
高山物种栗色鼠尾草(Salvia castanea Diels)访花昆虫多样性与传粉行为变化
2022-03-30 08:58肖汉文黄艳波魏宇昆
生态学报 2022年5期
肖汉文,黄艳波,王 琦,魏宇昆,*
1上海辰山植物园, 上海 201602 2 华东野生濒危资源植物保育中心, 上海 201602
鼠尾草属(SalviaL.)是唇形科(Lamiaceae)世界性分布的最大属,约1000种[1—2],主要分布于北半球的温带、亚热带和南半球的亚热带高海拔地区[3—5]。根据传粉者类型,鼠尾草属可分为蜂媒传粉(蜂媒花,melittophilae)和鸟媒传粉(鸟媒花,ornithophily)。蜂媒传粉物种主要分布于欧亚大陆、地中海沿岸、非洲东部及南非、澳洲以及北美部分地区;鸟媒传粉物种分布于北美西部至墨西哥、中美洲和南美洲安第斯山脉及东部[3,6]。从鼠尾草属的多样性分布中心来看,墨西哥北部至南美洲中部的多样性程度最高,东亚、西亚及地中海区系也是多样性分布中心[7],中南美洲是进化类群的物种分化中心[8]。鼠尾草属的2对雄蕊中的一对退化不育,而另一对雄蕊的花丝与伸长的药隔形成具活动关节的杠杆结构,这种结构与昆虫和蜂鸟的体型及传粉行为高度适应,反映出该属在唇形科中特有且演化程度较高的系统位置[9],杠杆状雄蕊可能是该属适应辐射和多样性形成的关键性状[10]。
作为鼠尾草属的3个主要多样性分布中心之一的东亚[11],该分支可能独立起源于中国西南的横断山至喜马拉雅山脉东段[12—14]。根据花器官结构和花粉接触传粉昆虫的部位,将东亚区系的鼠尾草属物种划分为3种传粉模式类型:短药隔杠杆、长药隔杠杆和退化杠杆[15]。在形态分类上,短药隔杠杆类型属于弧隔鼠尾草亚属(Subg.SalviaBenth.),是中国鼠尾草属下最大的分类群,包含44个种[16]。该类型物种花冠较大,雄蕊药隔伸长较短,一般呈弧形,上下臂花粉囊均发育,传粉时雄蕊上下臂的运动幅度较小,与典型长药隔类型相比(如丹参S.miltiorrhiza),杠杆作用较弱,以背部杠杆传粉模式为主,兼有其他传粉方式,物种的分布区集中在海拔2000 m以上的中国西南高山和喜马拉雅山脉两侧,以横断山区物种多样性最为丰富[15—16]。
1 材料和方法
1.1 研究样地及物种概况
本研究选择云南省玉龙县境内的玉龙雪山(27°09′ N, 100°15′ E)东坡为观测实验样地,样地生境为高山林缘草甸,平均海拔3050 m,面积约400 m2,样地内植株个体数量为50—60株。该地区属于西南季风和印度洋季风气候,干湿季分明,11月至次年5月为旱季,6月至10月为雨季,年平均气温12.8℃,年均降水量935 mm[28—29]。栗色鼠尾草为多年生草本植物,分布海拔在2500—3500 m,生于疏林、林缘或林缘草地,株高为30—65 cm,根系粗大,叶宽大密集多数基生,叶片较厚且多毛。
1.2 花器官形态特征与开花物候
我们对单株花序总数、花序侧生分枝数、单一分枝轮伞花序数、轮伞花序小花总数、轮伞花序同时开放小花数、单株总花数和单株每天总开花数进行统计记录。用电子游标卡尺(精确度:0.01 mm)对花各部结构进行测量,测量的各部分如图1所示。包括花冠长(corolla length, col)、花冠宽(corolla width, cow)、花冠高(corolla height, coh)、冠筒长(tube length, tul)、冠口高(entrance height, enh)、冠口宽(entrance width, enw)、花丝长(filament lenght, fil)、药隔长(connective length, conl)、药隔上臂长(length of upper connective arm, luca)、药隔下臂长(length of lower connective arm, llca)、雌蕊长(pistil length, pil)等。在栗色鼠尾草盛花期的8—9月,详细记录单花花期、单株花序花期和群体花期。

图1 栗色鼠尾草花器官测量示意图
1.3 访花昆虫的行为和传粉特征
在2014—2016年的盛花期,选择晴好天气条件(2014年9月10—12日;2015年9月4、5和8日;2016年9月13、15、17和18日),并在2014年选择不同日期的全天阴雨条件(2014年9月8—9日),用SONY数码摄录机(HDR-CX510E)连续观察录制白天(8:00—18:00)样地内固定标记的栗色鼠尾草单株花序所有访花昆虫及其访花行为,鉴定昆虫种类,统计访花昆虫的数量和访花次数,对访花昆虫进行形态指标测量,包括体长(body length)、胸宽(thorax width)、胸厚(thorax thickness)和喙长(tongue length)。
参考Talavera等[30]以及龚燕兵和黄双全[31]的方法,确定访花昆虫行为,统计计算传粉昆虫的访花时间及频率等传粉特征指标,具体内容如下:1)访花行为(visiting behavior):分为3种类型,包括有效传粉(effective pollination):访花昆虫在进入鼠尾草冠筒吸取花蜜的同时,将其携带的花粉带入或将正在访问花朵的花粉携带出去,并完成有效授粉的过程;盗蜜(nectar robbery):这类昆虫访花时并不进入花冠筒,而是通过在花萼上打洞取食花蜜,也不会触碰柱头及传递花粉,是一种非有效的传粉行为;取食花粉(feeding pollen):访花时,这类昆虫仅落至在花冠口取食花粉,并不触碰柱头,无法完成授粉过程,也是一种非有效的传粉行为。2)访花相对频次(relative frequency,F):指某一种访花昆虫在记录视频中的出现次数占所有访花昆虫出现总次数的相对比例。3)昆虫单花访花时间(visiting time per flower,T):即访花者从登陆花冠口到访花结束后飞离花冠的时间。4)活跃度(activity rate,AR):以单只昆虫每分钟的访问花朵数量表示,代表了此种昆虫对栗色鼠尾草的偏好程度和传粉潜力。5)访花频率(visitation rate):用指数IVR=F×AR表示,访花频率同时考虑了某种昆虫的活跃度及其出现的相对频次。6)昆虫访花日动态,特指某一种访花昆虫,在单位时间段的访花总次数占当天总开放花朵数的比例日变化曲线。
1.4 栗色鼠尾草的繁育系统特征
通过以下不同处理方式,可以了解栗色鼠尾草的繁育系统特征:1)开放授粉(open pollination),检测自然状态下的结实率;2)直接套袋(bagged),检测其主动自交结实情况;3)同株异花授粉(geitonogamy),去雄后人工同株异花授粉,检测自交亲和性的结实情况;4)异株异花授粉(xenogamy),去雄后人工异株异花授粉,检测异交亲和性的结实情况;5)去雄不套袋(emasculated and unbagged),剪除未成熟雄蕊,检测自然条件下昆虫传粉的结实情况。上述每种处理随机选择15株上共计101—158朵花,待果实完全成熟后统计结实率。
另外,为了划分繁育系统类型,还需计算花粉与胚珠比(P/O),即每朵花的平均花粉数量除以胚珠数量。以及异交指数(outcrossing index, OCI),参考Dafni[32]的方法进行异交指数的计算1)花朵直径<1 mm时记为0;1—2 mm时记为1;2—6 mm时记为2;花朵直径>6 mm时记为3。2)花药开裂时间与柱头可授时期的间隔,雌雄蕊同时成熟或雌蕊先熟记为0;雄蕊先熟记为1。3)花药与柱头的空间相对位置,相同高度记为0;空间分离记为1。三者之和即为OCI值。评判标准为:OCI=0时,繁育系统为闭花受精型;OCI=1时,繁育系统为专性自交型;OCI=2时,繁育系统为兼性自交型;OCI=3时,繁育系统为自交亲和型,有时需要传粉者;OCI≥4时,繁育系统为异交型,部分自交亲和,需要传粉者。
1.5 数据分析
所有数据用SPSS 22.0(Chicago, IL, USA)进行分析,2014和2016年花各部形态指标的比较采用t检验,连续3年花序数量和开花特征,2014年和2015年不同处理方式的结实率用单因素方差分析,并用Tukey分析进行事后两两比较,检验差异源。
酒店服务业市场的持续开拓发展,酒店管理专业迈向“专业化、国际化、企业化”的趋势都在激励着酒管专业试行新的人才培养模式以顺应市场。基于双证融通双相融合下的现代学徒制在试点过程中取得了不错的成果,为酒店业乃至社会培养了一批精英人才,同时也为培养体系探明了一条可行之路。还需要不断总结经验,分析当前现代学徒制面临的困难,不断探索完善现代学徒制,为高职院校人才的培育提供支持。
2 结果
2.1 栗色鼠尾草的开花特征、花形态与传粉方式
每年,栗色鼠尾草的始花期在07月25日左右,盛花期为9月5日—21日,至10月10日左右完全凋谢,果期大约为8月10日—10月20日。2014年我们记录的单花花期为(74.58±6.13)h(n=14),单株花序花期为(13.67±0.27)d(n=3),群体花期约78 d。决定鼠尾草开花特征和影响传粉行为比较重要的指标,如3年平均的单株花序总数为(4.49±0.21)个,花序侧生分枝数是(5.94±0.74)个,单株总花数为(563.61±43.86)朵,单株每天总开花数为(65.71±12.16)朵。其中3年平均花序侧生分枝数(平均为5.94,F=11.652,df=2/135,P<0.001)和单株每天总开花数(平均为65.71,F=15.508,df=2/259,P<0.001)有极显著差异,主要表现在2015年均值显著低于2014和2016年,3年的花序各层次的平均数量及开花特征见表1。
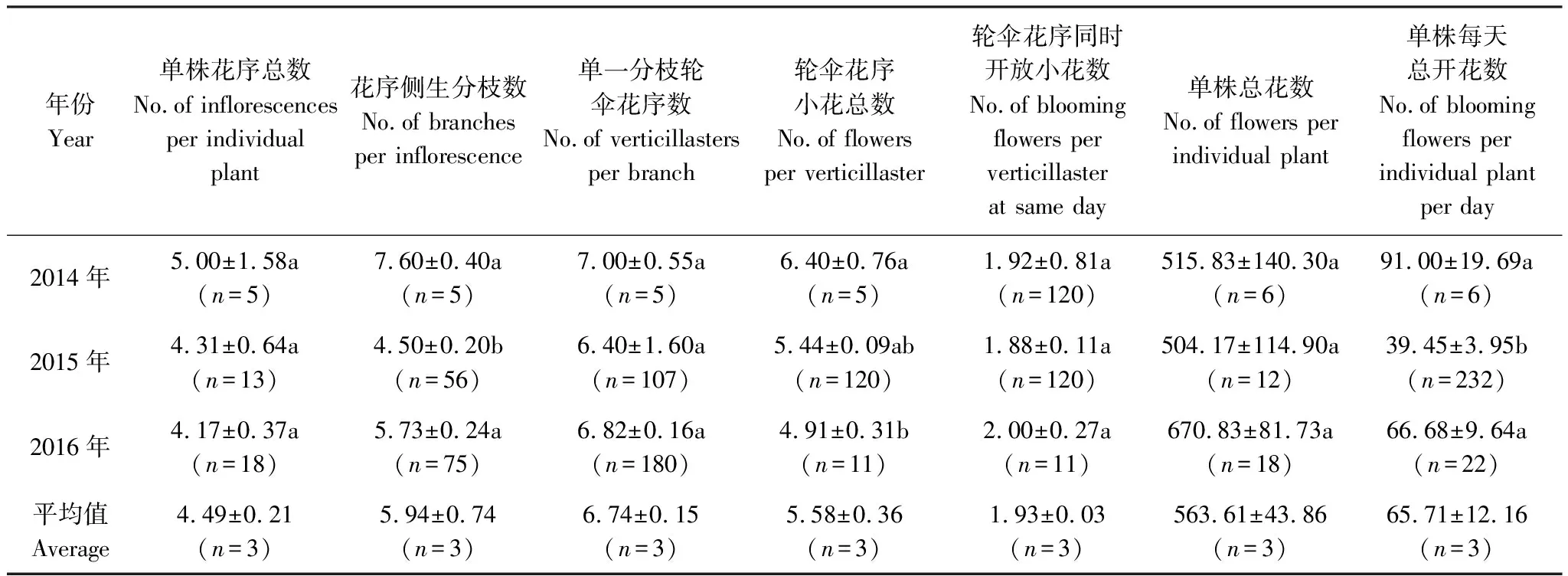
表1 栗色鼠尾草花序数量特征及年际差异比较
栗色鼠尾草的花呈现两侧对称的筒状结构,花冠口较宽,冠筒中部至基部明显变窄且呈“S”形弯曲,上唇略上翘,下唇外翻垂直下伸。4枚雄蕊2枚完全退化不育,另外2枚可育雄蕊演化为特殊的杠杆结构,即每个花粉囊中的药隔显著伸长将2个药室撑开,而花丝与药隔的连接处形成具有类似“关节”(joint,j)的组织,以使得药隔可以关节为轴,进行上下运动,关节以上称为上臂(upper connective arm,uca),以下称作下臂(lower connective arm,lca),上臂具有可育的花粉囊,组成上臂花药(upper arm anther, uaa),成熟后纵向开裂散粉,隐藏在上唇内部,下臂具有较小的花粉囊,组成下臂花药(lower arm anther, laa),下臂花药内侧靠合,只产生很少的花粉,成熟后横向开裂散粉(图1;图2D—F)。2个花丝生于冠筒底部,靠近冠筒口两侧,略向冠筒中间倾斜。柱头(stigma,s)稍伸出花冠,成熟后显著二裂(图2G)。花冠筒基部具有一个花盘(disc,d),可分泌花蜜(图2G)。柱头完成授粉后花冠脱落,花萼宿存包裹4枚小坚果(nutlet)。
栗色鼠尾草群体花期长,单株花朵数多,花序由轮伞花序组成假圆锥花序,单花具有熊蜂偏好型(typical bumblebee flower)的筒状结构。栗色鼠尾草花形态结构属于典型的短药隔传粉模式,杠杆作用较弱,以背部杠杆传粉模式为主,花粉常涂抹在传粉者头部或背部(图2H),借助腹部传粉的情况较少。

图2 栗色鼠尾草的花形态与传粉机制
2.2 栗色鼠尾草的访花昆虫及其行为
2.2.1访花昆虫及其行为多样性
我们对栗色鼠尾草连续3年的观察,共记录到9种访花昆虫(图3)。按照访花行为和传粉有效性可分为3类:第一类是有效传粉昆虫,包括桔尾熊蜂Bombusfriseanus、灰熊蜂B.grahami、圣熊蜂B.religiosus和中华蜜蜂Apiscerana。3种熊蜂的传粉行为相似,传粉时昆虫落置在下唇的着陆点,随后进入花冠筒取食花蜜的同时,推动位于花冠口中间的下臂,通过杠杆作用,将上臂花粉涂抹在传粉者背部,并携带出花冠,将其传递到相邻小花已成熟向下弯曲的雌蕊柱头,完成传粉过程。中华蜜蜂访花时,可正面推动杠杆运动,下臂花药可以将花粉传到蜜蜂头部,当正面无法推动杠杆运动时,蜜蜂可倒挂着进入花冠,带动杠杆上臂向下翻转将花粉落置在腹部(图3A—D);第二类是盗蜜昆虫,包括灰熊蜂B.grahami、桔尾熊蜂B.friseanus和小雅熊蜂B.lepidus,它们访花时并不进入冠筒,而是通过在花萼上打洞取食花蜜,也不会触碰柱头及传递花粉(图3E—G);第三类昆虫取食花粉,包括宽跗蚜蝇属Platycheirus.sp、鼻颜蚜蝇属Rhingia.sp、黑带食蚜蝇Episyrphusbalteatus和隧蜂科彩带蜂属Nomia.sp的物种,这些昆虫仅落至在花冠口取食花粉而并不触碰柱头,无法完成授粉过程,是一种非有效的传粉行为(图3H—K)。研究结果表明:圣熊蜂、小雅熊蜂、中华蜜蜂、宽跗蚜蝇属、鼻颜蚜蝇属、黑带食蚜蝇和彩带蜂属具有单一的访花行为,桔尾熊蜂和灰熊蜂则表现2种不同的访花现象,既能够有效传粉,也存在盗蜜行为。

图3 栗色鼠尾草的访花昆虫及其访花行为
2.2.2昆虫访花行为的年际变化
在良好的天气条件下,栗色鼠尾草传粉昆虫的年际(2014—2016年)变化很明显,2014年以有效传粉行为占绝对优势,至2015年发生逆转,有效传粉者的种类和传粉频率均急剧下降,并以盗蜜占绝对优势,尽管2016年的有效传粉频率有所恢复,但仍低于2014年。此外,连续3年访花昆虫的组成上也发生了明显变化,如具有相同行为的昆虫种类发生变化,以及同一种昆虫在年际间的访花行为发生了根本性的转变——从有效传粉改变为盗蜜。
2014年的传粉昆虫组成为桔尾熊蜂、灰熊蜂、圣熊蜂和中华蜜蜂,其中桔尾熊蜂的相对访花频次为0.65±0.04(n=3),访花频率最高为2.59,是最主要的有效传粉者。2种盗蜜昆虫尽管具有相对较高的活跃度(桔尾熊蜂7.47,灰熊蜂8.37),但访花频率较低(桔尾熊蜂0.07,灰熊蜂0.03),而取食花粉昆虫的活跃度(宽跗蚜蝇属0.88、鼻颜蚜蝇属1.46、彩带蜂属1.68)和访花频率均极低(均低于0.08,表2)。然而,2015年的传粉昆虫种类和有效传粉次数均显著减少,仅有桔尾熊蜂1种传粉昆虫,传粉频次(0.06±0.02)和频率(0.15)也降至很低的水平,大多数的传粉行为转变为盗蜜,桔尾熊蜂由2014年的主要传粉者转变为主要盗蜜者,盗蜜频次(0.74±0.10)和频率(7.14)均较高。至2016年则完全未发现桔尾熊蜂的活动,而是由灰熊蜂和圣熊蜂进行传粉,尽管两种传粉者的访花频次较低,但圣熊蜂单花访花时间较短(3.10±0.23 s/朵),具有相对较高的活跃度(19.35)和访花频率(3.68),盗蜜者则转变为灰熊蜂和小雅熊蜂,同时灰熊蜂有相对较高的盗蜜频次(0.71±0.21)和频率(16.58),而取食花粉的2种昆虫也具有较长的单花访问时间以及较低的访花频次和频率(表2)。
2.2.3昆虫访花行为与天气的相关性
本研究对2014年的晴天与阴雨天气的昆虫访花行为进行了对比观测,在两种天气条件下,尽管访花昆虫的组成没有改变,但不同访花昆虫出现的相对频次变化较显著(表2)。晴天时,桔尾熊蜂占所有访花昆虫的相对频次为0.65±0.04,有效传粉频率为2.59,全天超过一半的访花均为桔尾熊蜂的有效传粉。阴雨天气时,尽管访花昆虫的组成变化不大,但有效传粉的数量明显下降,转变为灰熊蜂的盗蜜为主(相对频次0.29±0.14,访花频率2.43),桔尾熊蜂的相对频次和有效传粉频率则分别降为0.20±0.07和0.80。同样的,圣熊蜂的访花频率也明显受到了天气状况的影响,而中华蜜蜂的影响却小很多。取食花粉在总的访花行为中占比较小,尤其在晴天条件下,但阴雨天宽跗蚜蝇属昆虫取食花粉频率有比较高的水平(表2)。

表2 连续3年栗色鼠尾草访花昆虫的访花特征及行为变化
桔尾熊蜂和圣熊蜂在两种天气条件下有相似的日动态规律,晴天主要都是传粉行为,而阴雨天的传粉频率明显降低,但总体趋势均为单峰型,即中午的访花频率最高,早晚频率降低(图4)。中华蜜蜂的日动态则略有不同,晴天主要在8:00—13:00进行传粉,阴雨天传粉频率没有明显规律,但基本出现在11:00以后。从图4中可以看出,晴天总有效传粉为单峰曲线,最大传粉频率出现在中午13:00—14:00,随后快速下降;阴雨天总有效传粉频率日变化也为单峰型,峰值稍有提前,且总体频率值显著降低。盗蜜的情况与有效传粉正好相反,阴雨天的访花频率显著高于晴天,且日变化呈现双峰曲线,早晚较高而中午较低。两种天气条件下取食花粉的日动态变化不显著,只有阴雨天早晚较高,而其他时间段天气对此行为的影响不大,均呈现较低的水平。

图4 2014年晴天和阴雨天的昆虫访花日动态(晴天,n=3; 阴雨天,n=2)
2.3 栗色鼠尾草的繁育系统
栗色鼠尾草每朵花的花粉数量为12915.03±1964.70(n=10),每朵花具4个胚珠,花粉胚珠比(P/O)为3228±491;栗色鼠尾草的花朵直径>6 mm,雌蕊先熟,雌雄异位,因此花朵异交指数(OCI)=5,根据P/O和异交指数,栗色鼠尾草繁育系统为专性异交,需要传粉者。
栗色鼠尾草2年的不同处理间结实率整体上存在显著差异(图5,2014,F=147.328,df=4/647,P<0.001;2015,F=216.591,df=4/711,P<0.001)。连续2年栗色鼠尾草都具有较高的自然结实率(2014,(88.58±1.54)%; 2015,(71.54±1.56)%,P<0.001)和较低的自交结实率(2014,(34.83±1.89)%; 2015,(15.48±1.94)%,P<0.001),且2015年均显著低于2014年(图5)。2年套袋的自交结实率显著低于其他处理的结实率(图5,2014,P<0.001; 2015,P<0.001),说明栗色鼠尾草主要依靠传粉者授粉,相同年份的开放授粉和异株异花授粉的结实率(图5,2014,P>0.940;2015,P>0.835)没有显著差异,说明栗色鼠尾草的传粉不受花粉限制,自然去雄和开放授粉的结实率(图5,2014,P>0.271;2015,P>0.308)也没有显著差异,表明传粉也不受传粉者的限制,同株异花授粉和异株异花授粉的结实率(图5,2014,P>0.858;2015,P>0.979)之间无显著差异,说明栗色鼠尾草没有近交衰退。

图5 连续2年栗色鼠尾草不同授粉方式的结实率比较
3 讨论
栗色鼠尾草单株花序总数(4.49±0.21)和花序侧生分枝数(5.94±0.74)较多,单株花量大(563.61±43.86)朵,说明其植株生长势强,分枝较旺盛,花序较密集,有利于增加整株植物群体花朵的展示度,从而吸引更多的传粉者。相应地,尤其是在自交亲和的物种中,传粉者可连续访问同一株上的几朵花,也增加了同株异花的授粉机会,从而增加花粉的散播和结实率[27]。花序侧生分枝数和植株总花数是植物生殖生长状况的一种反映,单株每天总开花数无疑会对传粉昆虫的吸引能力和访花频率产生影响[33]。例如,2015年鼠尾草花序侧生分枝数和单株每天总开花朵数显著低于2014年,而2015年传粉昆虫种类的急剧减少,访花行为的变化以及显著降低的结实率可能均与此相关。
从本研究结果看,西南高山分布的栗色鼠尾草访花昆虫多样性高,共有9种访花昆虫,分别属于熊蜂属Bombus、蜜蜂属Apis、宽跗食蚜蝇属Platycheirus、鼻颜蚜蝇属Rhingia和彩带蜂属Nomia。而华东区系同域分布的南丹参S.bowleyana和舌瓣鼠尾草S.liguliloba仅有熊蜂属1种传粉昆虫——三条熊蜂B.trifasciatus,当其访问这两种鼠尾草时,可通过机械隔离(mechanical isolation)使花粉落置在传粉者身体的不同部位,形成合子前隔离从而维持物种稳定,暗示它们具有较高程度的共生进化关系[34]。另外的研究还表明,我国北方分布的荫生鼠尾草S.umbratica有4种有效传粉熊蜂,在缺少传粉者时,可发生延迟主动自交为其繁殖提供保障,而如果一旦有昆虫访问,其单次授粉即可充足受精结实,花朵随即掉落从而减少资源消耗,是一种更加灵活的繁殖策略[35]。相比之下,在高山地区气候多变的情况下,栗色鼠尾草的访花昆虫多样性更高,较大冠筒的基部可能存贮更多的花蜜,产生更多的花粉吸引访花昆虫(未发表数据),从而强化传粉服务,提高植物的繁殖成功和生态系统功能[36—38]。
盗蜜是指鸟类、昆虫或其他访花者通过在花上打洞而取食花蜜的现象[39],事实上,几乎在所有具管状花或有距的植物中都会发生盗蜜现象[40]。通常当地的气候条件、动植物形态的不匹配、昆虫的资源竞争和领地争斗以及觅食行为等都可能引起盗蜜的发生[41—47],本研究发现,除了天气条件可能引起栗色鼠尾草的盗蜜现象外,传粉者体型与花器官的匹配程度和传粉行为密切相关。2014年桔尾熊蜂的胸宽((6.14±0.13)mm)和胸厚((5.22±0.07)mm)与冠口宽((6.74±0.15)mm)和冠口高((5.20±0.09)mm)完全匹配,而灰熊蜂的胸宽((7.57±0.06)mm)和胸厚((6.06±0.44)mm)则大于冠口宽和高,体长+喙长((16.86±1.37)mm)显著小于冠筒的长度((29.1±0.41)mm)(表3,表4),我们推测昆虫与花形态结构的适应性是桔尾熊蜂主要进行传粉而灰熊蜂主要进行盗蜜的原因之一。另一方面,栗色鼠尾草盗蜜者的访花时间显著短于有效传粉的时间,单花访花时间越短其相应的频率越高,盗蜜者就能获取更多的花蜜资源。同时,盗蜜减少了花蜜资源,传粉者不得不飞向更远的地方获取花蜜,可将花粉传递到更远的距离[48—50],可能促进了栗色鼠尾草的异花授粉。2015年传粉者种类的急剧减少可能与昆虫对环境的不同响应机制、熊蜂的活动规律、行为特征、不同天气条件下花朵吸引传粉昆虫的细微变化相关[41—43],传粉者行为的转变,可能降低了栗色鼠尾草的自然结实率。除盗蜜之外,栗色鼠尾草还有取食花粉的昆虫,取食花粉行为可能减少了花粉的数量,而花粉数量和质量的下降可能引起传粉限制[51]。有效传粉是保证植物繁殖成功的关键,尽管不同年份不同天气条件下栗色鼠尾草均存在盗蜜和取食花粉的行为,但鼠尾草属植物可能只需要很少的有效传粉者拜访,即可使单朵花的所有胚珠完成受精并结实[50]。另外,栗色鼠尾草在一定强度盗蜜和取食花粉情况下,仍然保证了较高的自然结实率,是否与较高的花粉质量和传粉效率有关,以及非有效传粉在多大程度影响结实,仍需进一步深入研究。
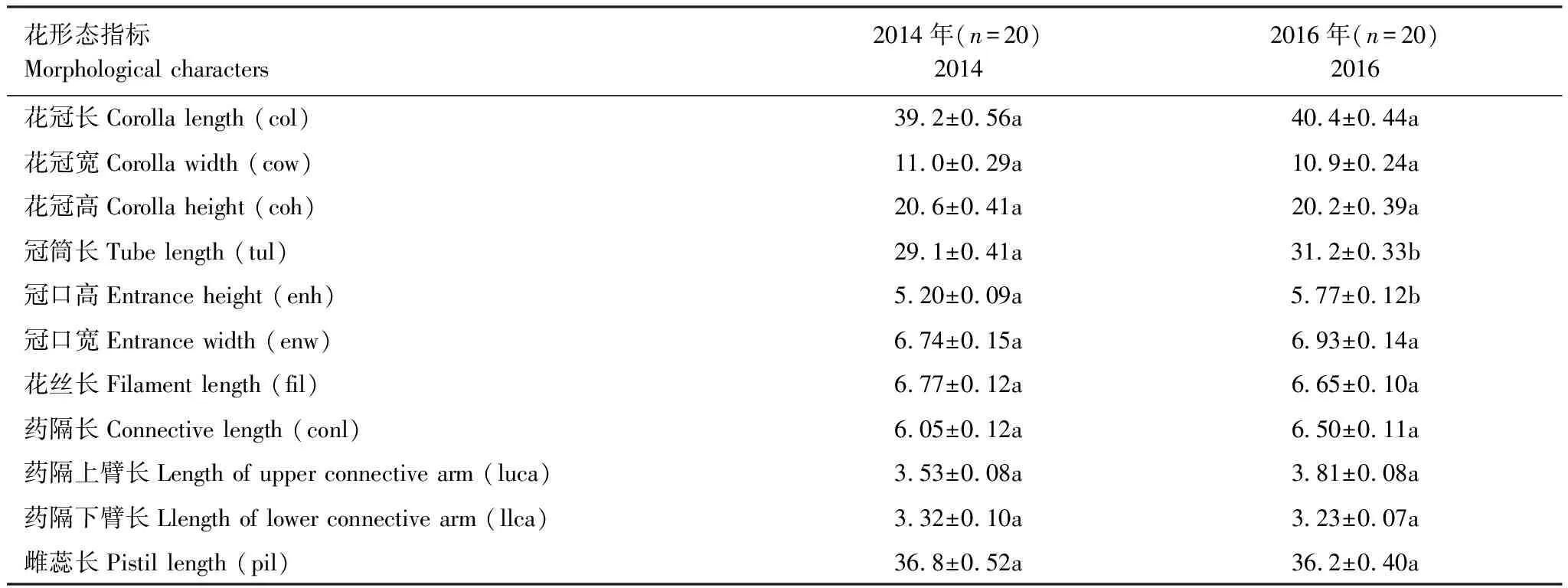
表3 栗色鼠尾草花形态指标及年际差异比较/mm
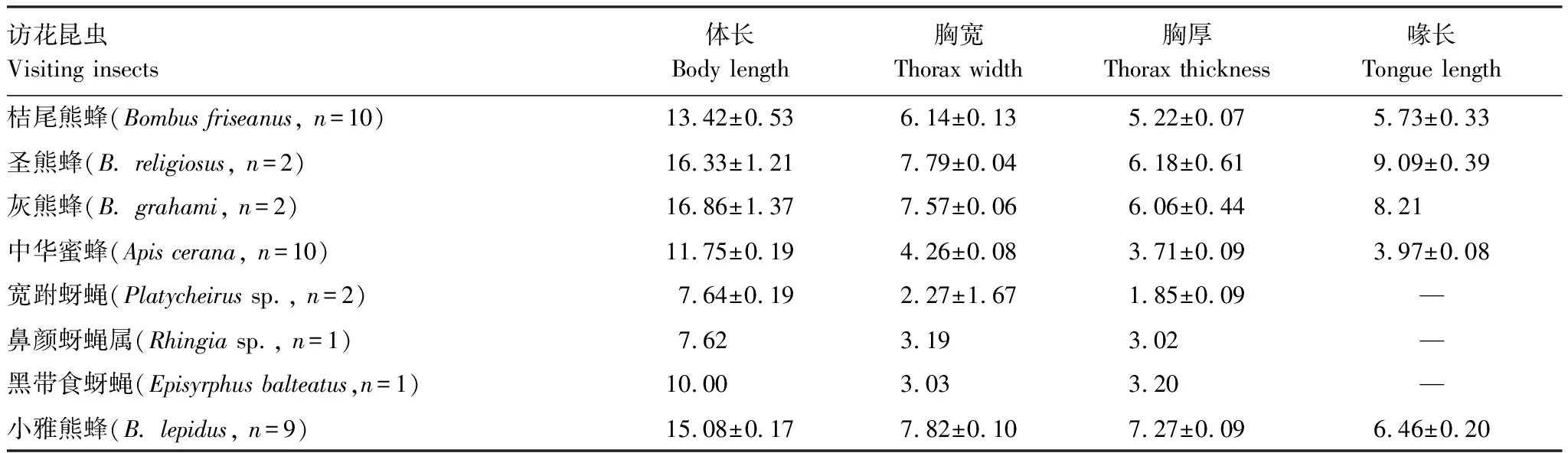
表4 2014年访花昆虫体型指标测量/mm
天气条件和长期的年际变化都可能影响传粉者的组成和行为变化[44,52—53]。在我们的研究中,2014年桔尾熊蜂在阴雨天有效传粉频率急剧下降(晴天∶阴雨天=2.59∶0.80),尽管盗蜜频率在阴雨天略高于晴天,但均维持在极低的水平(晴天∶阴雨天=0.07∶0.67),同时阴雨天的有效传粉和盗蜜频率也几乎没有差别(有效传粉∶盗蜜=0.80∶0.67)。另外,晴天天气没有影响灰熊蜂的有效传粉和盗蜜频率(有效传粉∶盗蜜=0.01∶0.03),阴雨天的盗蜜频率相比晴天发生了急剧增加(阴雨天∶晴天=2.43∶0.03)。因此桔尾熊蜂的有效传粉受阴雨天影响极大,盗蜜维持在较低水平,基本不受影响;同时灰熊蜂的盗蜜行为也显著受到阴雨天的影响,而传粉维持在较低水平,基本不受影响(表2)。两种天气条件下栗色鼠尾草的传粉特征主要体现在两种访花昆虫角色的替代和行为的改变。即晴天由桔尾熊蜂执行绝大部分的有效传粉工作,阴雨天由灰熊蜂替代桔尾熊蜂的工作,但却行使的是盗蜜行为。另一方面,桔尾熊蜂的行为变化主要与长期的年际变化相关,如2014年以有效传粉为主(晴天访花频次为65%),2015年以盗蜜为主(占所有访花频次的74%),到了2016年,桔尾熊蜂完全消失(访花频次为0)。而短期的天气变化仅显著影响其有效传粉,对盗蜜的频率没有影响。这种变化可能与桔尾熊蜂的生物学特性、鼠尾草的花器官年际变化或其他访花者的行为相关。灰熊蜂主要行使盗蜜行为,表面上看是受到了天气和年际变化的明显影响,本质上它的盗蜜行为可能很大程度上取决于桔尾熊蜂的访花行为以及出现频次,如两种熊蜂同时出现,资源竞争或领地争斗都可能引起行为的转变[45—47]。例如2014年灰熊蜂的盗蜜频率在阴雨天明显增加,可能与桔尾熊蜂在阴雨天的活动显著减少有关;到了2015年,尽管桔尾熊蜂访花行为发生显著改变,转变为盗蜜,但其出现频率仍很高,因而灰熊蜂的出现频率仍维持较低水平;至2016年桔尾熊蜂完全消失后,灰熊蜂的盗蜜频率发生了根本性的显著增加。而另一种有效传粉的圣熊蜂与灰熊蜂的行为相关性则不明显。从以上分析可以推测栗色鼠尾草的传粉效率和繁殖成功,取决于传粉者的有效传粉频率,在保证一定水平的有效传粉频率的基础上,繁殖成功则能够得到保障,而盗蜜只是有效传粉受到显著影响后的次生产物,对繁殖成功不会造成显著影响,可能只有当有效传粉下降至某一阈值,繁殖的有效性才明显下降,而传粉者与盗蜜者的互作关系,盗蜜行为存在的生态学意义,及其从植物和传粉者的角度如何理解其进化适应性,都有待更深入而系统地研究。
4 结论
连续3年的野外试验和观察,对广布于喜马拉雅山脉至横断山的栗色鼠尾草的传粉昆虫组成以及受环境气候条件影响下的年际间,以及晴天和阴雨天的传粉行为变化进行了研究。高山地区栗色鼠尾草访花昆虫和传粉昆虫多样,传粉者及其行为变化较为明显。栗色鼠尾草的盗蜜现象较为普遍,主要盗蜜昆虫为灰熊蜂和桔尾熊蜂。此外,天气条件显著影响桔尾熊蜂的传粉和灰熊蜂的盗蜜行为,桔尾熊蜂的行为变化受年际变化较大,同时灰熊蜂的盗蜜行为很大程度上与桔尾熊蜂的活动相关。栗色鼠尾草为专性异交,主要依靠传粉者进行授粉,缺少传粉者时低程度的主动自交可为其繁殖提供保障,且没有花粉限制和近交衰退。多变的天气条件可能显著影响着鼠尾草属植物的传粉者种类组成、访花行为和传粉效率,进而影响植物的繁殖成效和种群稳定。有效传粉频率是保证鼠尾草植物繁殖成功的主要因素,盗蜜对栗色鼠尾草的繁殖成功在一定水平维持中性的影响。
致谢:感谢杨飘,罗卷和李桂彬在野外实验中提供的帮助,马永鹏博士,任宗昕博士,谢东佑博士,金冬梅博士,蒋凯博士和胡超博士对写作给予帮助,特此致谢。
猜你喜欢
今日农业(2021年6期)2021-06-09
妇女生活(2019年3期)2019-03-18
中成药(2018年10期)2018-10-26
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
中成药(2017年7期)2017-11-22
中成药(2017年10期)2017-11-16
中学生天地(A版)(2017年10期)2017-10-28
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
Coco薇(2016年3期)2016-04-06
