
新型唾液酸糖基化探针Ac4ManNAl代谢标记研究
2022-03-09 02:10豆彪杨如梦郑路李霞王佳佳
中国医药生物技术 2022年1期
豆彪,杨如梦,郑路,李霞,王佳佳
哺乳动物细胞中大多数蛋白可发生糖基化修饰,糖基化作为一种重要的翻译后修饰类型,在信号转导、表观遗传、应激反应、蛋白质降解及细胞凋亡等生命进程中扮演着非常重要的角色,如细胞之间的识别、信号的传导、病原体的入侵以及癌症发生发展等。在脊椎动物中,每个细胞的表面几乎都分布着大量的聚糖分子[1],它们是由 9 种类型单糖组合成的生物聚合物,这些糖链的末端一般为唾液酸。唾液酸是一类带有负电荷且结构相似的九碳糖的总称,通常出现在 N-连接和 O-连接糖基化蛋白结构的末端。哺乳动物体内存在三种唾液酸,其中,N-乙酰神经氨酸(N-acetylneuraminic acid,Neu5Ac)和 N-羟乙酰神经氨酸(N-glycolylneuraminic acid,Neu5Gc)是最为常见的,脱氨神经氨酸(ketodeoxynonulosonic acid,KDN)[2]含量较少。细胞表面唾液酸在不同的生理和病理过程中起着重要作用,例如,细胞表面唾液酸聚糖可作为唾液酸识别蛋白的配体,介导淋巴细胞归巢、免疫反应、细胞之间的识别和病原体入侵等,唾液酸广泛地参与受精[3]、骨骼发育[4]、神经连接[5]等过程,而唾液酸合成和修饰的失调与癌症[6]、神经系统疾病[7]等严重疾病的发生与发展密切相关。
尽管唾液酸聚糖参与了许多重要的生物过程,但是糖基化过程不直接受中心法则调控,且聚糖结构复杂多变,一直缺乏有效的标记技术。前期唾液聚糖识别凝集素或抗体被用于唾液聚糖成像和糖蛋白组分析,然而,基于凝集素和抗体的方法具有低亲和力、毒性和低组织渗透性等,使得其标记方法局限[8]。近些年,化学生物学蓬勃发展,出现了对唾液酸化聚糖以及其他多种糖链结构修饰的代谢标记,科学家开发了基于生物正交反应的非天然糖代谢标记策略,并逐渐成为了在细胞和活体水平上研究糖基化修饰的最主要手段之一。N-乙酰甘露糖胺(ManNAc)是合成唾液酸的前体,在 ManNAc的 N-乙酰位引入一些非天然的基团,例如在乙酰基等结构中加长碳链,这些改造后的 ManNAc 类似物依然能顺利进入唾液酸的代谢途径,从而表达于细胞表面[9]。随后,该团队研究了其类似物 N-丙酰甘露糖胺(N-propanoylmannosamine,ManNProp,图1A)在大鼠中的代谢情况,ManNProp 在多个器官均可被代谢为 SiaNProp,随后被整合到细胞膜表面聚糖中;基于以上研究,Bertozzi 团队[10]开发了具有功能性的 ManNAc 类似物 N-4-氧代戊酰甘露糖胺(N-levulinoylmannosamine,ManLev,图1B)。同样的原理,ManLev 可以通过代谢进而转化为 SiaLev,随后整合到细胞膜上的唾液酸聚糖中,为细胞表面聚糖引入功能基团——羰基,实现了细胞膜表面聚糖可发生化学反应性的功能化。由于羰基和肼的偶联反应需要在弱酸性条件下进行,并且细胞中天然存在含有羰基的分子,导致其应用具有很大的局限性,因而需要设计和开发适用于活细胞中特异性高的偶联反应。随后,该团队于2000年又报道了含有叠氮的 ManNAc 类似物 N-叠氮乙酰甘露糖胺(N-azidoacetylmannosamine,ManNAz,图1C),该探针可通过代谢进入到细胞的唾液酸合成途径,在细胞膜表面糖链中引入含有叠氮基团的唾液酸 SiaNAz,接着利用叠氮与三苯基磷的偶联反应进行功能化修饰,这种方法被称为非天然糖代谢标记(metabolic glycan labeling,MGL)或糖代谢工程(metabolic oligosaccharide engineering,MOE)[11]。2004年 Bertozzi 课题组还将 Ac4ManNAz 采用连续腹腔注射方式成功对小鼠活体组织唾液酸标记[12]。唾液酸的代谢标记即基于上述原理,在唾液酸分子及其合成前体ManNAc 上引入生物正交基团[13],利用生物正交反应[14]或其他手段标记非天然糖[15-18],并通过荧光探针、化学发光等手段,检测细胞表面的糖链结构。
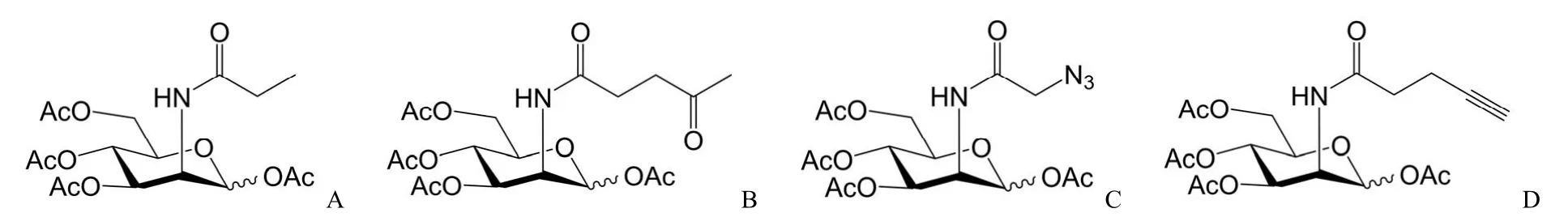
图1 唾液酸代谢标记探针(A:N-丙酰甘露糖胺;B:N-4-氧代戊酰甘露糖胺;C:N-叠氮乙酰甘露糖胺;D:N-丁炔酰基-全乙酰化甘露糖胺)Figure 1 Metabolic probes for sialic acid glycans labeling (A: ManNProp; B: ManLev; C: ManNAz; D: Ac4ManNAl)
随着生物正交反应的大力发展,唾液酸聚糖的代谢标记可选择多种新的生物正交基团,比如上文提到的 Ac4ManNAz,由于叠氮基团体积小、正交反应性好、化学性质较稳定以及便于引入等优点,从而被广泛使用。作为一价铜离子催化的叠氮-炔基环加成反应[18-20](Cu-catalyzed azide-alkyne cycloaddition,CuAAC)中叠氮基团的互补反应伙伴炔基也同样具有作为生物正交化学报告基团的良好特性。炔基具有较好的稳定性,且不天然存在于细胞中,便于特异性偶联。我们课题组前期报道了一系列含有叠氮基团的分子探针[21-24],然而含有炔基基团的探针却鲜有报道,借此本文探讨了能与叠氮正交反应的炔基探针 Ac4ManNAl(图1D),并对其进行活细胞和活体标记分析。
1 材料与方法
1.1 实验材料
人肺腺癌细胞系 A549、H1299、人肾上皮细胞系 293T、人宫颈癌细胞系 HeLa 细胞和小鼠肺癌细胞系 LLC-LUC 细胞、人结肠腺癌细胞系SW1116 均购于 ATCC;C57BL/6J 小鼠购自北京维通利华实验动物技术有限公司。
1.2 方法
1.2.1 细胞培养与传代 所有细胞系在 37 ℃、5% CO2的恒温培养箱中培养。当细胞生长至 90%时即可进行细胞传代。弃去培养基用 PBS 先洗一遍,加入胰酶消化至细胞收缩变圆,然后加入完全培养基终止消化。用移液枪轻轻将细胞吹下,转移至 15 ml 离心管,800 r/min 离心 5 min。弃去上清,用完全培养基重悬细胞,平均传至新的培养皿中,一般 2 天左右传代 1 次。
1.2.2 活细胞代谢标记 六孔板密度为 80% 的细胞用不同浓度的非天然糖处理 24 h。通过胰蛋白酶消化获取,用 PBS 洗涤 3 次。细胞在 RIPA 裂解液(1% NP-40,0.5% 脱氧胆酸钠,1% SDS,50 mmol/L Tris-HCl,150 mmol/L NaCl,蛋白酶抑制剂,pH = 7.4)中裂解并超声,12 000 r/min 离心 15 min 并取上清。用 BCA 法进行蛋白定量,随后取 200 μmol/L 蛋白裂解液与 100 μmol/L Azide-PEG4-Biotin 在 CuSO4/THPTA(100 µmol/L CuSO4,200 µmol/L THPTA)和 5 mmol/L 抗坏血酸钠催化下室温振荡 2 h 进行 Click 反应。加入500 μl 甲醇于 -80 ℃ 静置 2 h,沉淀出的蛋白经离心后,再用 500 μl 甲醇清洗 2 遍;晾干后,用80 μl 4% SDS 和 20 μl 5 × loading 调节蛋白浓度为 2 mg/ml。蛋白利用 10% SDS-PAGE 电泳分离,转膜,加入 5% 脱脂奶粉的 TBST(150 mmol/L NaCl,0.1% Tween-20,10 mmol/L Tris-HCl,pH 7.6)室温封闭 2 h 后进行 Streptavidin-HRP 抗体室温孵育 2 h 及 ECL 免疫印迹显色。
1.2.3 小鼠组织体内标记 选取 6 周龄C57BL/6J 雄性小鼠 2 只。将 2 只小鼠随机分为对照组和 Ac4ManNAl 组,对 Ac4ManNAl 组连续7 d 进行腹腔注射 200 μl Ac4ManNAl(300 mg/kg,溶于 70% DMSO 生理盐水),对照组小鼠腹腔注射同体积 DMSO 生理盐水溶液。第 8 天,对两组小鼠进行安乐死处理,取各组织约 20 mg并研磨成蛋白溶液,通过 Click 反应后,进行 Western blot分析 Ac4ManNAl 对各组织中的聚糖标记情况。
2 结果
2.1 Ac4ManNAl 在细胞代谢标记的浓度梯度
以 HeLa 细胞作为研究对象,分别将终浓度为0、25、50、100、200、500 μmol/L 的Ac4ManNAl探针与细胞共培养 24 h。弃去培养基用 PBS 洗一次,加入 200 μl 细胞裂解液收集细胞于 1.5 ml 离心管中,于 4 ℃ 反应 30 min;BCA 进行蛋白浓度定量,取 200 μmol/L 蛋白进行 Click 反应,最后经 Western blot 分析(图2)。结果表明,随着浓度的增加,代谢标记信号随之增强,提示Ac4ManNAl 探针对细胞的标记具有浓度依赖性,而且当浓度达到 100 μmol/L 时就已经具有很强的标记效果。
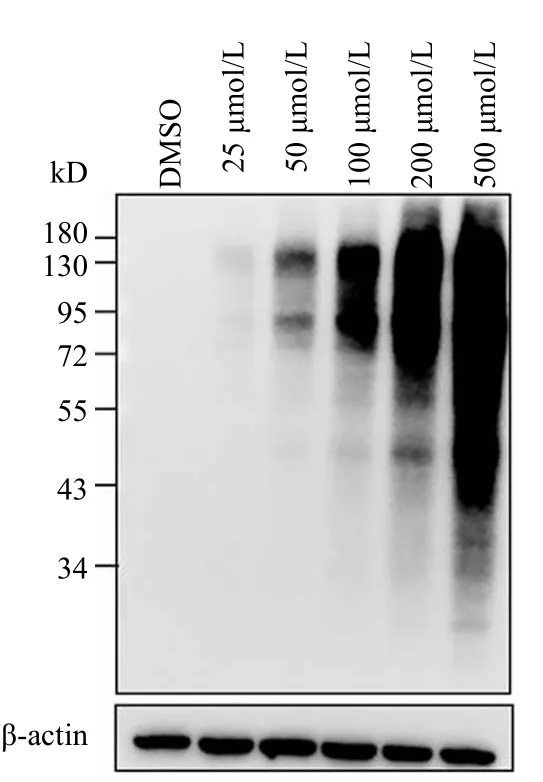
图2 不同浓度 Ac4ManNAl 在 HeLa 细胞中代谢标记效果Figure 2 Various concentrations of Ac4ManNAl metabolic labeling in HeLa cells
2.2 Ac4ManNAl 代谢标记的时间梯度变化
将 200 μmol/L Ac4ManNAl 与 HeLa 细胞共培养 0、6、12、24、36、48 h 六个时间段,收取并裂解细胞,蛋白定量后,进行 Click 反应。经Western blot 分析,如图3 所示,随着孵育时间的增加,代谢标记信号随之增强,且 24 h 时标记信号可以达到峰值,表明 24 h 为此探针的最佳标记时间,延长培养时间并不会有标记效果的提升。
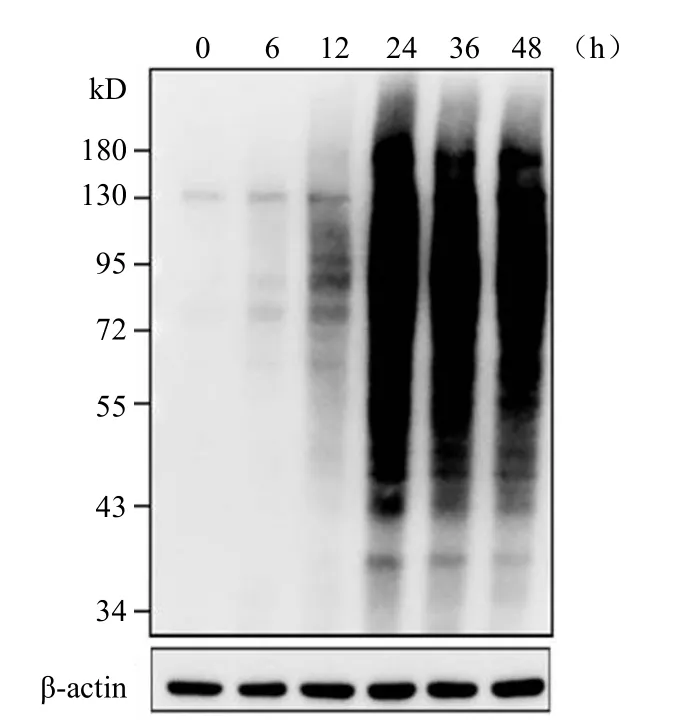
图3 Ac4ManNAl(200 μmol/L)在 HeLa 细胞代谢标记的时间依赖性Figure 3 Ac4ManNAl (200 μmol/L) metabolic labeling in HeLa cells cultured with different time lengths
2.3 Ac4ManNAl 对多种细胞的标记
为了探究 Ac4ManNAl 在不同细胞的标记效率,选取人肺腺癌细胞系 A549、H1299、人肾上皮细胞系 HEK293T、人宫颈癌细胞系 HeLa 细胞和小鼠肺癌细胞系 LLC-LUC 细胞、人结肠腺癌细胞系 SW1116 共六种细胞系与 200 μmol/L 终浓度的 Ac4ManNAl 共培养 24 h。并通过 Western blot分析,评价其在不同细胞系中的标记差异。如图4所示,Ac4ManNAl 对六种细胞均有不同程度的标记效果,证明此探针具有多样性,且根据细胞类型的不同又具有不同程度的标记效果,提示针对不同类型的细胞选取适合的探针进行相关蛋白标记研究。
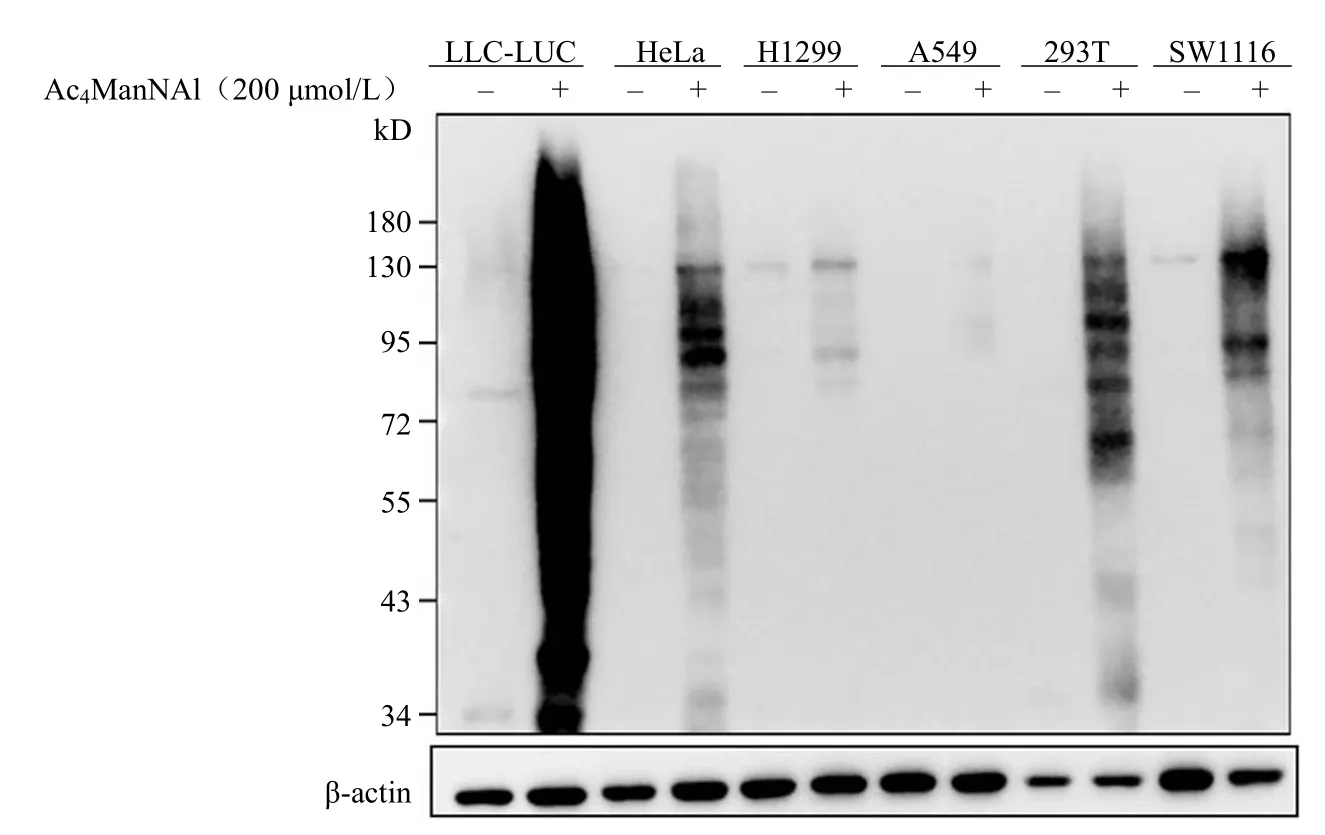
图4 Ac4ManNAl 在六种不同细胞系的代谢标记效果Figure 4 Ac4ManNAl metabolic labeling in a range of mammal cell lines
2.4 Ac4ManNAl 在小鼠体内代谢标记效果
小鼠体内评价了 Ac4ManNAl 代谢标记效果。首先,将小鼠分为对照组和实验组,每组 2 只;实验组连续 7 d 进行腹腔注射 200 μl Ac4ManNAl(300 mg/kg,溶于 70% DMSO 生理盐水),对照组小鼠腹腔注射同体积 DMSO 生理盐水溶液。第 8 天,将两只小鼠进行安乐死处理,随后取其心脏、肝脏、脾脏、肺、肾脏以及直肠组织,经研磨离心获取蛋白,BCA 蛋白定量后,每组取 200 μg蛋白,经 Click 反应后,进行 Western blot 分析,评价 Ac4ManNAl 在小鼠体内对各组织的标记情况。结果如图5 所示,Ac4ManNAl 对六种组织器官均能具有明显的标记效果,证明此探针可以很好地用于体内活体组织标记;而在肝脏和肾脏组织中,标记效果不太明显,推测其原因可能是由于组织中自身存在较高水平的生物素。为了验证这情况,选取肝脏和肾脏两种组织蛋白不经过 Click 反应,直接进行 Western blot 验证,可以观察到肝脏和肾脏本身就具有较高水平生物素的表达,此结果与查询的文献[25]相符。
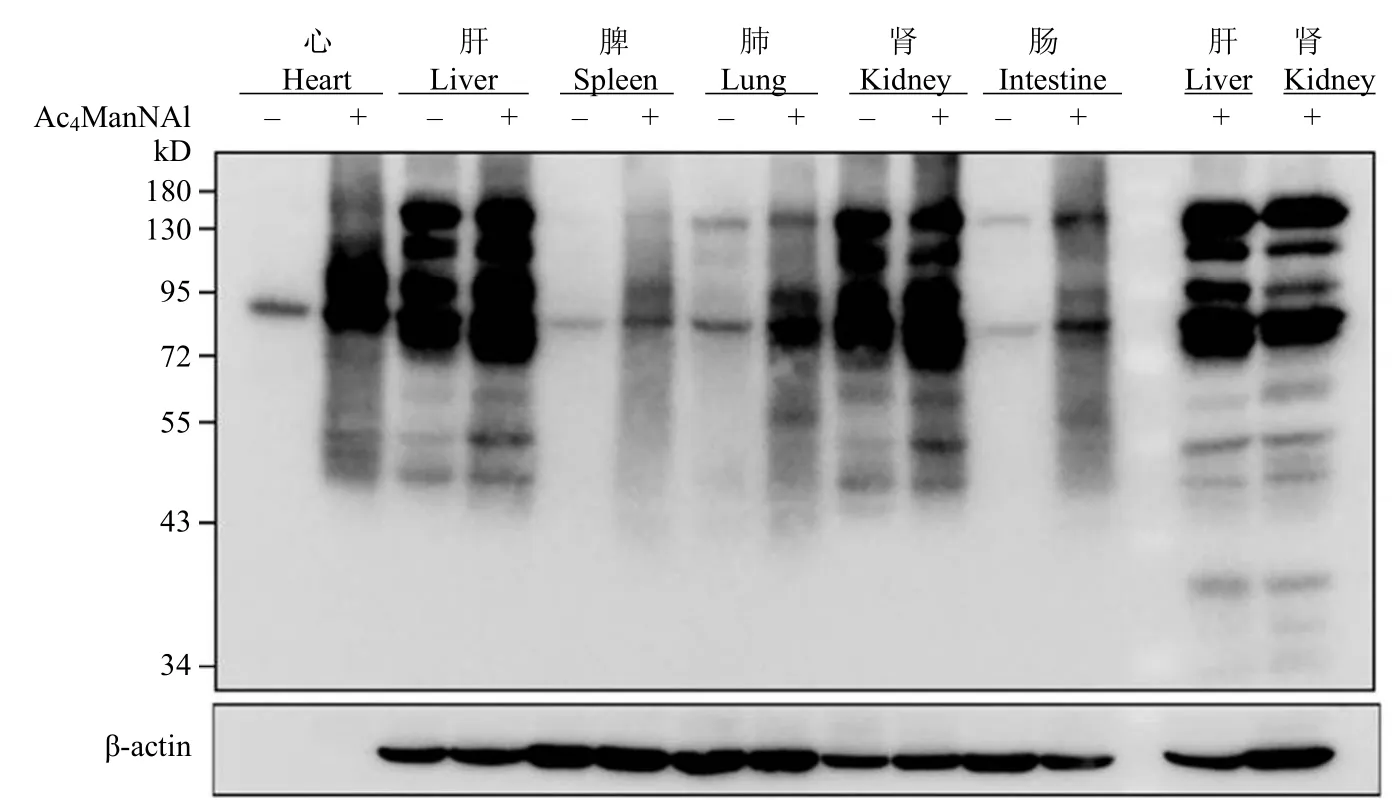
图5 Ac4ManNAl 在小鼠体内的代谢标记Figure 5 Ac4ManNAl metabolic labeling in vivo
3 讨论
核酸、蛋白、糖以及脂类是哺乳动物细胞中的四类生物大分子,其中聚糖广泛存在于已知所有的生物体中,从低等的微生物到高等的人类都存在着各种糖基化修饰。已知在哺乳动物细胞膜表面存在着大量唾液酸,其对细胞的物理、化学性质、生理及病理功能都具有不可缺少的关键调节作用,并且由于唾液酸化聚糖在神经功能、免疫调节功能、肿瘤发生发展等过程中发挥着重要功能,以非天然糖代谢标记策略为代表的化学生物学方法实现了唾液酸聚糖的成像和蛋白组学分析,新的合成策略为获取结构准确的天然糖类化合物进行功能研究开辟了新方向,非天然糖代谢标记技术将在解决相关生物学问题中得到更加广泛的应用。与叠氮基团类似,炔基是一个体积微小的、惰性的生物正交基团,同样可以通过点击化学进行选择性地标记。鉴于过去已经成功地将叠氮化糖类似物加入细胞表面的唾液酸聚糖中,炔基糖也同样可以被这些复杂的天然糖代谢途径接受;另外当用流式细胞术监测细胞结合时,相对于叠氮基团,炔基糖的毒性更小,产生更高的信号,背景更少[26];除此之外,炔基生物正交基团的拉曼信号位于细胞拉曼光谱的“静默区”,因此可以应用拉曼光谱对带有炔基的非天然糖进行成像,而无需进行生物正交反应[27]。
总之,本文运用炔基唾液酸聚糖类似物Ac4ManNAl,可以通过唾液酸代谢途径高效地代谢到细胞表面唾液酸聚糖中,而且我们的工作也证明了其对多种细胞系均有明显的标记效果,体内实验也可以对多种组织器官进行活体标记,对我们后续的肿瘤细胞的唾液酸蛋白的研究以及肿瘤靶向治疗提供一个切实可用的小分子工具。众所周知,唾液酸糖基化蛋白的合成和修饰的失调与多种肿瘤的发生与发展息息相关[6],虽然已报道一些表位,但还有许多其他尚未确认的唾液酸聚糖位点和变化,对这些聚糖的研究,一直以来困难重重。识别这些与糖相关的生物标记物和治疗干预的靶点是现阶段研究的关键目标之一。本文所研究的Ac4ManNAl 为唾液酸代谢标记相关疾病的研究提供了新的研究工具。
猜你喜欢
动物营养学报(2022年9期)2022-10-12
食品科学(2022年18期)2022-10-09
中国畜牧杂志(2022年3期)2022-03-16
吉林大学学报(理学版)(2022年1期)2022-01-21
科学与生活(2021年16期)2021-11-25
青岛大学学报(医学版)(2021年5期)2021-11-17
南方农业学报(2020年11期)2020-02-22
新课程·下旬(2019年7期)2019-09-17
学生导报·东方少年(2019年11期)2019-06-11
发明与创新·中学生(2017年11期)2017-12-07
