
LincRNA00858靶向miR⁃3126⁃5p影响肝癌细胞的增殖、迁移和侵袭
2022-04-08 06:52雷林翰郑迪杰吴纯宸郑戬谢华华华豪孙诚谊喻超
实用医学杂志 2022年4期
雷林翰 郑迪杰 吴纯宸 郑戬 谢华华 华豪 孙诚谊 喻超
贵州医科大学临床医学院(贵阳 550004);贵州医科大学附属医院肝胆外科,贵州医科大学肝胆胰脾重点实验室,贵州省肝胆胰脾疾病研究所(贵阳 550004)
肝细胞癌(hepatocellular carcinoma,HCC)是侵袭性较强的恶性肿瘤,也是临床上最常见的肝癌类型,预计到2025年全球肝癌发病人数将超过100 万例[1],但HCC 的治疗情况并不令人满意,究其原因主要在于发病机制尚未完全阐明,缺乏针对性的有效手段[2]。因此,深入研究HCC 病变过程的分子生物学机制及发现潜在治疗靶点是攻克HCC 的当务之急。目前研究发现长链非编码RNA(LncRNA)在肝癌、胰腺癌等各种癌症中失调,这表明lncRNA可以作为新的治疗靶点和潜在的生物标志物用于患者的诊断和预后[3-5]。Linc00858 长度为2 685 个核苷酸,在胃癌、结肠癌、卵巢癌、宫颈癌等恶性肿瘤中均为异常高表达。有研究[6-9]指出Linc00858 可以通过调控多种microRNA,从而影响肿瘤细胞的增殖、侵袭、转移和凋亡,另外高表达Linc00858 还与肿瘤的临床分期、淋巴结转移密切相关[10]。因此Linc00858 是在肿瘤靶向治疗中很有前景的候选基因,然而在HCC 中Linc00858 的研究尚少。根据生物信息学分析,本研究选择微小RNA⁃3126⁃5p(miR⁃3126⁃5p)作为Linc00858 的调控对象,通过初步探索Linc00858通过靶向调控miR⁃3126⁃5p 参与HCC 的增殖、迁移和侵袭的分子机制,以期探索新的HCC 治疗靶点。
1 材料与方法
1.1 临床样本及细胞系通过收集2018年9月至2020年12月贵州医科大学附属医院肝胆外科共24 例经病理诊断为肝细胞肝癌的手术切除标本。所有患者在手术前均未接受任何辅助放、化疗。每例标本均取得癌组织和相应的癌旁组织,所有手术标本使用液氮速冻,放置于-80 ℃冰箱保存。本研究获得贵州医科大学附属医院伦理委员会同意,患者均签署知情同意书。人正常肝细胞LO2和人肝癌细胞HUH⁃7,SK⁃HEP⁃1,HEP3B,MHCC⁃97H 及HEPG2 细胞系均购自中国科学院上海细胞生物研究所细胞库。
1.2 实验材料DMEM/High Glucose 培养基、0.25%胰蛋白酶消化液(含EDTA,含酚红)均购自武汉赛维尔生物科技有限公司;胎牛血清(Fetal Bovine Serum)购自以色列biological industries 公司;Trizol 试剂购自美国Invitrogen 公司;逆转录和PCR SYBRGREEN定量试剂盒均购自日本Takara生物技术有限公司,CCK⁃8 试剂盒购自江苏凯基生物技术股份有限公司,Transwell 小室和Matrigel 基质胶均购自美国Corning 公司,双荧光素酶试剂盒购自美国Promega公司;兔抗人波形蛋白(Vimentin)单克隆抗体、小鼠抗人E⁃钙粘蛋白(E⁃cdherin)单克隆抗体、兔抗人细胞周期蛋白E1(cyclinE1)单克隆抗体、兔抗人周期素依赖性激酶2(CDK2)单克隆抗体、兔抗人抗Kip1(p27)单克隆抗体、兔抗人GAPDH 单克隆抗体均购自武汉三鹰生物工程有限公司;RIPA 裂解液、辣根过氧化物酶标记的羊抗小鼠和兔IgG(二抗)、超敏ECL化学发光即用型底物剂盒均购自武汉博士德生物工程有限公司;PVDF 膜购自美国Sigma⁃Aldrich 公司;携带Linc00858cDNA的重组慢病毒及其阴性对照慢病毒、携带Linc00858⁃shRNA 的重组慢病毒及其阴性对照慢病毒、慢病毒感染增强液HitransG A 均购自上海吉凯基因化学技术有限公司;miR⁃3126⁃5p mimics 和miR⁃3126⁃5p inhibitor 及其相应阴性对照均购自广州瑞博生物科技有限公司。Linc00858 上游引物序列5'⁃TG⁃CAATCTGTTTAATGTGCCAGT⁃3',下游引物序列5'⁃CTGGAGCACCTCTCTAAGAACC⁃3';miR⁃3126⁃5p上游引物序列5'⁃ATGCGTGAGGGACAGATGC⁃3',下游引物序列5'⁃GTCGTATCCAGTGCAGGGTC⁃3';GAPDH 上游引物序列5'⁃GGAGCGAGATCCCTC⁃CAAAAT⁃3',下游引物序列5'⁃GGCTGTTGTCATA⁃CTTCTCATGG⁃3';U6 上游引物序列5'⁃CTCGCTTC⁃GGCAGCACA⁃3',下游引物序列5'⁃ACGCTTCAC⁃GAATTTGCGT⁃3'。
1.3 试验方法
1.3.1 实时荧光定量PCR使用Trizol 法提取出总RNA。使用PrimeScriptTMRT reagent Kit(日本Takara 公司)对提取出的总RNA 进行逆转录以获得cDNA;采用SYBR®Premix Ex TaqTM ⅡKit(日本Takara 公司)进行qRT⁃PCR 分析;分别以GAPDH 和U6 作为Linc00858 和miR⁃3126⁃5p 的内参。以2⁃ΔΔCt法值表示目的基因的相对表达水平。
1.3.2 细胞转染和分组将正常肝细胞和各肝癌细胞系从液氮罐中取出,37 ℃水浴锅复苏,用DMEM完全培养基培养(10%FBS、1%青链霉素),放置于恒温37 ℃、体积分数5%CO2的培养箱中进行培养。选取生长状态良好的HUH⁃7 细胞,将其在6 孔板中进行铺板,每孔密度为1×105个/孔,使用慢病毒感染增强液HitransG A 进行重组慢病毒感染,12~16 h 后更换新鲜培养基;分组为Empty Vector组(阴性对照组)和pcDNA⁃Linc00858 组;分别使用mimics⁃NC、miR⁃3126⁃5p mimics 与Linc00858 过表达慢病毒共转染HUH⁃7 细胞,分组为pcDNA⁃Linc00858+miR⁃NC 组与pcDNA⁃Linc00858+miR⁃3126⁃5p mimics 组;MCHH⁃97H 细胞的转染方式如上,分组为sh⁃NC 组(阴性对照)组和sh⁃Linc00858组,分别使用inhibitor⁃NC、miR⁃3126⁃5p inhibitor 与sh⁃Linc00858 共转染MCHH⁃97H 细胞,分组为sh⁃Linc00858+anti⁃NC 组和sh⁃Linc00858+anti⁃miR⁃3126⁃5p 组;以上步骤均严格按照吉凯基因公司《重组慢病毒载体使用手册》和锐博生物公司《miRNA 产品使用说明》进行操作,筛选出稳定表达的细胞株,并采用qRT⁃PCR 法检测转染效率。
1.3.3 CCK⁃8法检测细胞增殖各组细胞按照3×103个/孔的密度接种于96 孔板中,四周每孔加入100 μL PBS 以减少蒸发,随后放入培养箱中培养,待细胞贴壁后观察。分别取6、24、48、72 h 共4 个时间点,每孔加入10 μL 的CCK⁃8 试剂后避光孵育2 h,随后利用酶标仪在450 nm 的波长下检测其吸光值(OD值),并绘制生长曲线。每组设置5 个复孔,且每组实验重复3 次。
1.3.4 Transwell 法检测细胞迁移和侵袭能力将Matrigel 胶与无血清DMEM 培养基按1∶8 的比例配比稀释,在每个小室中加入60 μL 基质,置37 ℃30~60 min(培养箱内)使胶凝固。用200 μL 的无血清培养基将各组细胞密度调整为1×105个/孔,将其加入到侵袭小室内,每种细胞种3 个小室。下室加入含20% FBS 的完全培养基600 μL,培养24~48 h 后。弃去小室内培养基,用PBS漂洗3次。4%的多聚甲醇固定20~30 min,用0.1%结晶紫染色20~30 min,用清水洗3 次,用棉签轻轻拭去小室的上室膜中未穿过小室的细胞,待烘箱将其烘干后,在倒置光学显微镜选取5 个视野拍照记录,将各组细胞进行计数汇总,再进行统计学分析。Transwell 迁移实验中,小室不铺Matrigel 胶,其余步骤与前述的侵袭实验相同。
1.3.5 双荧光素酶报告基因检测构建含有miR⁃3126⁃5p 结合位点的(3'UTR)双荧光报告基因载体(广州市锐博生物科技有限公司),即linc00858野生型(linc00858⁃wt)质粒、linc00858突变型(linc00858⁃mut)质粒。将HUH⁃7 细胞接种于96 孔板中,孵育至细胞密度达70%。然后,将WT·Linc00858、MUT·Linc00858 与miR⁃3126⁃5p mimics 共转染HUH⁃7 细胞中。根据生产厂家的说明书,采用双荧光素酶试剂盒(美国promega 公司)测定各组细胞的荧光素酶活性。
1.3.6 蛋白质印迹法检测HUH⁃7 和MHCC⁃97H细胞中上皮⁃间质转化(epithelial mesenchymal transition,EMT)和细胞周期相关的蛋白表达水平的影响使用胰蛋白酶将各组细胞消化处理后,使用RIPA 细胞裂解液分别提取各组细胞的总蛋白,行SDS⁃PAGE,在90 V 下分离处理后的蛋白样本30 min 后再在110 V 下分离70 min,然后将分离后的蛋白转移到PVDF 膜上,用5% 脱脂牛奶的封闭液封闭2 h,放置于针对Vimentin、E⁃cdherin、cyclin E1、CDK2、p27 和GAPDH 的特异性一抗(体积稀释比例均为1∶1 000)中,于4 ℃下摇床反应过夜;随后加入辣根过氧化物酶标记的羊抗兔/小鼠IgG(体积稀释比例均为1∶5 000)的二抗中,室温下摇床反应2 h,然后将超敏ECL 化学发光液(A 液:B 液为1∶1)滴加在条带上,凝胶成像仪采集图像,最后用ImageJ1.42 软件进行灰度分析。
1.4 统计学方法所有实验至少独立重复3次,统计分析使用SPSS 22.0 软件。正态分布的计量资料以(±s)表示,两组间比较采用LSD⁃t检验。以P<0.05 为差异有统计学意义。
2 结果
2.1 Linc00858 在人肝癌组织和肝癌细胞系中高表达根据TCGA 数据库的数据分析,Linc00858在肝癌组织中的表达情况明显高于癌旁组织(P<0.05),见图1A。qRT⁃PCR 结果显示,相比癌旁正常组织,肝癌组织中Linc00858 的表达水平较高(P<0.05),见图1B;在肝癌细胞系中的Linc00858的相对表达量也显著高于正常肝细胞LO2(P<0.05),见图1C;筛选出相对表达量最高的MHCC⁃97H 和最低的HUH⁃7 作为后续研究对象。
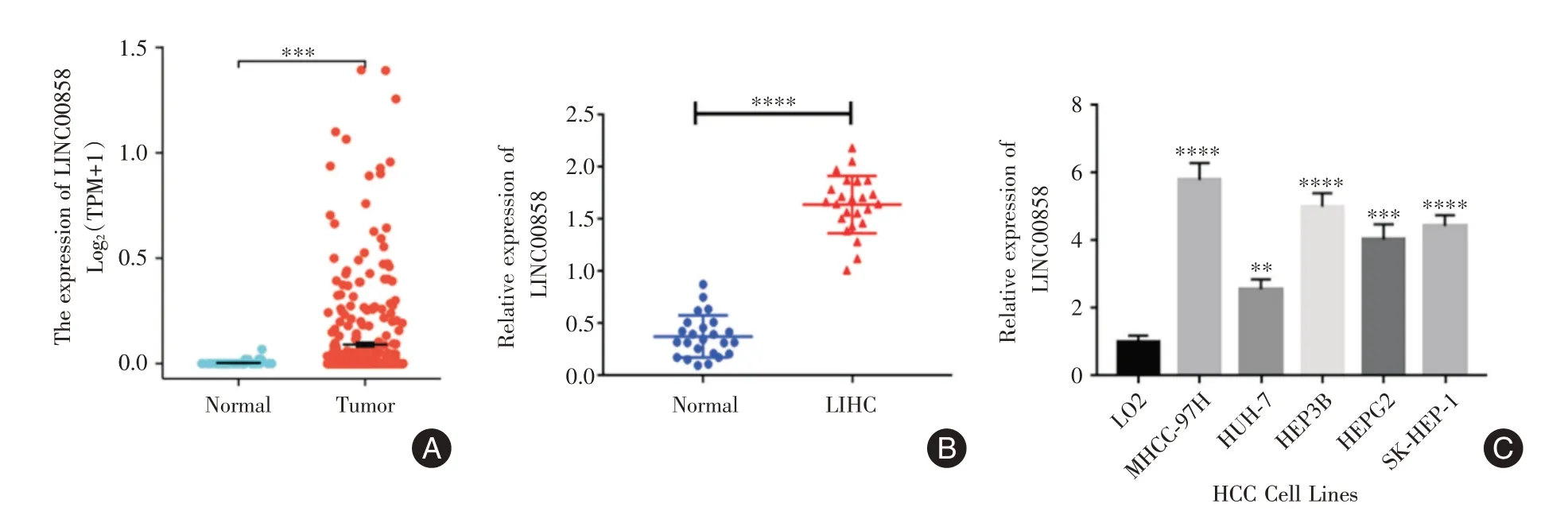
图1 Linc00858 在肝癌组织及细胞中表达情况Fig.1 The expression of Linc00858 in hepatoma tissue and cell
2.2 Linc00858 对肝癌细胞增殖、迁移和侵袭能力的影响qRT⁃PCR检测结果显示:pcDNA⁃Linc00858组的Linc00858 表达水平明显高于Empty Vector组,sh⁃Linc00858 组的Linc00858 表达水平明显低于sh⁃NC组(P<0.05),见图2A。与Empty Vector 组对比,pcDNA⁃Linc00858 组的细胞增殖、迁移和侵袭能力均显著上升(P<0.05),见图2B-D,同时cyclin E1、Vimentin 和CDK2 的蛋白表达水平明显上升,而p27 和E⁃cadherin 的表达水平则明显下降,差异均有统计学意义(P<0.05),见图2E。和sh⁃NC 组比较显示,sh⁃Linc00858 组的MHCC⁃97H细胞增殖、迁移和侵袭能力明显下降(P<0.05),见图2B、C、D;同时cyclin E1、Vimentin 和CDK2 的蛋白表达水平明显下降,而p27 和E⁃cadherin 的表达水平则明显上升,差异均有统计学意义(P<0.05),见图2E。

图2 改变Linc00858 的表达后对肝癌细胞增殖、迁移、侵袭能力和相关蛋白表达情况的影响Fig.2 Effects of altered expression of Linc00858 on proliferation,migration,invasion and expression of related proteins in hepatocellular carcinoma cells
2.3 Linc00858靶向调控miR⁃3126⁃5p的表达通过starbase 网站(https://starbase.sysu.edu.cn/index.php)和LncBASE 网站(https://labworm.com/tool/diana⁃lncbase)发现共有10 个microRNA 可能是其潜在的下游靶基因(图3A)。qRT⁃PCR 检测结果表示,在各组的对比中,miR⁃3126⁃5p 的表达量变化差异最为显著(图3B)。双荧光素酶报告实验结果表明,miR⁃3126⁃5p mimics 能明显降低WT⁃Linc00858 的荧光素酶活性(P<0.05),对MUT⁃Linc00858 则无明显影响(图3C)。qRT⁃PCR 检测结果显示,miR⁃3126⁃5p 在肝癌细胞系中的表达量也显著低于人正常肝细胞LO2(P<0.05,图3D);Spearman 等级相关性实验显示,Linc00858 的表达和miR⁃3126⁃5p 呈负相关(图3E)。以上结果均提示Linc00858 可以负向调控miR⁃3126⁃5p 的表达。
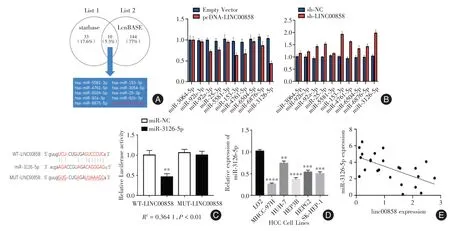
图3 Linc00858 负向调控miR⁃3126⁃5p 的表达Fig.3 Linc00858 negatively regulates the expression of miR⁃3126⁃5p
2.4 改变miR⁃3126⁃5p 的表达水平能够逆转Linc00858 对肝癌细胞增殖、迁移和侵袭能力的影响在HUH⁃7 细胞中,mimics⁃miR⁃3126⁃5p 组的miR⁃3126⁃5p 表达水平明显高于mimics⁃NC 组;而在MHCC⁃97H 细胞中,anti⁃miR⁃3126⁃5p 组的miR⁃3126⁃5p 表达水平明显低于anti⁃NC 组(P<0.05,图4A)。与pcDNA⁃Linc00858+mimics⁃NC 组相比,pcDNA⁃Linc00858+mimics⁃miR⁃3126⁃5p 组在HUH⁃7细胞中增殖、迁移及侵袭能力减弱;蛋白质印迹法检测结果显示对比pcDNA⁃Linc00858+mimics⁃NC 组,pcDNA⁃Linc00858+mimics⁃miR⁃3126⁃5p 组cyclin E1、Vimentin 和CDK2 的蛋白表达水平下降,而p27 和E⁃cadherin 的表达水平则上升(图4E);与sh⁃Linc00858+anti⁃NC 组相比,sh⁃Linc00858+anti⁃miR⁃3126⁃5p 组在MHCC⁃97H 细胞中增殖、迁移及侵袭能力增强,发生了逆转(图4B-D),蛋白质印迹法检测结果显示对比sh⁃Linc00858+anti⁃NC 组,sh⁃Linc00858+anti⁃miR⁃3126⁃5p 组的cyclin E1、Vi⁃mentin 和CDK2 的蛋白表达水平上升,而p27 和E⁃cadherin 的表达水平则下降(图4E)。
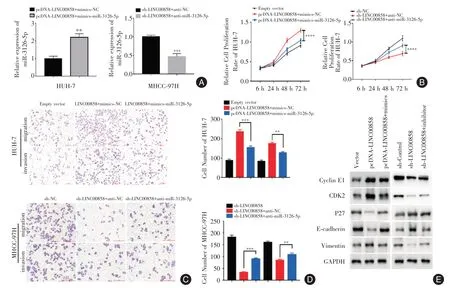
图4 改变miR⁃3126⁃5p 的表达水平后对Linc00858 导致的肝癌细胞增殖、迁移、侵袭能力和相关蛋白表达情况的影响Fig.4 Effects of altered miR⁃3126⁃5p expression on proliferation,migration,invasion and related protein expression of hepatoma cells induced by Linc00858
3 讨论
HCC 是世界范围内最常见的恶性肿瘤之一,是全球癌症相关死亡率的第二大原因[11],其中发生侵袭转移和术后复发是影响患者结局的主要原因之一[12]。目前,手术切除后化疗是最常见的HCC 治疗方案,但患者结局却不尽如人意,患者生存时间未能明显延长[13],因此探索侵袭转移的分子机制对于HCC 的分子诊断、预防和治疗具有重要意义。LncRNA 通常位于细胞的细胞核和细胞质中,尽管不参与编码蛋白质,但许多研究表明LncRNA 在细胞发育、分化和迁移等生物学过程扮演了重要角色[14-16]。刘泽锋等[17]发现,LncRNA DUXAP8 通过增加m6A 甲基化介导的稳定性来维持其表达上调核糖核酸。DUXAP8 水平,这与肝癌细胞增殖、迁移、侵袭和化疗耐药正相关。LncRNA有作为HCC 潜在的生物标志物和分子靶向治疗靶标的可能性。
本研究发现Linc00858 在肝癌组织和细胞中的均呈高表达,并且其能促进肝癌细胞的增殖、迁移和侵袭。在抑制Linc00858 的表达后,其能通过上调p27 的表达,从而抑制CDK2⁃Cyclin E1 的蛋白激酶复合物,而cyclin E1和CDK2是细胞周期G1和S 期转换的限速因子,p27 能抑制cyclin E1 与细胞周期依赖性蛋白结合并诱导细胞周期阻滞从而抑制细胞的增殖;另外在上调了Linc00858表达后,上皮样标志物E⁃cadherin 的表达降低,而间质样标志物Vimentin的表达上升,这提示Linc00858可能是通过EMT 现象增强了肝癌细胞的迁移和侵袭能力。本研究利用生物信息学数据库分析miR⁃3126⁃5p可能是Linc00858 的潜在下游靶点,而目前研究[18]指出,在HCC 患者血清中的miR⁃3126⁃5p 的表达水平比健康人群显著降低,并且其有可能作为HCC早期诊断的重要标记物。双荧光素酶报告基因实验和qRT⁃PCR 结果显示miR⁃3126⁃5p 的表达水平和Linc00858 呈负相关,且Linc00858 可以靶向调控miR⁃3126⁃5p 的表达。通过改变miR⁃3126⁃5p 的表达,本研究发现Linc00858 带来的肝癌增殖、迁移和侵袭能力发生了逆转,此外细胞周期蛋白和EMT 相关蛋白的表达水平也发生了逆转。即Linc00858 可通过发挥ceRNA 机制来竞争性吸附miR⁃3126⁃5p,进而诱导细胞周期蛋白和EMT 相关蛋白的表达。此外,对于Linc00858 和细胞周期与EMT 相关蛋白之间的关系,仍需要进一步研究来阐明其具体调控机制。
综上所述,本研究表明Linc00858 可以通过miR⁃3126⁃5p 调节细胞周期蛋白和EMT 相关蛋白的表达,并促进肝癌细胞的增殖、侵袭和迁移,这表明Linc00858 有望成为肝癌治疗的重要靶点。
猜你喜欢
广东药科大学学报(2022年3期)2023-01-04
生物学通报(2022年1期)2022-11-22
中国农学通报(2022年12期)2022-06-01
皮肤性病诊疗学杂志(2021年5期)2021-11-27
中国种业(2021年11期)2021-11-25
天津医科大学学报(2021年4期)2021-08-21
中日友好医院学报(2021年1期)2021-04-14
广东蚕业(2021年1期)2021-03-18
三农资讯半月报(2021年1期)2021-01-27
山东医药(2020年9期)2020-05-20
