
芦丁对断奶仔鼠生长、肠道抗氧化及脂代谢的影响
2022-04-01 12:09张佳琦熊玮黎智华刘慧娟周斌斌王铭洋王恬王超
南京农业大学学报 2022年2期
张佳琦,熊玮,黎智华,刘慧娟,周斌斌,王铭洋,王恬,王超
(南京农业大学动物科技学院,江苏 南京 210095)
随着畜牧业集约化、规模化的发展,抗生素作为一种促进动物生长,提高机体免疫力的饲料添加剂被广泛应用于饲料工业,由此造成动物性食品中抗生素残留和细菌耐药性问题加重,对人类健康和环境造成严重威胁。中华人民共和国农业农村部发布第194号公告,自2020年7月1日起,饲料生产企业停止生产含有促生长类药物饲料添加剂(中药类除外)的商品饲料。因此,研究和开发健康、无污染的绿色替抗产品迫在眉睫。
1 材料与方法
1.1 试验动物与饲养管理
选取体重相近的21 日龄雄性断奶仔鼠32只(SPF),随机均分为4组,在饲喂的基础日粮中分别添加0(对照组)、250、500和1 000 mg·kg-1芦丁(上海阿拉丁生物科技股份有限公司,纯度95%),于清洁级动物房[温度(22±2)℃,相对湿度55%~60%]中饲养21 d。试验期间仔鼠自由采食与饮水,室内保持12 h光照和12 h黑暗交替。分别在试验1和21 d,称量并记录小鼠体重。饲养试验结束后,将小鼠空腹12 h后称重,再腹腔注射5%水合氯醛(0.01 mL·g-1体重)进行麻醉,接着眼球采血,脱颈致死,取肝脏、肾脏、脾脏和睾丸,称重,然后取空肠迅速放入液氮保存,以备后续分析。血液在3 000 r·min-1条件下离心15 min后,取上清液(血清)于-80 ℃保存。另取肝脏和肠道样品置4%多聚甲醛中固定保存,以备制备组织切片。肝脏、肾脏、脾脏和睾丸的器官指数的计算公式为:器官指数=器官重(g)/小鼠空腹体重(g)×100%。
1.2 测定指标及方法
1.2.1 血清生化以及抗氧化相关指标的测定取0.1 g肠段加入0.9 mL生理盐水,冰水浴条件下匀浆,3 500 r·min-1离心 10 min。取上清液以测定总蛋白含量、总抗氧化能力(total antioxidant capacity,T-AOC)水平、总超氧化物歧化酶(total superoxide dismutase,T-SOD)活性和丙二醛(malondialdehyde,MDA)含量等抗氧化相关指标。取血清,测定T-AOC、T-SOD活性和MDA含量3个抗氧化相关指标,测定总胆固醇(total cholesterol,TC)、甘油三脂(triglyceride,TG)、高密度脂蛋白胆固醇(high-density lipoprotein cholesterol,HDL-C)和低密度脂蛋白胆固醇(low-density lipoprotein cholesterol,LDL-C)浓度等脂代谢相关指标。所有指标均采用南京建成生物工程研究所试剂盒测定。
1.2.2 空肠和肝脏的组织形态学观察将在4%多聚甲醛中固定的组织样(空肠和肝脏),用低浓度到高浓度的乙醇水溶液逐步脱水,再用二甲苯洗掉乙醇,浸蜡包埋。包埋蜡块被切成5 μm厚的切片,用伊红和苏木精进行染色(HE染色),再用带有数码相机(Nikon H550L)的光学双目显微镜(Olympus BX5)观察组织形态。肝脏组织每组选4张切片,每个切片选2~3个视野观察分析。空肠组织每组选4张切片,每张切片选15根完整且排列整齐的绒毛,用Image-Pro Plus 6.0软件测量肠绒毛高度、宽度和隐窝深度。用Excel 2010计算出每张切片中15根绒毛的绒毛高度、宽度、隐窝深度及绒毛高度与隐窝深度比值的平均值。最后,以1张切片数据的平均值为1个重复,在每组4个重复的基础上进行统计学分析。
1.2.3 空肠中基因mRNA相对表达量的测定为了进一步探究芦丁对小鼠肠道抗氧化、脂代谢及肠屏障功能的影响,测定核转录因子E2相关因子2(Nrf2)、血红素氧合酶1(HO-1)、超氧化物歧化酶2(SOD2)、过氧化氢酶(CAT)、紧密连接蛋白3(CLDN3)、黏液蛋白2(MUC2)、过氧化物酶体增殖物激活受体α(PPARα)、过氧化物酶体增殖物激活受体γ(PPARγ)、乙酰辅酶A羧化酶α(ACACA)、硬脂酰辅酶A去饱和酶1(SCD1)和脂肪酸合酶(FASN)基因mRNA的表达量。取约0.1 g的空肠组织样,加1 mL Triol试剂,按照说明书提取RNA,用NanoDrop1000检测RNA的浓度和纯度后,用 15 g·L-1琼脂糖凝胶电泳检验 RNA 的完整性。用反转录试剂盒将统一好浓度的RNA反转录为cDNA,置于-20 ℃冰箱待测。最后,使用SYBR Green qPCR 试剂盒进行荧光定量PCR(qPCR)。反应体系(20 μL):2 μL cDNA模板、0.4 μL上游引物、0.4 μL下游引物、10 μL 2× ChamQ SYBR qPCR Master Mix、0.4 μL 50×ROX Reference Dye和6.8 μL ddH2O。反应程序:95 ℃预变性30 s;95 ℃变性10 s,60 ℃退火30 s,共40次循环。以β-actin为内参,采用 2-ΔΔCT法[16]计算目的基因的相对表达量。试验所使用试剂盒均购自南京诺唯赞生物科技股份有限公司。引物序列见表1。

表1 qPCR引物序列
1.3 数据统计与分析

2 结果与分析
2.1 芦丁对断奶仔鼠体重的影响
由表2可知:与对照组相比,添加250、500和1 000 mg·kg-1芦丁对断奶仔鼠的体重并未产生显著影响(P>0.05)。
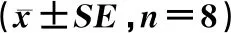
表2 芦丁对断奶仔鼠体重的影响
2.2 芦丁对断奶仔鼠器官指数的影响
由表3可知:与对照组相比,添加250~1 000 mg·kg-1芦丁对肝脏、肾脏、脾脏和睾丸的器官指数均未产生显著影响(P>0.05)。

表3 芦丁对断奶仔鼠器官指数的影响
2.3 芦丁对断奶仔鼠血清抗氧化及脂代谢相关指标的影响
由表4可知:与对照组相比,日粮添加1 000 mg·kg-1芦丁能显著提高血清中HDL-C浓度(P<0.05),而日粮添加250和500 mg·kg-1芦丁能使血清中TG浓度显著降低(P<0.05)。随着芦丁添加量的增加,血清中HDL-C浓度呈线性增加(PL<0.05)。血清TG浓度与芦丁添加量呈二次曲线相关(PQ<0.01)。但日粮添加250、500和1 000 mg·kg-1芦丁对LDL-C和TC浓度未产生显著影响(P>0.05)。
表4 芦丁对断奶仔鼠血清脂代谢相关指标的影响
2.4 芦丁对断奶仔鼠血清和空肠抗氧化指标的影响
由表5可知:日粮添加500 mg·kg-1芦丁能显著提高仔鼠空肠的T-AOC水平和血清中T-SOD活性(P<0.05),添加1 000 mg·kg-1芦丁能显著提高仔鼠空肠中的T-SOD活性(P<0.05)。从线性分析来看,随着芦丁添加量的增加,仔鼠空肠中T-AOC水平和T-SOD活性与血清中T-SOD活性呈线性增加趋势(PL<0.01)。另外,日粮中添加250和500 mg·kg-1芦丁能显著降低仔鼠空肠中MDA含量(P<0.05),且MDA含量与芦丁添加量呈二次曲线相关(PQ<0.05)。与对照组相比, 日粮添加250、 500和1 000 mg·kg-1芦丁对血清中T-AOC水平和MDA含量没有产生显著影响(P>0.05)。
表5 芦丁对断奶仔鼠血清和空肠抗氧化指标的影响
2.5 芦丁对断奶仔鼠肝脏组织形态的影响
由图1可知:对照组肝脏切片中肝细胞形态正常且排列整齐,细胞核呈圆形且居中,未出现核固缩和炎症细胞浸润等现象。日粮添加250、500和1 000 mg·kg-1芦丁对断奶仔鼠肝脏组织形态没有产生明显影响。
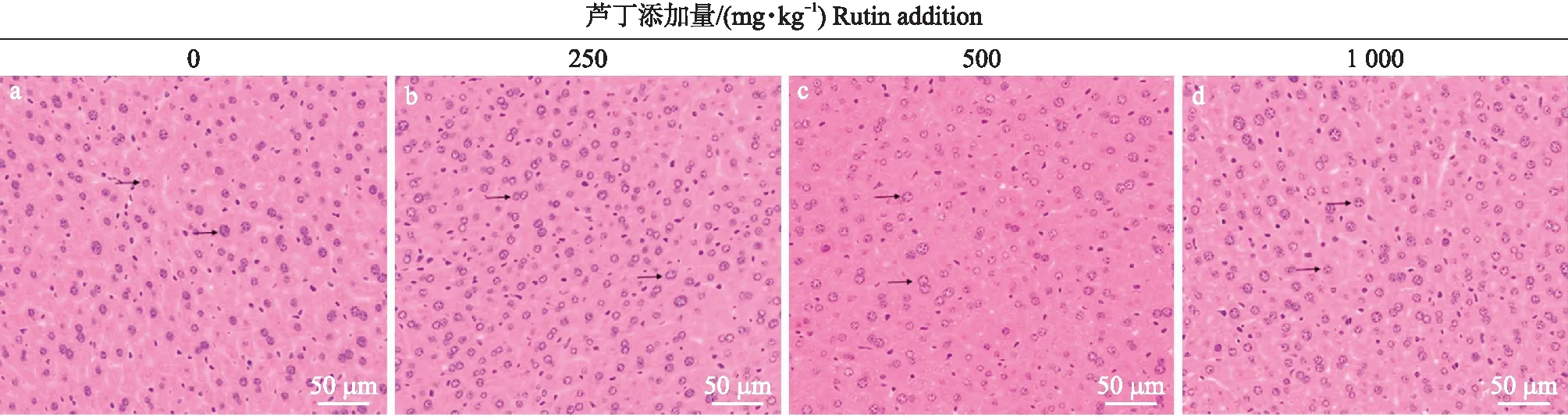
图1 芦丁对断奶仔鼠肝脏组织形态的影响(HE染色)
2.6 芦丁对断奶仔鼠空肠组织形态的影响
由表6可知:与对照组相比,日粮添加250和1 000 mg·kg-1芦丁能显著提高仔鼠的空肠绒毛高度(P<0.05),添加500 mg·kg-1芦丁能显著增加仔鼠绒毛宽度(P<0.05)。从线性分析来看,随着芦丁添加量的增加,仔鼠肠道绒毛高度和宽度以及绒毛高度/隐窝深度比值均呈线性增加趋势(PL<0.05)。但是,日粮添加250、500和1 000 mg·kg-1芦丁对空肠隐窝深度未产生显著影响(P>0.05)。
表6 芦丁对断奶仔鼠空肠组织形态的影响
2.7 芦丁对断奶仔鼠空肠中抗氧化相关基因mRNA表达量的影响
由图2可知:与对照组相比,日粮添加250和500 mg·kg-1芦丁能显著增加仔鼠空肠中HO-1mRNA表达量,添加 500和1 000 mg·kg-1芦丁能显著增加CATmRNA表达量(P<0.05)。250、500和1 000 mg·kg-1芦丁并没有对Nrf2和SOD2mRNA表达量产生显著影响(P>0.05)。

图2 芦丁对断奶仔鼠空肠中抗氧化相关基因mRNA表达量的影响(n=8)
2.8 芦丁对断奶仔鼠空肠中肠屏障相关基因mRNA表达量的影响
由图3可知:日粮添加500 mg·kg-1芦丁能显著增加空肠中CLDN3mRNA表达量(P<0.05)。250、500和1 000 mg·kg-1芦丁对空肠中MUC2mRNA表达量并没有产生显著影响(P>0.05)。

图3 芦丁对断奶仔鼠空肠中肠屏障相关基因mRNA表达量的影响(n=8)
2.9 芦丁对断奶仔鼠空肠中脂代谢相关基因mRNA表达量的影响
由图4可知:日粮添加1 000 mg·kg-1芦丁能显著降低PPARγmRNA表达量(P<0.05),而且250、500和1 000 mg·kg-1芦丁均显著降低FASNmRNA表达量(P<0.05)。日粮添加250、500和1 000 mg·kg-1芦丁对空肠中PPARα、ACACA和SCD1mRNA表达量并没有产生显著影响(P>0.05)。

图4 芦丁对断奶仔鼠空肠中脂代谢相关基因mRNA表达量的影响(n=8)
3 讨论
3.1 芦丁对断奶仔鼠体重和器官指数的影响
芦丁作为一种天然黄酮类化合物,具有改善动物生长性能的功能。Hassan等[17]对肉鸡的研究表明,与对照组相比,饲喂添加1 g·kg-1芦丁的日粮42 d能显著提高肉鸡体重和体增重并降低饲料转化率;在湖羊日粮中添加50和100 mg·kg-1芦丁能够提高湖羊的末重、平均日增重,降低料重比[18]。在本试验中,日粮添加250、500 和1 000 mg·kg-1芦丁对仔鼠断奶后21 d体重未产生显著影响,对肝脏、肾脏、脾脏和睾丸的器官指数也未产生显著影响。这可能与动物种属差异或芦丁的饲喂周期有关,具体原因还有待进一步研究。此外,部分学者对芦丁安全性也进行了研究,Féres等[19]在小鼠急性攻毒试验中发现,在用3 500和5 000 mg·kg-1芸香苷(dimorphandra mollis,芦丁含量约为76%)处理的小鼠体内可观察到部分毒性作用,且出现这种毒副作用的小鼠在24 h后可自我恢复;在对大鼠的慢性毒性评估中,在2 000 mg·kg-1剂量处理下,大鼠血清中丙氨酸氨基转移酶活性在雌性中显著下降,天门冬氨酸氨基转移酶活性在雄性中显著降低但在雌性中显著升高,并显现出明显肺出血,因此推荐芦丁的安全剂量是1 000 mg·kg-1。本试验中饲喂添加250~1 000 mg·kg-1芦丁的日粮21 d未对小鼠肝脏组织形态产生明显影响,提示其对肝脏无明显毒性作用。
3.2 芦丁对断奶仔鼠空肠形态及屏障功能的影响
肠道不仅是机体消化吸收的重要器官,还是重要的免疫器官。多数肠道疾病,例如腹泻、肠炎和食物过敏等均与肠屏障功能障碍密切相关[20]。肠黏膜形态是反映肠道健康与营养物质消化吸收功能的重要指标,绒毛越高、越宽,吸收面积越大,对营养物质的吸收能力也越强[21-22]。本试验结果表明,日粮添加250和1 000 mg·kg-1芦丁均能显著提高仔鼠的空肠绒毛高度,而500 mg·kg-1芦丁能显著增加仔鼠的空肠绒毛宽度,提示日粮添加芦丁可改善断奶仔鼠空肠绒毛形态。研究表明,芦丁因其独特的黄酮多酚结构,主要在小肠被吸收或与小肠内源性蛋白质结合,进而在肠组织或入血转运至机体其他组织发挥作用[23-24]。芦丁可清除肠组织内多余的活性氧(reactive oxygen species,ROS),进而保护肠道免受损伤[11]。这可能是芦丁改善断奶仔鼠肠绒毛形态的主要原因之一。紧密连接蛋白的缺失会破坏肠黏膜的完整性,增加肠道黏膜通透性,影响肠屏障功能发挥[25-26]。CLDN3是一种重要的紧密连接蛋白,也是评价肠道结构完整性的重要指标[27]。本试验结果表明,日粮添加500 mg·kg-1芦丁能显著增加断奶仔鼠空肠中CLDN3mRNA表达量,这与肠道组织形态学结果吻合。以上结果提示日粮添加500 mg·kg-1芦丁对断奶仔鼠肠道黏膜结构及屏障功能有改善作用。
3.3 芦丁对断奶仔鼠抗氧化功能的影响
MDA是机体内评价脂质氧化损伤的重要指标。本试验结果表明,日粮添加250和500 mg·kg-1芦丁能使断奶仔鼠空肠中MDA含量显著降低,提示添加芦丁能有效缓解机体的脂质氧化损伤,这与Liu等[10]在小鼠心脏上的研究结果一致。T-SOD是机体内一种重要抗氧化金属酶,它能有效清除机体内的超氧阴离子,保护细胞免受损伤[28]。HO-1和CAT作为抗氧化酶,可有效维持机体氧化平衡[29-30]。T-AOC代表了待测样品中所有抗氧化剂的总和,一定程度上反映机体清除ROS和一氧化氮合酶(ROS/NOS)的总能力。本试验结果表明,随着日粮中芦丁添加量的增加,血清和空肠中的T-SOD活性呈线性增加的趋势,而且日粮添加500 mg·kg-1芦丁能显著增加空肠的T-AOC水平,与Li等[31]在小鼠肝脏上的研究结果相似,提示日粮添加芦丁可提高小鼠空肠抗氧化功能。芦丁因其结构含有丰富的酚羟基,容易失去H产生较稳定的苯氧自由基进而终止链式反应,同时活泼H与机体内自由基结合而及时清除机体内过多的ROS,这可能是其提高抗氧化功能,保护机体免受氧化损伤的重要原因之一[11,32]。从基因水平来看,日粮添加250和500 mg·kg-1芦丁能显著提高空肠中HO-1mRNA表达量,而且添加500和1 000 mg·kg-1芦丁可显著提高CATmRNA表达量,该结果与血清和空肠组织中的结果相吻合,提示日粮添加芦丁可通过调节抗氧化基因与增强抗氧化酶活性从而提高断奶仔鼠抗氧化功能。Nrf2信号通路是调节HO-1、CAT基因表达及抗氧化酶活性的重要信号通路[33],但本试验中添加250~1 000 mg·kg-1芦丁均未改变空肠黏膜中Nrf2mRNA表达量,因此芦丁调节机体抗氧化功能的机制有待进一步研究。
3.4 芦丁对断奶仔鼠肠道脂代谢功能的影响
脂类的消化吸收主要发生在小肠,在脂酶的作用下大部分脂类水解为甘油一酯和游离脂肪酸后被小肠黏膜摄取,并在小肠黏膜细胞上重新合成TG,最后TG进入血液循环转运[34]。肝脏是机体内重要的代谢器官,是机体合成脂质的重要场所,肝内合成脂质部分以极低密度脂蛋白等为载体通过血液转运到外周(脂肪)组织储存[34-35]。无论是在肠道吸收的脂类,还是在肝和脂肪组织中合成的脂类在血液中多以脂蛋白形式被运送到适当组织去利用、储存或转化[35]。其中,LDL-C的主要功能是向组织转运TC,HDL-C则可以把血液中的TC运至肝脏代谢[35]。因此,血清中TC、TG、HDL-C和LDL-C的浓度可反映机体脂类代谢情况。本试验结果表明,日粮添加1 000 mg·kg-1芦丁显著提高了血清HDL-C浓度,而250和500 mg·kg-1芦丁显著降低血清TG浓度,这与Fernandes等[36]和Prince等[37]的研究结果相似,提示芦丁可能通过抑制肝脏、肠道或脂肪组织中脂质的合成,以及通过调节血清HDL-C浓度促进血浆中的TC向肝脏的转运,进而发挥降低血脂的功能。在本试验中,尽管日粮添加芦丁可降低血清TG及升高HDL-C浓度,但其未对血清TC和LDL-C浓度产生显著影响,这可能与芦丁处理的剂量或时间有关。过氧化物酶体增殖物激活受体(PPARs)是重要的脂代谢传感器和调控因子[38]。PPARγ是PPARs的亚型之一,调控多个基因的表达,编码参与脂肪细胞分化和脂肪合成的蛋白[39]。FASN是一种多功能蛋白,在哺乳动物的新生脂肪合成中起着重要作用[40]。在本试验中,日粮添加1 000 mg·kg-1芦丁能显著降低空肠中PPARγmRNA表达量,250、500和1 000 mg·kg-1芦丁均能显著降低FASNmRNA表达量,该结果与Hsu等[41]和Saleh等[42]的研究结果相一致,提示日粮添加芦丁可能通过抑制肠组织中脂类的重新合成而影响肠TG进入血液,这与本试验中芦丁降低血脂的结果相一致。
综上所述,本试验结果提示饲喂添加250~1 000 mg·kg-1芦丁的日粮21 d未对小鼠产生明显毒性作用,而且适宜剂量芦丁可有效提高断奶仔鼠肠道抗氧化,改善肠道黏膜结构与屏障功能,并可能通过抑制肠道脂质合成和促进血脂转运发挥降血脂功能。芦丁的建议添加量为500 mg·kg-1。
猜你喜欢
新乡医学院学报(2022年6期)2022-11-23
今日农业(2022年2期)2022-11-16
动物医学进展(2022年9期)2022-09-06
现代消化及介入诊疗(2022年4期)2022-07-12
医学前沿(2021年18期)2021-04-14
考试与评价·高二版(2020年1期)2020-09-10
家庭科学·新健康(2018年12期)2018-03-01
分析科学学报(2016年5期)2016-10-15
饮食科学(2016年3期)2016-07-04
药学研究(2015年11期)2015-12-19
