雨形成小水洼,为鸟类提供水源。
4.2 干扰控制策略
1)边界设计。在公园与城市主要道路相邻的公园边界,可以通过堆叠较高的地形,搭配郁闭度较高的植物来达到削减干扰的作用。地形和植物都对城市噪声干扰具有削弱作用,是典型的生态声屏障[24]。李冠衡、李梦圆的研究表明,边界植物组合应高低错层多排种植[25],散点式的“品”字形植物排列,对道路的噪声削弱更为明显。种植密度则需根据植物种类的不同进行设计[26]。边界缓冲带越宽、越高,对干扰的削减程度越大,当噪声源为500Hz且与受声点在同一水平面的理想情况下,声屏障高度在3m时,噪声衰减将达到最大值,继续增加高度则减噪效果不明显[27]。在进行边界设计前,可以根据具体道路环境情况,如受声点距离、高度等参数,代入模型进行计算得出相应数值。
2)空间布局。功能布局上,干扰较强的健身场地等,可尽量布置在公园的一角,使其干扰范围减小。同时,可通过地形、郁闭度较高的植物对这些场地进行围和,以提高缓冲效率。
总而言之,社区公园作为家门口的自然,在满足居民的日常需求后,也应重视其生态支撑功能。本次研究揭示了有效的空间生境营造手段能对社区公园的鸟类多样性提升产生积极影响,希望为小微绿地的设计提供参考。本研究的局限性为:因样本量有限,在分析中不能更深度挖掘出各设计策略的主次关系。另外,生境是复杂动态的,影响鸟类对绿地的选择是空间、时间多因子综合作用的结果,而非单一结论可涵盖。本文意在阐明本次实验中出现的显性相关因子,可作为风景园林师提升社区公园鸟类多样性的参考,而非充分必要条件。
猜你喜欢
舰船科学技术(2022年21期)2022-12-12
学与玩(2022年9期)2022-10-31
中国农村水利水电(2021年11期)2021-11-29
风景园林(2021年2期)2021-04-24
落叶果树(2021年6期)2021-02-12
文苑(2020年12期)2020-04-13
小太阳画报(2019年1期)2019-06-11
劳动保护(2019年3期)2019-05-16
体育科技(2016年2期)2016-02-28
客车技术与研究(2014年6期)2014-02-28
