
低温胁迫对3种蕨类植物生理指标的影响
2022-03-15 04:04毛林鲜朱芸琦吴美丽
西南农业学报 2022年12期
毛林鲜,朱芸琦,吴美丽,刘 建,区 智,2
(1.西南林业大学园林园艺学院/国家林业和草原局西南风景园林工程技术研究中心, 昆明 650224;2.西南山区森林资源保护与利用教育部重点实验室/西南林业大学, 昆明 650224)
【研究意义】低温作为一种最常见、最主要的非生物胁迫因素,对植物的生长发育与地理分布有着极大的影响[1]。蕨类植物也被称为羊齿植物,现存蕨类植物约12 000种,大多数主要原分布于热带及亚热带地区,温度是影响蕨类植物地理分布的一个重要因子。因此,对低温的适应是蕨类植物维持种群生长的关键。大多数蕨类植物对低温环境非常敏感,在引种驯化时,低温通常是一个限制性因素。鸟巢蕨(Aspleniumnidus)、波士顿蕨(Nephrolepisexaltatacv. Bostoniensis)和狼尾蕨(Davalliatrichomanoides)均是具有优良观叶价值的蕨类植物,常在园林景观中用作地被植物和室内的观叶盆栽植物,3种蕨类植物喜温暖湿润遮荫环境,均不耐强光和低温。低温会对鸟巢蕨、狼尾蕨、波士顿蕨3种蕨类植物的叶片造成不同程度的损伤,影响其观赏价值。【前人研究进展】当温度处于植物正常生长温度以下时,植物的生长受到影响,最终致使植株死亡。相关研究发现低温胁迫能使植物体内渗透调节物质积累[2]。田丹青等[3]研究发现3个红掌(Anthuriumandraenum)品种在低温胁迫处理后叶片的丙二醛(MDA)含量呈先降低后上升的变化趋势,叶片相对电导率上升。赵一航等[4]对4个苜蓿(Medicagosativa)品种的研究发现,低温胁迫下超氧化物歧化酶和过氧化物酶酶活性均有上升的现象。黄一波等[5]研究发现低温胁迫下7种木莲属(Manglietia)植物的可溶性糖和可溶性蛋白含量上升。马晓华等[6]研究发现低温胁迫后2个蝴蝶兰(Phalaenopsisamabilis)品种叶片的部分光合参数均有所下降。【本研究切入点】目前,关于蕨类植物的研究主要集中在繁育、资源考察等方面,对于蕨类植物抗性生理方面的研究并不多[7]。迄今为止,关于蕨类植物抗性生理的研究又限于干旱胁迫、重金属胁迫、盐胁迫、耐阴性等对其生理生化指标的影响[8-13],对蕨类低温胁迫的研究鲜见报道。【拟解决的关键问题】本研究以鸟巢蕨、狼尾蕨和波士顿蕨3种蕨类植物为研究对象,通过测定3种蕨类植物在低温胁迫下相关生理生化指标及光合参数,初步探究其生理响应机制,比较3种蕨类植物的抗寒性,筛选出较耐寒的种类,为今后蕨类植物的育种、园林运用和栽培养护提供理论依据。
1 材料与方法
1.1 试验材料
以长势良好且一致、无病虫害的一年生鸟巢蕨、狼尾蕨和波士顿蕨为试验材料,试验材料培养在塑料盆(口径为10 cm,高为8.7 cm,底径为6.8 cm)中,培养基质中泥炭土、珍珠岩和蛭石的配比是3∶1∶1,日常进行正常的水肥管理及病虫害防治等栽培养护工作。
1.2 处理方法
先将植株放在恒温20 ℃,光照12/12 h(昼/夜),光照强度为4000 Lx,湿度为70%~75%的光照培养箱中预培养2 d,随后放入0 ℃的光照培养箱中进行处理,分别在0 ℃处理0 h(CK)、48 h、96 h以及恢复常温20 ℃ 48 h时对植株相同部位叶片进行采样及光合参数的测定,设置3个生物学重复,取样后立即进行相对电导率的测定,其余指标对应每个指标取样品0.1 g包裹在锡箔纸中,液氮速冻后,用自封袋装好放入-80 ℃冰箱中冷冻贮存,以便后期测定相关生理指标。
1.3 生理指标测定方法
叶绿素含量、相对电导率、丙二醛(MDA)含量、可溶性蛋白质含量、可溶性糖含量、超氧化物歧化酶(SOD)和过氧化物酶(POD)活性等指标的测定方法均参照前人方法用T-6新世纪紫外可见分光光度计进行测定[14]。用LI-6400XT便携式光合仪(LI-COR,美国)在8:30—11:30时测定净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)、胞间CO2浓度(Ci)及水分利用率(WUE,WUE=Pn/Tr)等光合参数相关的光合指标,使用红蓝光源进行测量,光强设定为1000 μmol/(m2·s),CO2浓度为外界CO2浓度,相对湿度为70%,流速设置为500 μmol/s,叶室温度为(25±1)℃,每片叶子重复测量3次后取平均值进行后期数据分析。
1.4 主成分分析和综合评价
采用前人方法对各项生理及光合指标进行主成分分析[15-16]。
(1)
式中,Zj表示某蕨类植物第j个主成分综合得分值,Bij表示第j个主成分第i个指标的特征向量值,xi表示某蕨类植物第i个指标的测定值。
利用以下公式计算低温胁迫下3种蕨类植物耐受性的综合评价D值[17]。
(2)
式中,U(xi)表示各蕨类第i个主成分的隶属函数值;Wi表示第i个主成分在所有主成分中的权重。
隶属函数值的计算参考前人方法[18],分析和评价3种蕨类植物的抗寒性。分别使用公式(3~4)计算与抗寒性呈正相关和负相关的指标:
隶属函数=(Xi-Xmin)/(Xmax-Xmin)
(3)
反隶属函数=1-(Xi-Xmin)/(Xmax-Xmin)
(4)
式中,Xi为指标测定值,Xmin为该指标测定值中的最小值,Xmax为该指标测定值中的最大值。
1.5 数据处理
用Microsoft Excel 2010和SPSS 22.0软件对数据进行数据统计和单因素方差分析(One-way ANOVA),采用Origin 2021作图。
2 结果与分析
2.1 低温胁迫对3种蕨类植物叶片生理生化指标和光合参数的影响
2.1.1 叶绿素含量 如图1-A所示,低温胁迫后,3种蕨类叶片的叶绿素含量变化趋势不同。随胁迫处理时间的延长,鸟巢蕨叶片的叶绿素含量呈上升趋势,波士顿蕨叶片的叶绿素含量变化不明显,狼尾蕨叶片的叶绿素含量则呈先下降后上升趋势。鸟巢蕨叶片的叶绿素含量在胁迫处理96 h 达到最大值,为2.53 mg/g FW,较0 h(CK)上升32.20%。波士顿蕨叶片的叶绿素含量在胁迫处理96 h 降至最小值,为2.34 mg/g FW,较0 h(CK)下降14.55%,恢复常温20 ℃ 48 h后有所回升。狼尾蕨叶片的叶绿素含量在胁迫处理48、96 h及恢复48 h与0 h(CK)相比均有所下降,较0 h(CK)分别下降21.98%、7.17%和19.08%。狼尾蕨叶片的叶绿素含量在各个时间点均显著(P<0.05)高于鸟巢蕨和波士顿蕨。综上表明,3种蕨类植物对低温胁迫作出了不同响应,导致其叶片的叶绿素含量变化规律不一致。
2.1.2 相对电导率 如图1-B所示,随着低温胁迫处理时间的延长,3种蕨类叶片的相对电导率均呈上升趋势,但三者的增幅存在较大差异。在胁迫处理48和96 h时,与0 h(CK)相比,鸟巢蕨分别上升10.57%和121.62%,波士顿蕨分别上升83.33%和198.62%,狼尾蕨分别上升16.94%和76.41%。3种蕨类在受到胁迫时存在显著差异(P<0.05),在胁迫处理48和96 h时波士顿蕨的相对电导率显著高于鸟巢蕨和狼尾蕨(P<0.05)。三者均在胁迫处理96 h 达到最大值,显著高于0 h(CK)(P<0.05)。恢复常温20 ℃ 48 h时鸟巢蕨、波士顿蕨和狼尾蕨叶片的相对电导率与胁迫96 h相比显著下降(P<0.05),分别下降60.11%、57.70%和54.47%,说明低温解除后,低温胁迫对3种蕨类植物的伤害程度减轻,细胞内外渗的电解质减少。
2.1.3 MDA含量 MDA是膜脂过氧化物的产物之一。如图1-C所示,3种蕨类的MDA含量在各个时间点均存在显著差异(P<0.05),且在各个时间点狼尾蕨叶片的MDA含量均显著高于波士顿蕨。随着低温胁迫处理时间的延长,鸟巢蕨叶片的MDA含量呈上升趋势,波士顿蕨和狼尾蕨叶片的MDA含量变化呈下降趋势。鸟巢蕨叶片的MDA含量在胁迫处理96 h 时到达峰值,为10.06 mmol/g FW,较0 h(CK)上升53.19%,恢复常温20 ℃ 48 h时相比胁迫处理96 h时有所下降。波士顿蕨和狼尾蕨叶片的MDA含量在胁迫处理96 h 均降低到最小值,分别为0.52和3.36 mmol/g FW,较0 h(CK)分别下降88.90%和69.47%,
2.1.4 可溶性糖含量 从图1-D可知,随着低温胁迫处理时间的延长,3种蕨类叶片中可溶性糖含量均呈上升趋势。在胁迫处理48和96 h时,与0 h(CK)相比,鸟巢蕨分别上升184.10%和390.80%,波士顿蕨分别上升98.96%和110.93%,狼尾蕨分别上升127.66%和138.45%,三者均在胁迫处理96 h达到最大值,显著高于0 h(CK)(P<0.05)。狼尾蕨在每个时间点的可溶性糖含量均显著高于其他2种蕨类(P<0.05)。恢复常温20 ℃ 48 h时鸟巢蕨、波士顿蕨和狼尾蕨叶片的可溶性糖含量与胁迫96 h显著下降(P<0.05),分别下降51.83%、52.50%和57.29%,除鸟巢蕨外,其他2种蕨类均恢复到0 h(CK)水平,与0 h(CK)相比无显著差异(P>0.05)。
2.1.5 可溶性蛋白含量 从图1-E可知,随着低温胁迫处理时间的延长,3种蕨类叶片中可溶性蛋白含量均呈上升趋势。在胁迫处理48和96 h时,与0 h(CK)相比,鸟巢蕨分别上升43.83%和275.42%,波士顿蕨分别上升11.84%和99.31%,狼尾蕨分别上升5.59%和268.33%,鸟巢蕨上升的幅度大于其他2种蕨类。三者均在胁迫处理96 h 达到最大值,显著高于0 h(CK)(P<0.05)。恢复常温20 ℃ 48 h时鸟巢蕨、波士顿蕨和狼尾蕨叶片的可溶性蛋白含量与胁迫96 h相比显著下降(P<0.05),分别降低13.42%、21.60%和19.03%。
2.1.6 SOD活性 从图1-F所示,随着低温胁迫处理时间的延长,3种蕨类植物SOD活性的变化趋势不同,鸟巢蕨和波士顿蕨呈上升趋势,狼尾蕨SOD活性变化不明显。在胁迫处理48和96 h时,与0 h(CK)相比,鸟巢蕨分别上升16.53%和38.25%,波士顿蕨分别上升133.16%和43.50%。3种蕨类植物SOD活性出现峰值的时间点不同,鸟巢蕨和狼尾蕨在低温胁迫96 h到达峰值,波士顿蕨在低温胁迫48 h到达峰值。恢复常温20 ℃ 48 h时鸟巢蕨和波士顿蕨叶片的SOD活性与胁迫96 h相比显著下降(P<0.05),分别降低39.60%和9.32%。
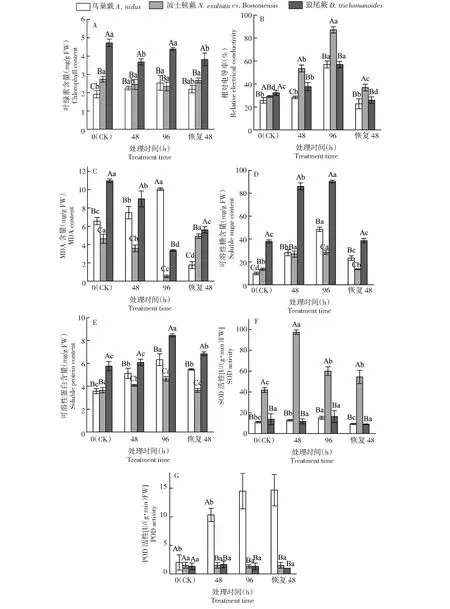
不同大写字母表示不同蕨类在同一时间点下差异显著(P<0.05),不同小写字母表示同一蕨类在不同时间点下差异显著(P<0.05);恢复48 表示低温胁迫后恢复常温(20 ℃) 48 h,下同Different capital letters indicate significant differences between fern species at the same time (P<0.05), and different lowercase letters indicate the significant difference of the same fern species at different time (P<0.05). Recovery 48 indicates 48 hours recovered at air temperature (20 ℃) after low temperature stress, the same as below图1 低温胁迫对3种蕨类植物生理生化指标的影响Fig.1 Effects of low temperature treatments on physiological biochemical indexes of three fern species
2.1.7 POD活性 由图1-G可以看出,随低温胁迫处理时间的延长,鸟巢蕨的POD活性呈升高趋势,波士顿蕨和狼尾蕨POD活性变化不明显。鸟巢蕨的POD活性在胁迫48、96 h及恢复48 h时与0 h(CK)相比分别上升416.67%、625.00%和633.33%。鸟巢蕨叶片的POD活性变化幅度远高于其他2种蕨类植物。
2.1.8 光合参数 由图2-A可知,在低温胁迫下,3种蕨类植物叶片的净光合速率Pn均呈下降趋势。鸟巢蕨、波士顿蕨和狼尾蕨叶片的净光合速率Pn在胁迫处理48 h时出现大幅度下降,与0 h(CK)相比,分别下降30.52%、76.10%和84.93%,狼尾蕨下降的幅度最大。在胁迫处理96 h下降至最小值,与0 h(CK)相比,分别下降50.64%、90.13%和92.28%,此时,鸟巢蕨叶片的净光合速率Pn显著高于波士顿蕨和狼尾蕨,狼尾蕨的下降幅度最大。恢复常温20 ℃ 48 h后鸟巢蕨、波士顿蕨和狼尾蕨叶片的净光合速率Pn与胁迫处理96 h时相比均有大幅度回升,分别上升54.04%、470.42%和868.78%。
由图2-B可知,随着低温胁迫处理时间的延长,3种蕨类植物叶片的蒸腾速率Tr均呈下降趋势,但下降的时间点有所差异。低温胁迫48 h时,狼尾蕨叶片的蒸腾速率Tr大幅下降,与0 h(CK)相比下降68.96%,且显著低于鸟巢蕨、波士顿蕨,而鸟巢蕨、波士顿蕨与0 h(CK)相比无显著变化。胁迫处理96 h时,3种蕨类植物叶片的蒸腾速率Tr均下降至最小值,与0 h(CK)相比鸟巢蕨、波士顿蕨和狼尾蕨分别下降59.29%、63.11%和78.99%,此时,鸟巢蕨、波士顿蕨显著高于狼尾蕨(P<0.05)。恢复常温20 ℃ 48 h后鸟巢蕨、波士顿蕨和狼尾蕨叶片的蒸腾速率Tr与胁迫处理96 h时相比均有大幅度回升,分别上升81.67%、75.45%和393.40%。综上,狼尾蕨叶片的蒸腾速率Tr受低温胁迫的影响更大。
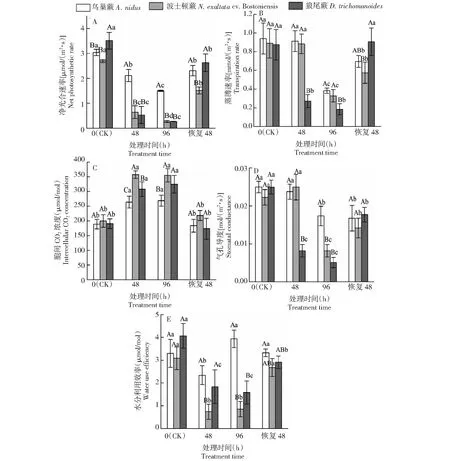
图2 低温胁迫对3种蕨类植物光合参数的影响Fig.2 Effects of low temperature treatments on photosynthetic parameters of three fern species
由图2-C可知,随着低温胁迫处理时间的延长,3种蕨类叶片的胞间CO2浓度Ci均呈上升趋势。在胁迫处理48 h和96 h时,与0 h(CK)相比,鸟巢蕨分别上升39.73%和42.19%,波士顿蕨分别上升78.86%和77.76%,狼尾蕨分别上升61.69%和70.33%,三者均在胁迫处理96 h 达到最大值,显著高于0 h(CK)(P<0.05)。波士顿蕨在胁迫处理48 h的胞间CO2浓度Ci显著高于其他两种蕨类(P<0.05)。恢复常温20 ℃ 48 h时鸟巢蕨、波士顿蕨和狼尾蕨叶片的胞间CO2浓度Ci与胁迫96 h相比显著下降(P<0.05),分别下降31.69%、38.50%和46.57%。
由图2-D可知,低温胁迫下,3种蕨类植物叶片的气孔导度Gs总体呈下降趋势,均在胁迫处理96 h 时大幅度下降至最小值,与0 h(CK)相比分别下降30.53%、63.29%和79.58%。此时,3种蕨类植物叶片气孔导度均显著低于0 h(CK)(P<0.05),鸟巢蕨叶片气孔导度显著高于波士顿蕨和狼尾蕨(P<0.05)。恢复常温20 ℃ 48 h后波士顿蕨和狼尾蕨叶片的气孔导度Gs与胁迫96 h相比显著上升(P<0.05),分别上升73.66%和247.33% 。
由图2-E可知,低温胁迫下,鸟巢蕨、波士顿蕨和狼尾蕨叶片水分利用效率WUE总体上呈下降趋势,但出现最低值的时间点有所差别。鸟巢蕨和波士顿蕨叶片的水分利用效率WUE均在48 h 下降到最低值,为2.34和0.74 μmol/mol,较0 h(CK)相比分别下降30.53%和76.05%。在胁迫48 h时,狼尾蕨叶片水分利用效率WUE下降到最低值,为1.84 μmol/mol,较0 h(CK)相比下降61.12%。恢复常温20 ℃ 48 h后波士顿蕨和狼尾蕨叶片水分利用效率WUE与胁迫96 h相比显著上升(P<0.05),分别上升213.67%和84.13%。
2.2 低温胁迫下3种蕨类植物各生理指标的主成分分析
采用Origin 2021对测定的指标共12个进行主成分分析(PCA分析),第一主成分的贡献率是43.0%,第二主成分的贡献率是25.7%。在第一主成分中,最大的特征向量是Pn,其次是Tr;在第二主成分中,最大的特征向量是叶绿素含量,其次是可溶性蛋白和可溶性糖;第一、二主成分主要反映低温胁迫下3种蕨类植物的光合能力、水分利用能力和渗透调节能力受到影响。PC1对POD、MDA、WUE、Pn、Gs、Tr均有正向荷载,对Chl、SP、SS、REC、Ci、SOD均有负向荷载。PC2对POD、MDA、WUE、Pn、Chl、Tr、SP、SS均有正向荷载,对REC、Ci、SOD、Gs、Tr均有负向荷载(图3)。
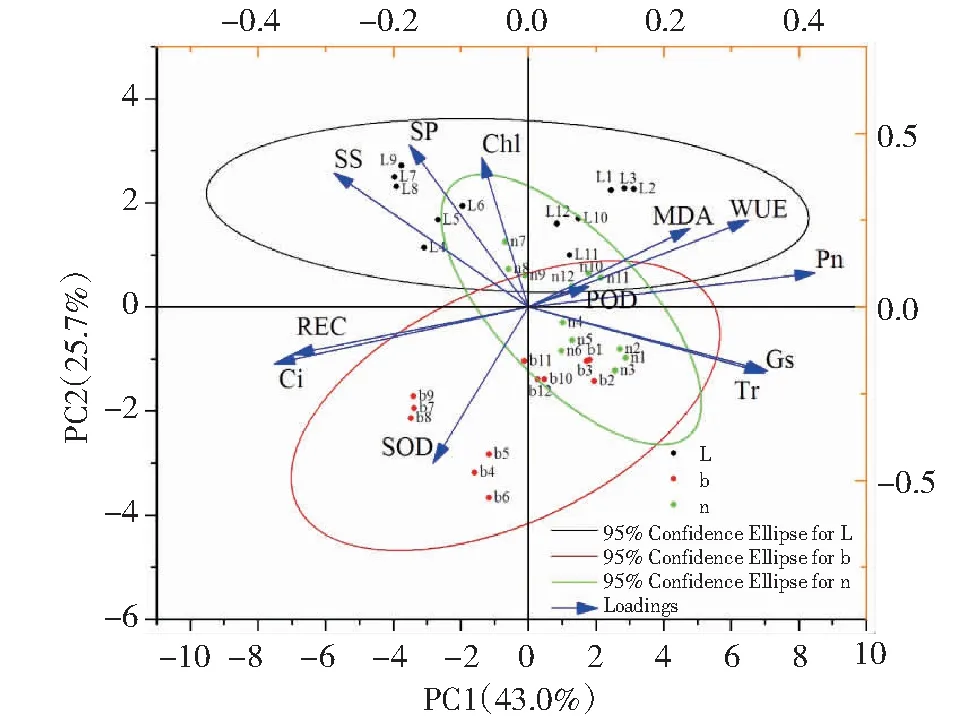
L:狼尾蕨;b:波士顿蕨;n:鸟巢蕨;1~3代表0 h;4~6代表48 h;7~8代表96 h;9~12代表恢复48 hL: D.trichomanoides; b: N.exaltata cv. Bostoniensis; n: A.nidus; 1-3 represent 0 hours; 4-6 represent 48 hours; 7-9 represent 96 hours; 10-12 represent recovery for 48 hours图3 低温胁迫下3种蕨类植物各生理指标的主成分分析Fig.3 Principal component analysis of different indexes of three fern species under low temperature treatments
2.3 低温胁迫下3种蕨类植物的耐受性综合评价
利用公式(1)可得出3种蕨类3个主成分的得分值Z(1)、Z(2)和Z(3)。根据公式(3~4)可算出3种蕨类的隶属函数值U(x),综合评价D值用公式(2)算出,D值越大,表明低温胁迫下耐受性越强,D值越小则越弱。由表1可知,用低温胁迫下3种蕨类D值大小对其进行耐受性排序,可得出:鸟巢蕨>狼尾蕨>波士顿蕨。
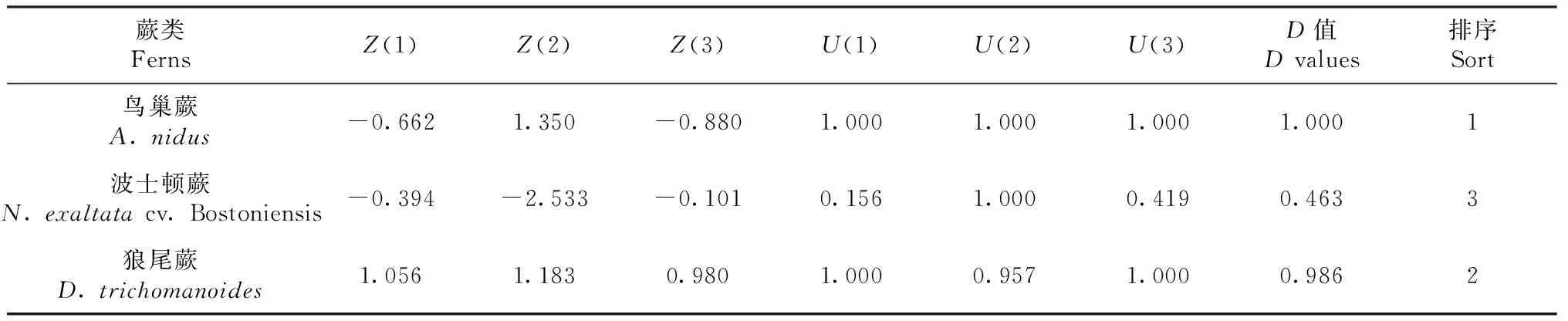
表1 3种蕨类植物综合指标值、隶属函数值、D值
3 讨 论
3.1 低温胁迫对3种蕨类植物叶片光合特性的影响
光合作用是植物重要的生理活动之一,常受到温度的影响。3种蕨类植物光合作用对低温胁迫的响应存在差异。鸟巢蕨在低温胁迫(0 ℃)后叶片的叶绿素含量升高,这与张学财等[19]对剑麻低温处理的研究结果相似。波士顿蕨叶片的叶绿素含量较对照有所降低,表明低温胁迫使其叶绿素的积累减少,这与易拓等[20]的研究结果一致。狼尾蕨叶片的叶绿素含量先下降后上升,参考前人研究[21]可知,这是由于狼尾蕨在低温胁迫处理前期光合作用降低及叶绿素合成受制而导致的,从而造成狼尾蕨叶片的叶绿素含量降低,但低温胁迫强度并未完全破坏狼尾蕨叶片的光合能力,所以随着胁迫时间的延长绿素含量又上升,这是狼尾蕨对低温胁迫环境逐步适应的表现。各个光合参数能反映植物的光合作用水平[22]。随着低温胁迫时间的延长,鸟巢蕨、波士顿蕨及狼尾蕨3种蕨类植物叶片的净光合速率(Pn)、蒸腾速率(Tr)、气孔导度(Gs)及水分利用率(WUE)等光合参数总体呈下降趋势,表明低温胁迫对3种蕨类植物的光合作用产生不利影响,导致净光合速率(Pn)下降。同时胞间CO2(Ci)浓度显著上升,表明净光合速率(Pn)的下降是由非气孔因素导致的。恢复常温20 ℃ 48 h后3种蕨类植物叶片的Pn、Tr、Gs及WUE等光合参数均有一定程度的回升,Ci浓度下降,这与邵怡若等[23]对假连翘的研究结果一致,表明3种蕨类植物叶片的光合作用得到了一定程度恢复,且3种蕨类植物均有一定的抗寒能力。3种蕨类植物低温处理间Pn、Tr、Gs、Ci和WUE等光合参数差异显著,应该是由于3种蕨类植物遭到低温胁迫时,光合系统受损程度不同,从而导致各光合参数存在显著差异,这与黄丽芳等[24]对3种咖啡的研究结果一致。
3.2 低温胁迫对3种蕨类植物叶片相对电导率和丙二醛的影响
随低温胁迫时间的延长,3种蕨类植物的相对电导率均呈现升高趋势,相对电导率与蕨类植物耐寒性呈负相关,研究结果与前人研究一致[20]。低温胁迫会使植物的细胞膜遭遇破坏,膜的透性变大,电解质从细胞内往外渗出,发生膜脂过氧化作用,相对电导率变大。因此,在一定程度上相对电导率的变化可以反映细胞膜受损伤的程度[25]。丙二醛为植物细胞膜脂过氧化的产物之一[26]。本研究不同蕨类MDA含量变化趋势存在差异,表明不同蕨类间细胞膜氧化程度存在显著差异。鸟巢蕨叶片的MDA含量呈上升趋势,表明低温胁迫下鸟巢蕨体内活性氧的产生与清除之间特有的动态平衡被打破,产生膜脂过氧化作用,造成MDA含量增加,这与苜蓿上的研究结果一致[27]。低温胁迫下MDA含量并不总是增加,低温胁迫下波士顿蕨和狼尾蕨叶片MDA含量呈下降趋势,这可能是低温胁迫导致保护物质迅速增加,从而降低植物体内MDA含量,在藜麦幼苗叶片和苜蓿根中都存在此现象[28-29]。
3.3 低温胁迫对3种蕨类植物叶片可溶性糖和可溶性蛋白的影响
可溶性糖(SS)和可溶性蛋白(SP)均为植物体内关键的渗透调节物质,二者具有增强细胞保水能力及清除过多活性氧等功能。低温胁迫时,植物对低温胁迫环境作出响应,SS和SP含量升高,以维持低温胁迫时细胞内的渗透平衡[30-31]。本研究中3种蕨类植物叶片的SS和SP含量均有不同程度的上升,且随着胁迫时间的延长而不断升高,这可能是3种蕨类通过积累SS和SP以维持细胞内的渗透平衡,抵御低温胁迫环境。恢复常温20 ℃ 48 h时SS和SP含量较低温胁迫96 h显著降低,表明3种蕨类植物正常生理及代谢活动得到一定恢复,这与邓仁菊等[32]对火龙果低温胁迫研究结果一致。
3.4 低温胁迫对3种蕨类植物叶片超氧化物歧化酶(SOD)、过氧化物酶(POD)的影响
超氧化物歧化酶(SOD)、过氧化物酶(POD)是植物细胞内2种主要抗氧化酶。该研究中,随低温胁迫时间的延长,3种蕨类植物SOD和POD活性均有不同程度上升,不同蕨类间存在显著差异。低温胁迫下,SOD和POD 2种酶活性均上升,使植物具备抵御低温逆境的功能。这也表明在低温胁迫下,3种蕨类植物可通过增加SOD和POD活性,从而具备清除细胞内过多活性氧的能力,维护细胞内活性氧的代谢平衡,提高其抗寒性,以减轻低温给植物带来的伤害。马晓华等[6]研究发现,2种蝴蝶兰均能通过增强SOD和 POD 这2种酶活性以达到维持细胞内环境的稳定,与本研究结果一致。
4 结 论
通过对3种蕨类植物的生理生化指标及光合参数的测定,可知3种蕨类植物均能对低温作出响应,能忍受一定的低温胁迫强度,均有一定抗寒能力。对鸟巢蕨、波士顿蕨和狼尾蕨进行综合评价时采用隶属函数法,得出3种蕨类低温胁迫下耐受性强弱顺序为鸟巢蕨>狼尾蕨>波士顿蕨,鸟巢蕨较狼尾蕨和波士顿蕨具有更强的抗寒能力。该研究较为系统地探究3种蕨类植物对低温的生理响应机制,可为将来蕨类植物抗寒品种的培育和引种栽培等工作提供理论依据。本研究缺少从分子方面探究其抗寒机理,今后可从转录组学和代谢组学等方面对抗寒机制进行更深层次的研究。
猜你喜欢
哈尔滨轴承(2021年1期)2021-07-21
保鲜与加工(2021年1期)2021-02-06
矿产综合利用(2020年1期)2020-07-24
华人时刊(2020年23期)2020-04-13
少年漫画(艺术创想)(2020年9期)2020-03-19
NBA特刊(2018年17期)2018-11-24
中国民族医药杂志(2016年2期)2016-05-14
中国民族医药杂志(2016年2期)2016-05-14
中国民族医药杂志(2016年3期)2016-05-09
中国民族医药杂志(2016年3期)2016-05-09
