
尾巨桉与灰木莲、香梓楠混交对土壤养分及酶活性的影响
2022-03-03 11:03韩小美
广东农业科学 2022年1期
刘 军,毛 纯,韩小美,杨 梅
(1.广西国有七坡林场,广西 南宁 530004;2.广西大学林学院,广西 南宁 530004)
【研究意义】土壤养分状况能综合反映林木生长状况及林地生产力[1]。研究表明,尾巨桉与其他树种混交造林能够显著增加林木种类组成与林地凋落物,有效改善土壤环境[2],同时,合理的混交模式有利于改善林分的光环境,使林分能在时间、空间上最大限度利用光能,提高地上部分光合速率,形成混交树种之间正向互馈效应[3]。【前人研究进展】杨瑞等[4]研究了秦岭山脉典型林分土壤养分,结果表明不同林分土壤养分变化差异较大,土壤性质受到林分种类及海拔等生态环境条件的影响强烈。黄笑等[5]研究了不同类型闽楠人工林土壤养分,结果表明混交林可以改善由于单一树种连栽造成的地力衰退。研究发现,将珍贵树种与桉树混交造林,能够使林地土壤微生物活动旺盛,微生物群落结构和功能显著改善,进而提高土壤有机质含量和土壤酶活性[6]。同时,混交造林能够提高林分冠层结构的空间异质性,直接改善林内光环境,进而间接影响林下植被类型,提高土壤养分及酶活性[7]。【本研究切入点】不同树种混交造林是能够在时间、空间上最大化利用资源的一种高效种植方式,营造科学合理的混合栽培能够使林分充分利用环境资源,改善林分光环境及通风条件,提高生物多样性,促进混交树种协调发展,有利于提高林地生产力和森林资源价值[8]。尾巨桉为桃金娘科树种,生长迅速、抗逆性强、投资收益大,是我国南方地区重要的速丰树种。然而人们长期以来过于看重尾巨桉的经济效益,忽视了纯林连栽及不合理使用化学肥料导致土壤酸化、肥力下降[9]。大量研究发现,营林经营中将尾巨桉与多树种搭配、多模式复合种植,能够显著提高尾巨桉的综合效益,改善土壤地力[10-11]。但目前关于尾巨桉与广西的引种栽培的木兰科珍贵树种混交后土壤质量的研究却鲜有报道。【拟解决的关键问题】本研究以尾巨桉、灰木莲、香梓楠纯林及灰木莲×尾巨桉、香梓楠×尾巨桉混交林为研究对象,比较不同栽培模式下林地土壤养分和土壤酶活性的差异,为营造既能提高林地经济又能提升生态价值的尾巨桉与珍贵树种混交模式提供依据。
1 材料与方法
1.1 研究区概况
研究区位于广西南宁市国有七坡林场七坡分场场直1 林班8、9 小班(107°45′~108°51′E,22°13~23°32′N),属亚热带季风气候,温和湿润,雨季集中在5—9 月份,年均降雨量1 304.2 mm,年平均气温在21.6 ℃左右,平均相对湿度为79.0%,地貌类型以丘陵为主,土壤为典型的赤红壤,土层厚度为70~120 cm。
1.2 试验方法
2014 年6 月,对尾巨桉DH32-29 第一代纯林,株行距为3 m×2 m 按地块进行林分清理和整地,定植坑为40 cm×40 cm×40 cm,每坑施桉树专用肥0.5 kg。2014 年12 月在清理和整地后的林地上设置4 种造林模式,即:种植灰木莲和香梓楠,形成灰木莲、香梓楠纯林;按垂直等高线伐除5列尾巨桉保留10 列萌芽的方式,伐除带上种植灰木莲、香梓楠,与尾巨桉形成带状混交林,株行距均为3 m×2 m(表1)。灰木莲及香梓楠均使用1 年生实生苗造林,伐除带上的尾巨桉根部采用除草剂处理并套袋,使其伐根不产生萌芽;保留带上的尾巨桉则保留1 根萌芽条。定植后,于每年4 月施肥(N∶P∶K=15∶9∶11)1 次,施肥量每株0.5 kg,每年6—7 月除草1 次。于2017年8 月进行试验林调查和土壤样品采集。
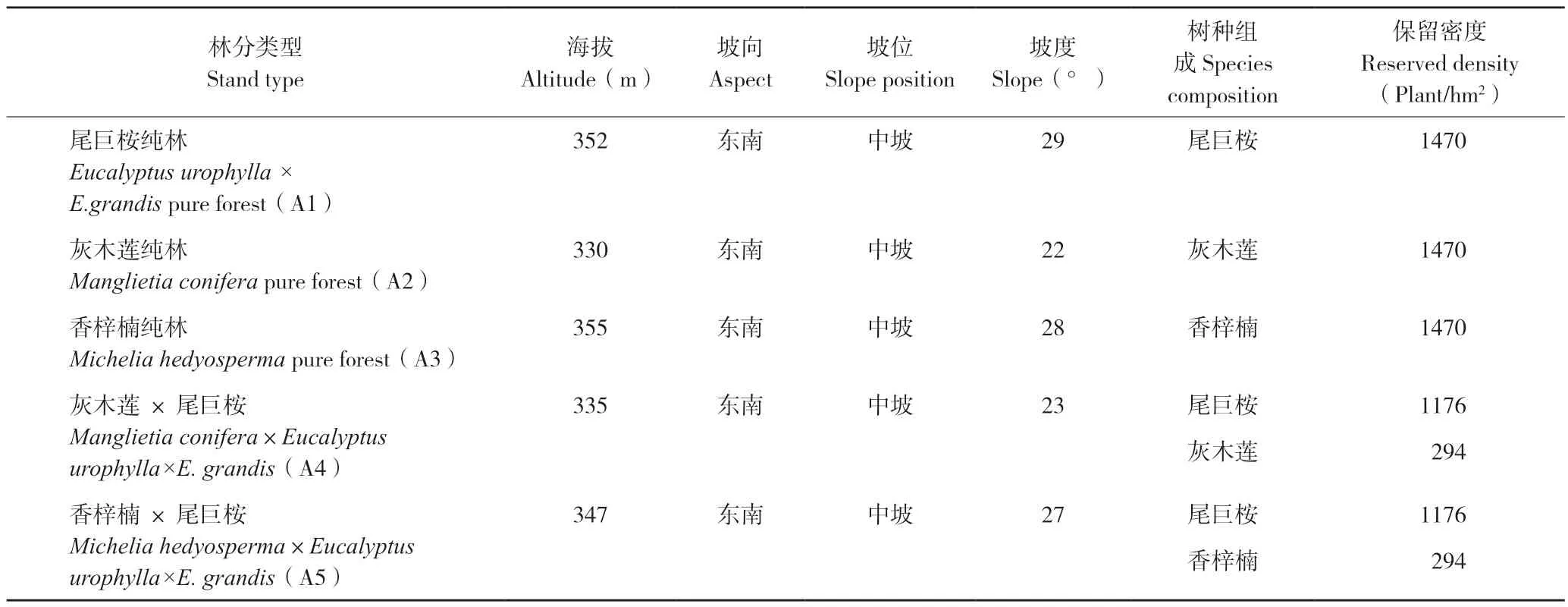
表1 5 种类型林分的样地概况Table 1 Profile of five types of stands
1.3 土壤样品采集及分析
在标准地内采用对角线法布点,挖掘土壤剖面,分别采集0~20、20~40 cm 土壤样品1 kg,装入自封袋,带回实验室自然风干,用于土壤养分及土壤酶活性测定。全氮含量采用浓H2SO4-HClO4消化法-自动凯式定氮仪(KDY-9380,KETUO)测定,全磷含量采用NaOH 碱熔-钼锑抗比色法测定,全钾含量采用NaOH 碱熔-火焰光度法测定,速效氮含量采用NaCl 浸提蒸馏法测定,速效磷含量采用HCl-H2SO4溶液浸提法测定,速效钾含量采用CH3COONH4浸提-火焰光度法测定;土壤过氧化氢酶活性采用容量法(高锰酸钾滴定法)测定,蔗糖酶活性采用3,5-二硝基水杨酸比色法测定,脲酶活性采用靛酚比色法测定,酸性磷酸酶活性采用磷酸苯二钠比色法测定[12]。
采用Excel 2010 对各林分土壤养分及酶活性进行数据处理,用SPSS20.0 对进行单因素方差分析,Duncan 检验进行差异显著性分析。
2 结果与分析
2.1 不同混交模式对林地土壤养分的影响
2.1.1 对土壤全氮、速效氮的影响 如图1 所示,不同混交模式表层土壤的全氮、速效氮含量均高于深层土壤。表层土壤中全氮含量表现为A4>A5>A2>A1>A3,A4 的土壤全氮含量最高为0.50 g/kg,显著高于其他处理,A3 最低为0.35 g/kg;速效氮含量表现为A5>A4>A2>A3>A1,A5 的土壤速效氮含量最高为62.97 mg/kg,A1 最低为36.8 mg/kg。深层土壤中全氮含量表现为A5>A4>A1>A2>A3,A5 的土壤全氮含量最高为0.44 g/kg,分别较A4、A1、A2、A3 高1.13、1.34、1.39、1.51 倍,A3 的土壤全氮含量最低;速效氮含量表现为A4>A2>A5>A3>A1,A4 的土壤速效氮含量最高为36.86 mg/kg,显著高于其他模式,A3 与A5 差异不显著。香梓楠×尾巨桉混交林(A5)不同土层的全氮、速效氮含量均高于香梓楠(A3)、尾巨桉(A1)纯林,灰木莲×尾巨桉混交林(A4)不同土层的全氮、速效氮含量亦均高于灰木莲(A2)、尾巨桉(A1)纯林,且差异均显著,说明尾巨桉混交林具有更高的生物归还量,进而提高土壤氮素的含量。

图1 不同造林模式对土壤全氮、速效氮的影响Fig.1 Effects of different afforestation models on soil total nitrogen and available nitrogen
2.1.2 对土壤全磷、速效磷的影响 如图2 所示,不同造林模式表层土壤的全磷、速效磷含量均高于深层土壤。表层土壤中全磷含量大小顺序为A5>A4>A2>A3>A1,A5 的土壤全磷含量最高为3.79 g/kg,显著高于其他处理,A4 和A2 差异不显著,A3 和A1 差异也不显著;速效磷含量大小顺序为A4>A5>A1>A3>A2,A4 的土壤速效磷含量最高为4.99 mg/kg,A2 最低为1.29 mg/kg,5 种造林模式的土壤速效磷含量差异显著。深层土壤中全磷含量大小顺序为A4>A5>A2>A1>A3,5 种模式差异显著,A4 的土壤全磷含量最高为1.94 g/kg,分别较A5、A2、A1、A3高9.8%、18.6%、46.9%、51.5%,A3 的土壤全磷含量最低;速效磷含量大小顺序为A5>A4>A3>A1>A2,A5 的土壤速效磷含量最高为3.17 mg/kg,显著高于其他模式,A2 模式最低为0.63 mg/kg,A1 和A3 差异不显著。香梓楠×尾巨桉混交林(A5)不同土层的全磷、速效磷含量均高于香梓楠(A3)、尾巨桉(A1)纯林,灰木莲×尾巨桉混交林(A4)不同土层的全磷、速效磷含量亦均高于灰木莲(A2)、尾巨桉(A1)纯林。

图2 不同造林模式对土壤全磷、速效磷的影响Fig.2 Effects of different afforestation models on soil total phosphorus and available phosphorus
2.1.3 对土壤全钾、速效钾的影响 如图3 所示,不同造林模式表层土壤的全钾、速效钾含量均高于深层土壤。表层土壤中全钾含量大小顺序为A4>A5>A2>A3>A1,A4 的土壤全钾含量最高为0.41 g/kg,A1 最低为0.18 g/kg,5 种模式差异显著;速效钾含量大小顺序为A5>A4>A1>A2>A3,A5 和A4 的土壤速效钾含量分别为42.88、40.99 mg/kg,二者差异不显著但显著高于其他模式,A2 和A3 模式土壤速效磷含量较低,二者差异不显著。深层土壤中全钾含量大小顺序为A4>A2>A5>A3>A1,A4 的土壤全钾含量最高为0.29 g/kg,显著高于其他模式,A1 最低为0.16 g/kg,A2、A5 和A3 差异不显著;速效钾含量大小顺序A5>A2>A4>A3>A1,A5 最高为23.35 mg/kg。除深层土壤的A4 速效钾含量小于A2 外,其他均表现为香梓楠×尾巨桉混交林(A5)不同土层的全钾、速效钾含量均高于香梓楠(A3)、尾巨桉(A1)纯林,灰木莲×尾巨桉混交林(A4)不同土层的全钾、速效钾含量均高于灰木莲(A2)、尾巨桉(A1)纯林。

图3 不同造林模式对土壤全钾、速效钾的影响Fig.3 Effects of different afforestation models on soil total potassium and available potassium
2.2 不同造林模式对林地土壤酶活性的影响
如表2 所示,同一造林模式,4 种土壤酶活性均表现为表层(0~20 cm)高于深层(20~40 cm)土壤。表层土壤过氧化氢酶活性以A4 最高、为0.82 U/g·24h,其次为A2、0.76 U/g·24h,二者差异不显著,A3 最低为0.56 U/g·24h;土壤蔗糖酶活性以A5 最高、为60.14 U/g·24h,A2最低为21.06 U/g·24h,5 种造林模式差异显著;土壤脲酶活性以A4 最高、为3.5 U/g·24h,显著高于其他造林模式,A2 最低为1.36 U/g·24h,5 种造林模式差异显著;土壤酸性磷酸酶活性以A5 最高、为1.50 U/g·24h,5 种造林模式差异显著。深层土壤过氧化氢酶活性以A4 最高、为0.74U/g·24h,A1 和A5 差异不显著,A2 和A3 差异亦不显著;蔗糖酶、脲酶和酸性磷酸酶活性均以A5 最高,分别为38.02、2.05、0.95U/g·24h。香梓楠×尾巨桉混交林(A5)不同土层的4 种土壤酶活性均高于香梓楠纯林(A3)和尾巨桉纯林(A1),灰木莲×尾巨桉混交林(A4)4 种土壤酶活性均高于灰木莲纯林(A2)和尾巨桉纯林(A1),总体来看,混交林模式比纯林经营有利于改善土壤酶活性。

表2 不同造林模式土壤酶活性状况Table 2 Soil enzyme activities in different afforestation models
3 讨论
土壤是林木生态过程的载体,是林木生存的物质基础。土壤养分状况直接影响林木的生长代谢[13-14]。不同树种混交必然会导致该林分生态系统发生变化,直接影响林地土壤的养分流动和物质循环,土壤养分含量也必然发生改变[15-16]。本研究中,不同混交模式表层土壤的养分含量均高于深层土壤,林木的凋落物主要分布在土壤表层,0~20 cm 土壤的腐殖质含量必然大于20~40 cm 土壤,导致土壤表层微生物比深层微生物活动更旺盛[17]。尾巨桉是阳性树种且具有速生性,灰木莲和香梓楠在生长初期偏阴性,使得尾巨桉与3 年半生灰木莲和香梓楠混交时长期处于林冠层上方,同时造林密度明显下降,林木间竞争受到抑制,能够充分利用侧方光能和地力,有利于资源获取,因而比纯林长势好;灰木莲×尾巨桉、香梓楠×尾巨桉混交林的土壤养分及土壤酶活性均高于尾巨桉、灰木莲、香梓楠纯林,混交林半分解层凋落物存量均大于未分解层,而纯林相反,说明混交模式有利于林地凋落物积累并能提高其分解速率,增加养分的归还量,提高养分有效性,加快土壤养分矿化,纯林因其化感作用会抑制土壤微生物的分解活动[18]。尾巨桉纯林单一经营模式容易产生次生代谢产物酚酸类物质,进而造成土壤酸化及土壤微生物活动减弱,与其他树种混交有利于抑制土壤酸化[19]。Forrester 等[20]研究发现,混交林分土壤凋落物中氮素含量、氮素同化量及其转化效率均高于纯林,进而导致混交林分土壤中速效氮、磷含量多于纯林。一般认为枯落物分解是林地养分回归的最主要途径,混交林相比纯林,凋落物种类及营养物质更为丰富,在淋溶作用下养分分解释放更快[21]。高空间异质性使灰木莲、香梓楠与尾巨桉混交林的凋落物空间分布更为合理,所营造的土壤微环境更有利于微生物代谢活动,碳源底物能够被分解者更加高效利用[22]。土壤全磷含量在0.8~1.0 g/kg 及以下时,通常被认为林地磷素供应不足。我国南方土壤的磷元素极易被吸附固定成难溶的磷酸盐,表现总体供磷不足[23-24]。大量研究发现,林木混交能够增加根际周围的土壤有效养分含量,混交树种间相互作用在一定程度上能够促进根系分泌物将土壤钾元素和难溶性的磷酸盐活化成易被林木同化吸收的速效钾、速效磷[25]。营造混交林有较少林木和土壤水分的淋洗,有利于土壤速效养分富集表层土壤[26]。
土壤酶活性和土壤微生物是土壤生物化学过程的主要参与者,是土壤生物催化剂[27-28]。本研究中,不同混交模式表层土壤的4 种土壤酶活性均高于深层土壤,主要是因为混交林土壤表层凋落物、微生物较多,土壤微生物和多种酶生理生化反应剧烈,而深层土壤凋落物、微生物较少,生理生化反应相对稳定[29]。香梓楠×尾巨桉混交林4 种土壤酶活性均高于香梓楠、尾巨桉纯林,灰木莲×尾巨桉混交林4 种土壤酶活性均高于灰木莲、尾巨桉纯林,其原因是灰木莲和香梓楠的叶片较大,凋落物所含有机质含量丰富,土壤微生物数量显著增多,混交树种地下部分的互作作用能够促进根系分泌物的分泌,为土壤微生物提供充足的营养,促进微生物的代谢活动,同时根系代谢活动能够释放大量酶类物质,这些酶类物质提高了土壤酶活性[4]。灰木莲、香梓楠与尾巨桉混交改变了林分的生态系统,有效调整了土壤微生物群落结构和功能,在林木-土壤-微生物3 者共同作用下显著提高土壤有机碳含量和土壤酶活性[30]。酸性磷酸酶的主要作用是将有机磷化合物转化为无机磷,其活性高低直接影响林木和土壤中磷素的储量[31]。本研究中,混交林和尾巨桉纯林的速效磷含量相差不大,可能是因为混交林凋落物和微生物数量较多,提高了磷酸酶的活性,促进林分土壤中难溶性磷分解,有利于林木对速效磷的吸收利用,进而导致混交林速效磷含量偏低[32]。
4 结论
本研究结果表明,不同种植模式对林分土壤养分及土壤酶活性影响显著,表层土壤(0~20 cm)的养分含量及土壤酶活性均高于深层土壤(20~40cm),混交林分的土壤养分、土壤酶活性与纯林差异显著,总体表现为:香梓楠×尾巨桉混交林不同土层的养分含量及酶活性均高于香梓楠、尾巨桉纯林,灰木莲×尾巨桉混交林不同土层的养分含量及酶活性均高于灰木莲、尾巨桉纯林。混交林有利于改善林地小气候,改善林木生长环境,增加营养物质储备及提高养分循环速度,使林地土壤地力得到维持和改良。从维持林分生产力和木材产量效益考虑,营造香梓楠×尾巨桉、灰木莲×尾巨桉混交林在改善林地的立地条件、提高林地生产力等方面具有较大潜力。南方的木兰科用材树种多生长较快,木材用途广,可进一步扩大树种筛选,作为桉树人工木林分结构调整的混交树种,促进桉树人工林的可持续发展。
猜你喜欢
小天使·二年级语数英综合(2021年11期)2021-11-21
养生阅刊(2021年5期)2021-06-03
安徽农学通报(2020年13期)2020-07-24
海峡姐妹(2019年1期)2019-03-23
小学阅读指南·低年级版(2018年2期)2018-05-08
小猕猴学习画刊(2017年8期)2017-08-08
中老年健康(2016年12期)2017-01-18
家庭医药·快乐养生(2015年8期)2015-09-10
