
SLC30A8在糖尿病的研究进展
2022-03-21 02:30田雨薇邓琼严跃红王雨晨
广州医科大学学报 2022年6期
田雨薇,邓琼,严跃红,王雨晨
(广州医科大学附属第五医院,广州 510000)
1 SLC30A8的结构与表达
1.1 SLC30A8的结构
人类的SLC30基因家族有10个成员,至少有8个(SLC30A1,2,4,5,6,7,8,9)在人类β细胞中表达。SLC30A8基因[溶质载体家族30(锌转运蛋白),成员8]位于8q24.11,编码锌转运蛋白8(ZnT8),它包含8个外显子,全长41.72 kb。ZnT8蛋白由369个氨基酸组成,亲水分析表明,40 kDa ZnT8蛋白具有一个由6个α-螺旋组成的跨膜(TM)束[1]。其主要在胰岛β细胞中的胰岛素分泌颗粒膜上特异性高度表达,介导锌离子从胰腺β细胞的细胞质运输到胰岛素分泌颗粒中,从而对胰岛素的合成与储存进行调节[2]。在胰岛素囊泡中,胰岛素与2个锌离子结合形成较稳定的六聚体储存起来,当胰岛β细胞受到葡萄糖的刺激时,则将其分泌到循环中去。由此可见,SLC30A8基因编码的蛋白在锌离子介导的胰岛素成熟和分泌中发挥重要作用[3](见图1)。
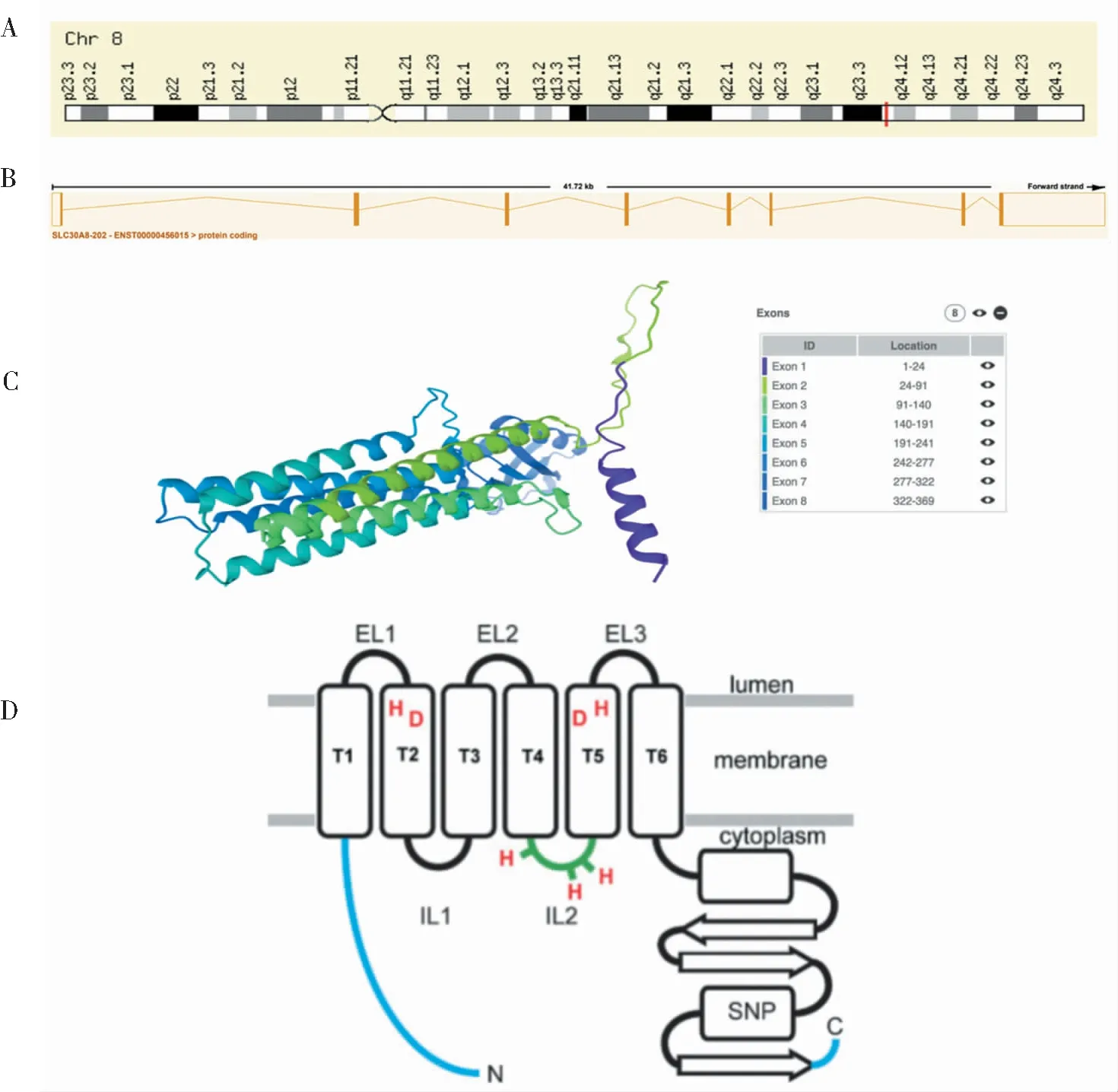
注:A:SLC30A8基因位于染色体8q24.11上;B:该基因包含8个外显子,全长41.72 kb,图中矩形表示外显子;C:ZnT8蛋白的AlphaFold预测模型,右上角图例:不同颜色表示外显子1-8,以及它们的相应位置;D:ZnT8的拓扑模型,指示R325W SNP、锌敏感的富含His 环(IL2)的位置,以及在哺乳动物ZnT蛋白中保守的TM 2和5中的一对 His(H)和Asp(D)残基[1]
1.2 SLC30A8的表达
SLC30A8最初被认为是胰腺特异性的,即高水平表达于胰腺内分泌腺中的β细胞,在α细胞中表达水平显著降低,不存在于胰腺外分泌腺中[4]。但后续研究表明在人体一些胰腺外部位也存在SLC30A8低表达,如视网膜色素上皮[5]、脂肪组织、淋巴细胞以及甲状腺滤泡和肾上腺皮质的立方上皮细胞。另外,在啮齿动物中的一些胰腺外内分泌腺中也检测到了SLC30A8的存在,包括垂体、肾上腺、颌下腺、甲状腺和睾丸。但在动物猪中,SLC30A8的表达似乎仅限于β细胞中[6]。
1.3 SLC30A8的多态性
迄今为止已鉴定出超过700个与T2DM 相关的基因位点,其中SLC30A8多态性是迄今发现的与2型糖尿病发生关联比较强的变异,尽管这种表型表现的分子机制尚不完全清楚,但这种多态性可以通过影响基因产物的表达水平来发挥它们的作用。2007年至今,全基因组关联研究(GWAS)已经在2型糖尿病的研究领域取得了重大进展。人类全基因组扫描技术目前已确定有近百个易感基因位点与2型糖尿病相关。从2007年Sladek在T2DM的个体中发现rs13266634的富集[7],到随后的研究中陆续确定了其他的SNP。rs13266634是SLC30A8基因的最后一个外显子的错义SNP,即碱基对位置973处的C被T取代,使位置325处的氨基酸从精氨酸(R)变为色氨酸(W)(Arg325Trp,R325W),该变异导致细胞中Zn2+转运活性下降[8]。SLC30A8中的其他SNP也与T2DM显著相关,如位于3’ 非翻译区的rs3802177和rs11558471以及rs16889462,它将氨基酸325从精氨酸(R)变为谷氨酰胺(Q)(Arg325Gln,R325Q)[9]。多态性rs3802177和rs11558471与rs13266634处于强连锁不平衡状态,两者也与T2D风险有关[10, 11]。随后Scott的研究也揭露了SLC30A8多态性和T2DM发展之间的潜在联系,值得注意的是其研究结果反驳了以往“存在多个常见SNPs对T2DM疾病有很大影响”的言论,Scott表示每个SNPs都具有适度的风险[12]。
1.4 SLC30A8的调控
目前动物实验中调节SLC30A8表达的因素有锌消耗、葡萄糖浓度的改变和缺氧、细胞因子暴露,以上因素均降低小鼠和大鼠胰岛衍生细胞系中SLC30A8基因的表达。首先,锌离子可以调节SLC30A8的表达。由于锌离子与胰岛素共结合共分泌,密切参与胰岛素生物合成和胰岛素分泌颗粒的成熟过程[13],细胞内锌的缺乏可以引起糖耐量的降低,胰岛素分泌颗粒减少,胰岛素含量降低。锌消耗时ZnT8的急剧下降已被证明会使大鼠胰岛细胞瘤细胞(INS-1)在葡萄糖刺激下的胰岛素分泌试验(GSIS)中的胰岛素分泌减少50%以上。此外,Slepchenko等[14]研究表明,细胞外锌水平也可以作为一种反馈机制来调节胰岛素的分泌,但其是否是通过影响SLC30A8的表达来调节,目前没有具体文章阐述。与此一致的是,过量锌被证明会导致胰岛素基因的表达减弱以及细胞活力的降低。局部升高的锌离子浓度通过诱导细胞凋亡导致胰腺细胞死亡,而游离锌的减少则与细胞中胰岛素含量的减少有关。除了锌离子可以作用于SLC30A8的表达,在高葡萄糖浓度下也发现了ZnT8表达的下调[15]。同时缺氧和细胞因子暴露会介导ZnT8的下调,通过降低细胞内锌含量,从而降低细胞死亡率[16]。细胞因子暴露会进一步使β细胞容易积聚错误折叠的蛋白质,从而加剧内质网应激。研究表明,人类β细胞在急性细胞因子暴露下,细胞中ZnT8水平会出现快速、分级和可逆的下调[17]。因此,SLC30A8能对动物体内葡萄糖浓度变化和锌消耗、细胞因子暴露、缺氧作出反应,且对β细胞应激期间的胰岛素分泌和体内葡萄糖代谢至关重要。
2 SLC30A8的功能
2.1 SLC30A8与胰岛分泌功能
与糖尿病相关的现有基因位点大多都与胰岛素分泌有关,而与胰岛素抵抗无关。关于SLC30A8功能缺失对小鼠胰岛分泌功能影响的报道结论不一。一些研究表明SLC30A8功能缺失后β细胞功能恶化,另一些研究表明没有净效应,还有一些研究则提示胰岛分泌功能有改善。
大多数β和α细胞特异性ZnT8敲除(ZnT8BKO和ZnT8AKO)小鼠或ZnT8整体敲除(ZnT8-/-)小鼠模型中,SLC30A8编码的ZnT8失活都能引起较大的表型变化,导致胰岛素颗粒内致密核心形成异常、葡萄糖耐量不同程度受损,葡萄糖刺激下的胰岛素分泌功能显示受损或无变化[18]。最新的hIAPP(人胰岛淀粉样多肽)转基因小鼠中,缺乏ZnT8导致β细胞中淀粉样蛋白积累增加,β细胞功能恶化,细胞同步性受损[19],这表明ZnT8活性不仅对胰岛素生物合成和β细胞功能很重要,而且对保护β细胞免受淀粉样蛋白聚集也很重要。也有研究报道,当β细胞中ZnT8过表达时,虽然小鼠体内葡萄糖耐量得到改善,但葡萄糖刺激的胰岛素分泌是减少的[20]。早于此项研究以前,人们认为ZnT8过表达刺激了锌的积累,并增强了葡萄糖刺激下的胰岛素分泌。所以目前关于ZnT8过表达对胰岛功能到底是起到增强还是减退的效果,仍存在争议。另外,Karsai等[16]在ZnT8-/-胰岛的基础上用细胞因子进行干预,在细胞因子暴露后,ZnT8-/-胰岛的胰岛素分泌下降到与正常胰岛相似的程度。
人类SLC30A8多态性研究中也可见,携带rs 13266634C等位基因者表现出胰岛素分泌的显着减少[21],同时可能有胰岛素原-胰岛素转化效率的下降。由于胰岛素原向胰岛素的转化受到Zn2+的抑制[22],合成ZnT8缺乏会影响胰岛素敏感组织可用的胰岛素水平,这与直接下调ZnT8表达得到的结果一样。其原理可能为增大胰岛β细胞凋亡或者降低增殖使β细胞质量下降,造成胰岛素原在转变为胰岛素分子时受到损坏,进而体现为胰岛素分泌功能的减退。
但也有研究表明,SLC30A8功能缺失对胰岛分泌功能有改善。在上述Karsai的研究中,细胞因子干预前的ZnT8-/-胰岛在葡萄糖刺激下的胰岛素分泌是增加的。近期研究报道,ZnT8参与了人类胚胎干细胞(hESCs)衍生的β细胞(SC-β细胞)的功能成熟过程,其功能缺失(LOF)会加速SC-β细胞的功能成熟,因此ZnT8 LOF在体内外均改善葡萄糖刺激下的胰岛素分泌,并提高SC-β细胞的胰岛素原-胰岛素转化效率[23]。Sandra Kleiner等[24]研究也表明,在携带SLC30A8 R138X LOF突变的小鼠模型中,高糖状态下的R138X小鼠胰岛素分泌增加了50%。以上研究指向同一个结论,即SLC30A8功能缺失突变可通过增加细胞在高糖条件下分泌胰岛素的能力而对葡萄糖代谢产生有益影响,这意味着随着ZnT8的功能丧失,β细胞的胰岛素分泌增加降低了患糖尿病风险。在对携带SLC30A8 W325或截短变体的人类受试者中进行的研究同样表明,口服葡萄糖后激发胰岛素分泌增加[25],这一发现与前期动物实验所得到的结论形成鲜明对比。
在另一种胰腺细胞——α细胞中,ZnT8过表达使胰高血糖素分泌受阻以应对低血糖[26],相反ZnT8敲低则会刺激α细胞分泌胰高血糖素,这提示胰高血糖素分泌可能直接受ZnT8调节,从而间接影响胰岛素分泌。
总的来说,有关SLC30A8与胰岛分泌功能的关系在啮齿动物模型中的研究呈现出一幅复杂的图景,可能无法概括实际上SLC30A8与人类的联系。这种差异会导致未来更多研究以人类受试者为基础,集中揭示SLC30A8在人类β细胞功能中的作用。
2.2 SLC30A8与空腹血糖、餐后血糖
空腹血糖受损(IFG)是临床定义的糖尿病前期状态,25%的IFG患者在3至5年内进展为2型糖尿病。所以进行遗传信息的评估能有效预防糖尿病的发生发展。吴英在基于中国汉族人群的基础上证实了SLC30A8的多态性(SNP)与空腹血糖受损的风险相关,且这些SNP中的大多数与受损的β细胞功能有关[11]。既往研究表明,影响2型糖尿病风险和基线空腹血糖水平的基因位点与10年内空腹血糖的非显著增加相关联[27],但也有人表示,空腹血糖和影响空腹血糖的遗传变异之间的关联随着时间的推移是稳定的,类似于非糖尿病个体的年龄相关变化[28]。尽管SLC30A8风险等位基因对糖尿病的影响贯穿整个病程,但Walford[29]等研究表明,相较于已患2型糖尿病人群,SLC30A8在未患糖尿病但空腹血糖受损人群中更广泛存在,即SLC30A8的生物学效应对糖尿病前期空腹高血糖的发展影响更大。也有研究表示SLC30A8基因变异并不能提示空腹血糖受损。Dewivedi[25]等对SLC30A8功能丧失变异等位基因(p.Arg138*)的人类载体杂合子进行代谢研究,发现p.Arg138*携带者空腹葡萄糖浓度较非携带者更低。
常见的遗传变异会导致餐后2小时血糖水平与年龄相关性的升高[28]。全基因组关联研究和既往36项荟萃分析表明,SLC30A8基因的错义SNP R325W变体与葡萄糖调节受损(IGR)和T2DM风险增加有关[30]。在EUGENE2研究中,SLC30A8中常见的遗传风险C变异在不同程度上影响空腹血糖受损的发展、从IFG向T2DM的转变以及葡萄糖代谢受损。研究表明,在口服葡萄糖耐量试验期间,胰岛素原与胰岛素的比率增加,而在静脉葡萄糖耐量试验期间则出现胰岛素分泌受损以及β细胞功能降低[31],这表明SLC30A8可能调节胰岛素稳态。Dong 等[32]在以中国人群为研究背景时也得到相同结论。然而,也有从另一角度进行研究发现R325W变体增加了锌转运蛋白的活性[33],该变体可以被靶向抑制以降低患2型糖尿病的风险。
3 SLC30A8与糖尿病
3.1 SLC30A8与1、2型糖尿病
1型糖尿病(T1DM)是一种自身免疫性疾病。除了胰岛细胞质自身抗体(ICA)、胰岛素抗体(IAA)、65kD形式的谷氨酸脱羧酶(GADA)和蛋白质酪氨酸磷酸酶IA2(IA2A)这些自身抗体可用于识别80%以上T1D患者的发病或发展风险外,目前SLC30A8编码的ZnT8也成为了一种用于识别T1DM的新型胰岛自身抗原,并作为该病的一项相关预后特征。ZnT8自身抗体(ZnT8A)在前驱糖尿病阶段持续存在,此外,还存在于2%的非糖尿病患者、3%的T2DM患者和30%的其他自身免疫疾病患者,60%~80%的新发T1DM患者中,并且在T1D新诊断儿童中,存在42%的ZnT8A阳性[34]。如果同时结合GADA、IA2A、IAA和ZnT8A检测,自身免疫检测率将在疾病发作时上升至98%。行ZnT8A检测在胰岛素自身抗体随年龄增长而减弱的老年人中尤其有用。另外,ZnT8A相较于其他抗体具有高度的β细胞特异性,因此ZnT8A可用于监测发病后的胰岛破坏,以及评估限制β细胞特异性自身反应或进行恢复β细胞质量的干预治疗。
2型糖尿病(T2DM)是由遗传因素和环境因素共同作用而形成的多基因遗传病,是一种复杂的异质性的糖代谢性疾病,其发病机制是多因素的,主要包括高血糖反应、胰岛功能受损和(或)胰岛素分泌障碍。研究证明,SLC30A8多态性与2型糖尿病风险有着密切联系,不同地理分布和文化因素导致了SLC30A8 SNP对2型糖尿病的影响在全世界的流行分布存在差异[35]。欧洲人群中所鉴定SLC30A8基因中的SNP,与西班牙裔美国人和非洲裔美国人的葡萄糖稳态特征相关的适度证据,提示SLC30A8的糖尿病风险是通过胰岛素分泌缺陷特异性介导。Nikitin等[21]研究发现,在俄罗斯人群中,SLC30A8基因 SNP rs13266634与葡萄糖代谢受损或β细胞功能受损显着相关,影响胰腺β细胞胰岛素合成和分泌,在T2DM的发展中起着重要作用。视线转向亚洲,Mashal等[36]证明在约旦人群中 C/T rs13266634 SNP 与 T2DM 之间存在显着关联,同样在日本人群中,SLC30A8多态性与2型糖尿病、空腹血浆胰岛素水平相关联。在中国人群中,一项荟萃分析也同样确认了来自SLC30A8 SNP对2型糖尿病的影响,表明了SLC30A8 rs13266634 是 T2DM 的潜在危险因素[32]。但并非所有人类研究都可以确定SLC30A8多态性与T2DM易感性风险之间的联系。上述SLC30A8 SNP可能是多数亚洲和欧洲人群2型糖尿病风险的重要遗传因素,不包括所有的非洲人群[37],在墨西哥裔美国家庭[38]和部分欧洲血统的非洲裔美国人中也没有发现T2DM与SLC30A8多态性相关。以上结果的不一致可能是由于研究采集样本量小,且种族不相同[37]。
尽管多数研究证明SLC30A8基因中常见的单核苷酸多态性使2型糖尿病风险升高,但该基因中罕见的功能丧失(LOF)突变可能会有意外的保护作用,这项结论与人类和小鼠模型中的早期工作所得的结论恰恰相反。Flannick等[39]报告了15万个基因型中约350个罕见的SLC30A8功能丧失突变,可以降低T2DM风险。研究中确定的12种抑制性功能丧失变体共同使T2DM风险降低了65%。常见于芬兰西部的变体之一p.Arg138*,显示SLC30A8表达明显减少[25],其编码一种不稳定的蛋白质,对T2D具有53%的保护作用。除此之外,Flannick还报告了一个保护性移码等位基因p.Lys34Serfs50*,在冰岛人群中提供了83%的T2D保护。尽管主要的p.Arg325等位基因(>70%的人群)会增加患T2D的风险,但次要的p.Trp325等位基因仍具有保护性,但保护作用的确切机制尚未确定[18]。
3.2 SLC30A8与糖尿病发病机制
尽管SLC30A8多态性与2型糖尿病的易感性增加有关,但其在全身能量代谢中的确切机制仍不清楚。回顾当前各项研究将SLC30A8中的多态性与T2DM风险改变联系起来的潜在机制包括:胰岛素分泌减少、胰岛素清除率增加以及对活性氧物质的敏感性增加[18](见图2)。

图2 SLC30A8在糖尿病发生发展机制图
一方面,SLC30A8多态性可以通过降低胰岛β细胞数量和质量,影响胰岛素合成途径,导致胰岛素分泌功能减退。最新研究发现,缺乏ZnT8的β细胞中存在人胰岛淀粉样多肽(hIAPP)会加剧淀粉样蛋白生成和β细胞功能障碍[19],hIAPP聚集体诱导的毒性会导致β细胞面积和功能的损失,构成了SLC30A8影响糖尿病发病机制之一。还有动物实验表明,部分急性的ZnT8抑制会损害4周龄小鼠的葡萄糖耐量,这种损害与基础胰岛素清除率增加和胰岛素分泌延迟有关,但这种胰岛功能的损伤是短暂的,并在发育过程中发生补偿性适应[40]。另一方面,当ZnT8功能下降时,肝脏胰岛素清除失调,也可能增加2型糖尿病风险。Tamaki等[2]报道了 ZnT8B-KO小鼠的血浆胰岛素降低,是由于胰岛素的肝脏清除率增强所致。此项研究表明,与胰岛素协同分泌的锌通过抑制网格蛋白依赖性胰岛素内吞作用来抑制肝脏胰岛素清除。另外也有报道提出,在胰腺组织之外出现的ZnT8有助于调节肠道内分泌细胞中的5-羟色胺生物合成,从而改变全身葡萄糖和脂质代谢,最终导致T2DM 的敏感性增加[41]。鉴于这些可能的机制,SLC30A8/ZnT8已被提出作为个性化疾病治疗的潜在易处理新靶点。
通过检测基因我们能够确定重要的遗传因素,在早期阶段预测糖尿病,避免后续的并发症。然而目前有关SLC30A8的研究仍存在以下几个方面的不足,(1)多数研究集中于欧美人群,不同地域和种族内SNP位点分布的基因频率差异也很大,需要后续进一步扩大研究范围。(2)对于SLC30A8基因多态性和国内人群T2DM易感性的相关报道仍有存疑,还需要进一步的验证以获取真实性[42]。(3)存在于胰腺外组织中的SLC30A8对生物体能量稳态也起着不可忽视的作用,具体机制值得我们去探讨。
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22
实用器官移植电子杂志(2022年5期)2022-03-24
中老年保健(2021年3期)2021-08-22
恋爱婚姻家庭(2021年17期)2021-07-27
生命科学研究(2018年1期)2018-05-29
小学生导刊(低年级)(2016年8期)2016-09-24
西南农业学报(2016年6期)2016-04-16
中央民族大学学报(自然科学版)(2015年1期)2015-06-11
肝胆胰外科杂志(2015年1期)2015-02-27
中国医学科学院学报(2014年6期)2014-03-11
