
金塔柏扦插不定根形成与内源激素的调控研究
2022-03-02 06:51刘国彬郭丽琴赵今哲姚砚武
植物研究 2022年2期
刘国彬 廖 婷 王 烨 郭丽琴 赵今哲 姚砚武 曹 均
(北京市林业果树科学研究院,北京 100093)
植物激素是植物体通过自身代谢产生的极其微量的、能够产生明显生理效应的信号分子,对植物的生长发育起至关重要的作用,它可以在合成部位起作用,也可以通过运输在其他组织中发挥作用。内源激素参与调控植物根系发育的各个方面,包括愈伤组织形成、器官发生、根的伸长、侧根的发生等,是控制不定根再生的重要因素。生长素是不定根再生的核心激素,阻断生长素合成、极性运输或信号转导则会阻断整个再生过程。在杜鹃属()扦插生根过程中,生长素作用最为明显,较高浓度的生长素(Indole-3 acetic acid,IAA)利于根原基分化形成和生根。茉莉酸(Jasmonic,JA)能够与组蛋白甲基化修饰协同开启伤口信号转导以促进离体器官生长素合成,进而促进不定根再生。脱落酸(Ab‐scisic acid,ABA)可以调节插穗内水解酶和养分的动态变化促进根原基形成。麻文俊等对日本落叶松()扦插生根期内源激素动态变化的研究表明,随着扦插时间的延长,插穗内玉米素(Zeatin,ZT)含量呈下降趋势,在扦插后30和60 d较易生根无性系的内源ZT含量低于难生根无性系,这个两个时期正是不定根发生期和生根高峰期,表明ZT在日本落叶松不定根再生过程中起重要作用。
金塔柏(‘Beverleyensis’)属 于 柏 科(Cupressaceae)侧 柏 属()侧柏()的一个变种,因其叶色在春秋呈金黄色而极具观赏价值,是常绿针叶树种中的一个独特彩叶类型。我国特有的柏科彩叶种质资源极为稀缺,收集、保存、培育易繁殖、生态抗逆效果好的彩色针叶种质资源对于丰富我国园林绿化树种多样性具有重要意义。自1980年以来,本课题组即开始柏科植物良种选育工作,挖掘出了大量性状优异、具有园林绿化价值的优良类型,并于上世纪末收集、繁育多个金塔柏实生群体,在连续20余年的观测和示范中,选育出了多个抗寒抗旱、叶色稳定的金塔柏优株。目前,迫切需要进一步开展无性系育种工作,以促进彩色针叶种质资源的推广应用。
组培、嫁接和扦插是无性繁殖的主要技术,是植物细胞全能性和再生机制的应用与体现。但侧柏组培体系不健全、嫁接繁殖系数低,而扦插是规模化快繁较优的技术,通过扦插获得的单株能够稳定遗传母本优良遗传特性,是目前开展植物无性系育种的主要技术。人工诱导侧柏扦插生根技术及生理生化机制的研究较少,但也取得了一定进展,如梁荣纳和沈熙环分析了不同侧柏种源扦插繁殖时在生根特性上的地理变异规律,赵今哲等研究了生长调节剂对金塔柏插穗扦插的影响并观察了不定根发育过程中插穗外部形态及组织结构变化,杜常健等比较分析了侧柏古树实生树和嫁接树的插穗中内源激素、非结构碳水化合物、木质化程度和形态解剖学的区别,魏黔春等研究了不同类型植物生长调节剂、不同浓度处理对侧柏古树扦插生根的影响,但均缺乏对不定根形成过程及内源激素动态变化的系统研究。
本文在课题组前期研究基础上,通过连续组织切片技术系统研究不定根的发生模式和组织学起源,通过分析插穗内源激素的动态变化并探讨金塔柏不定根发生模式和不同内源激素对金塔柏扦插生根的影响,旨在为彩色针叶植物扦插不定根发生发育机制研究提供参考。
1 材料与方法
1.1 试验材料
试验材料为20 年生金塔柏和‘蝶叶’侧柏(‘Dieye’)中上部的一年生半木质化枝条,分别于2017年7月上旬、2018年6月下旬采集于北京市林业果树科学研究院彩色针叶树种采穗圃。生根促进剂为自配试剂,有效成分为400 mg·L吲哚丁酸(Indole-3 butyric acid,IBA)。
1.2 扦插方法
6 月底7 月初从树体中上部剪取一年生半木质化枝条,修剪成长12~15 cm、粗2.5~3.5 mm 的插穗,每根插穗上部留1/2 针叶带叶扦插。将修剪好的插穗每30 根一捆,在自配生根促进剂中浸泡插穗基部4 h,然后将处理后的插穗垂直插入日光温室内的扦插床中(基质为河沙,扦插前用0.25%高锰酸钾消毒),扦插深度为插穗长度的1/2,每行30根,株行距为5 cm×5 cm,共计900根,450根/品种,3 次重复。扦插完成后,LK-400 24 h 自动喷雾系统控制插床内温湿度。
1.3 试验方法
1.3.1 组织结构观察
自扦插当天(0 d)开始,每隔5 d采集金塔柏插穗基部茎段,直至起苗,共计20 次,每次采集插穗3~5 根,其中在形态变化的关键时期增加取样量。取样时截取插穗基部2 cm 左右茎段,FAA(Formal‐dehyde-acetic acid-ethanol)固定,改良石蜡切片法制片及显微观察。
1.3.2 内源激素提取
根据课题组前期形态学和组织结构观察及不定 根 发 生 阶 段 的 划 分,采 集IBA 处 理 后1 d(Stage1)、35 d(Stage 2,愈伤组织形成)、50 d(Stage 3,不定根起始)、60 d(Stage 4,不定根表达)、70 d(Stage 5,不定根伸长)的金塔柏和‘蝶叶’侧柏插穗基部或根样品(见图1:A~E),放入液氮中进行冷冻,然后存于-80 ℃超低温冰箱保存,以备提取内源激素。采集插穗基部2 cm 区段的茎段,液氮速冻,然后冻存于-80 ℃超低温冰箱,用于IAA、ABA、ZT、GA和JA的测定。
从超低温冰箱中取出插穗样品,加入500 µL提取液((异丙醇)∶(水)∶(盐酸)=2∶1∶0.002),经研磨机匀浆后于-20 ℃静置20 min,冰浴中超声提取30 min;然后加入1 mL 氯仿,经-20 ℃静置,冰浴中超声,涡旋振荡,13 000 r·min离心后分两次提取,合并提取液并挥干;最后利用100µL甲醇复溶,于-20 ℃冰箱保存备用。
1.3.3 内源激素含量测定
质谱条件:采用UPLC-ESI-MS/MS 法对插穗茎段中5 种植物激素进行定性定量检测。采用电喷雾离子源(ESI),正离子模式检测IAA、ZT,负离子模式检测ABA、GA和JA,检测方式为正负离子多重反应监测(MRM)。质谱分析条件如下:Capillary voltage:2.5 kV(-);Cone voltage:30 V;Collision:15 V;Source Offset voltage:50 V;Desolvation tempera‐ture:500 ℃;Desolvation Gas Flow:800 L·h;Cone Gas Flow:150 L·h;Collision Gas Flow:0.17 mL·min。
计算机网络在当前信息社会中所扮演的角色非常重要,如今,已经被运用到了社会的各个领域,网络信息化也是农机安全监理工作的重要组成部分。像农机监理办证局域网、服务器、程序运行以及数据存储等都需要网络的支持,运用网络信息技术,农机监理工作很好的实现了数据集中管理,提升了农机监理办证业务的效率。除此之外,农机监理部门还要将与监理相关的法律法规、规章制度、培训内容以及事故处理等放在农机平台上。这样,农机使用者就可以直接在平台上下载自己所需的内容。
色谱条件:柱温40 ℃,进样体积2µL;流动相A(0.1%甲酸—水溶液),流动相B(0.1%甲酸—甲醇)。洗脱程序:0~5 min,5% B;5.0~6.6 min,70%B,6.6~9.6 min,98%B,9.6~12.0 min,5%B。
根据植物激素标准品的保留时间和质谱定性离子对样品中的植物激素定性,外标曲线计算植物激素的含量。测定激素峰面积,与标准曲线对比,计算各激素含量。为保证测定激素的稳定性,在实验过程中全程保持低温,在提取及进样过程中采用棕色进样瓶避免光影响。实验设3次重复。
1.4 数据分析
实验所得数据利用Excel 2020软件处理,采用SPSS软件进行数据相关性分析。
2 结果与讨论
2.1 不定根的发生与发育
金塔柏插穗经人工诱导扦插后,不定根主要从愈伤组织和皮部两个部位伸出,形态上属于愈伤生根、皮部生根和混合生根3种生根类型(见图1:F~H)。

图1 金塔柏扦插不定根的发育时期及三种类型A~E 分别是金塔柏插穗经IBA 诱导后1 d(stage 1)、35 d(stage 2)、50 d(stage 3)、60 d(stage 4)、70 d(stage 5);F~H 分别为混合生根、愈伤生根及皮部生根Fig.1 Development stages and rooting types of shoot cuttingsA-E are 1(stage 1),35(stage 2),50(stage 3),60(stage 4),70(stage 5)days after cutting,respectively;F-H are callus and bark rooting,callus rooting and bark rooting
连续组织切片发现,金塔柏插穗组织结构主要由髓、木质部、维管形成层、韧皮部、皮层、表皮等组成(见图2,0 d)。扦插后20 d,皮层薄壁细胞逐渐变大增厚,向外分化形成细胞较大、排列疏松的愈伤组织,外观上表现为插穗基部膨大或在切口部位出现白色愈伤组织;同时,在次生韧皮部和皮层中发现不定根原基(见图2,20 d)。35 d后,愈伤组织进一步增多,其内部的薄壁细胞独立分化形成不定根原基(见图2,35 d)。随着扦插的进程,木射线原始细胞分化产生木质部的各类细胞,在此过程中,有的区域分化缓慢,尤其在初生木质部与次生木质部结合部位,会形成一团由未分化成熟的木质部细胞组成的分生细胞团,具有较强的分生能力,不断向形成层方向反分化形成不定根原基(见图2,45 d)。扦插后50 d,随着愈伤组织的不断增殖,其内部不断独立分化出圆团状的不定根原基(见图2,50 d)。
扦插后60 d,观察到源自髓射线的不定根原基穿过木质部、韧皮部进入皮层,即将突破皮层形成不定根;同时,该时期也发现在初生木质部与次生木质部结合处分化缓慢的分生细胞团在合适条件下不断反分化形成不定根原基(见图2,60 d)。扦插后70 d,在愈伤组织中观察到大量不定根原基(见图2,70 d)。扦插后75 d,不定根原基穿过皮层或愈伤组织伸出体外,形成不定根(见图2,75 d)。不定根原基的形成贯穿不定根发生发育的整个过程,在不定根伸长的过程中,在插穗组织内仍发现不定根原基的形成与发育。在扦插后80~90 d,在皮层、髓射线与形成层交界处、木质部靠近形成层一侧发现发育中的不定根原基,部分根原基成帽子结构向外分化发育(见图2,80 d)。自扦插后70~90 d,进入不定根伸长期,同时伴随着不定根原基的形成与不定根发生(见图2,90 d)。

图2 人工诱导金塔柏扦插生根过程中的组织结构变化Xy.木质部;Vc.维管形成层;Ph.韧皮部;Co.皮层;Ep.表皮;Rp.根原基;Ca.愈伤组织;Pi.髓;Pr.髓射线;Ar.不定根;Rc.根冠;0 d.插穗基本结构(10×);20 d.韧皮部与皮层中产生根原基(20×);35 d.愈伤组织中产生根原基(20×);45 d.木射线细胞分化形成根原基(20×);50 d:愈伤组织内部不断独立分化形成根原基(20×);60 d.来自髓射线的不定根原基进入皮层,同时在木质部伴随不定根原基的产生(20×);70 d.愈伤组织中产生大量不定根原基(10×);75 d.不定根伸出体外(4×);80 d.皮层和髓射线与形成层交叉处不定根原基正在发育(10×);90 d.不定根原基形成与发育,不定根形成同时存在(4×);120 d-A.不定根原基在形成层产生(20×);120 d-B.不定根原基突破皮层(10×);120 d-C.愈伤组织中产生的不定根原基(20×);120 d-D.不定根突破愈伤组织伸出(4×);120 d-E.伸出中的不定根(4×);120 d-F.伸长中的不定根(4×)Fig.2 Tissue structure changes of cuttings during root formationXy.Xylem;Vc.Vascular cambium;Ph.Phloem;Co.Cortex;Ep.Epidermis;Rp.Root primordium;Ca.Callus;Pi.Pith;Pr-Pith ray;Ar.Adventitious root;Rc.Root cap;0 d.The structure of the cutting(10×);20 d.Root primordiums originated from phloem and cortex(20×);35 d.Root primordiums origi‐nated from callu(s20×);45 d.Root primordiums originated from xylem ray cell(s20×);50 d.Root primordiums continue to differentiate and form in callu(s20×);60 d.Root primordiums which originated from pith ray is coming into cortex,with the formation of root primordiums in xylem(20×);70 d.More and more root primordiums formed in callu(s10×);75 d.Adventitious roots stick ou(t4×);80 d.Root primordiums originated from cortex and the junction of pith ray and cortex(10×);90 d.Root primordia and adventitious root formation coexisting in late cutting(4×);120 d-A.Root pri‐mordiums originated from vascular cambium(20×);120 d-B.Root primordiums breaks through the cortex to form adventitious root(s10×);120 d-C.More root primordiums originated from callu(s20×);120 d-D.Root primordiums breaks through the callus to form adventitious root(s4×);120 d-E.The adventitious roots are forming(4×);120 d-F.The adventitious roots start to grow(4×)
2.2 不定根发生过程中内源激素含量的变化
2.2.1 扦插生根特性
研究发现,在相同的IBA 处理下,金塔柏和‘蝶叶’侧柏不定根发生进程较为一致,扦插后35 d 愈伤组织开始形成,50 d 后肉眼可见米粒状突起,60 d 后开始形成不定根,70 d 后不定根开始伸长。120 d 时生根调查显示金塔柏生根率为51.11%,不定根条数为2~9 条,平均4.4 条;根长范围为6.1~13.2 cm,平均8.71 cm(见表1)。‘蝶叶’侧柏生根进程与金塔柏较为一致,调查显示其生根率为33.33%,不定根条数为2~4 条,平均2.8 条,平均根长7.99 cm(见表1)。金塔柏对照(CK)生根进程极为缓慢,扦插后50 d 偶见愈伤组织形成,多数插穗死亡,少量处于休眠状态,120 d 调查发现生根率为3.33%,不定根条数1~3 条,平均2 条,平均根长6.05 cm(见表1)。经IBA 诱导处理的所有指标都优于对照,且不同个体间存在差异。
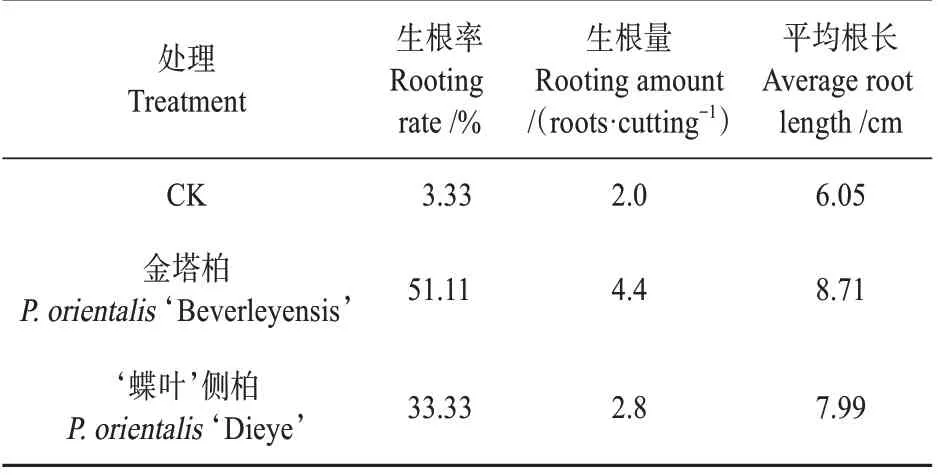
表1 金塔柏生根特性Table 1 Rooting characteristics of P. orientalis‘Bever‐leyensis’
2.2.2 内源激素的动态变化
金塔柏与‘蝶叶’侧柏插穗不定根发生过程中内源激素的动态变化如图3 所示。本实验在插穗中检测到的激素为IAA、ABA、ZT 和JA,GA未检测到。
金塔柏插穗内源IAA含量在不定根形成过程中呈现持续降低趋势,后期略有升高(见图3A)。IBA诱导处理后1 d,内源IAA含量最高(24.5 ng·g),随后急剧下降,从扦插前的24.5 降至4.2 ng·g,降低了82.86%,此时愈伤组织开始形成,在部分插穗中观察到根原基的发生;从诱导期(stage 2)至不定根起始期(stage 3),再至不定根表达(stage 3),内源IAA 含量仍呈下降趋势,但变化趋势平缓,从4.2 ng·g降 至3.0 ng·g,再 降 至2.6 ng·g,降 低 了38.10%;从不定根伸长阶段(stage 5)开始,内源IAA 含量逐渐回升。‘蝶叶’侧柏中IAA 含量在5 个阶段的变化趋势与金塔柏相似,但IAA 水平略低。分析认为在不定根发生过程中,愈伤组织的形成和不定根原基的诱导、不定根的起始与表达过程是消耗IAA 的过程,尤其愈伤组织的形成与根原基的诱导会消耗大量内源IAA,从而导致内源IAA急剧降低;而随着愈伤组织和根原基的形成与发育,插穗进入自我发育阶段,在消耗IAA 的同时也开始自我合成新的IAA,但IAA的消耗与合成相一致,从而使总IAA 维持在平稳水平;而随着不定根进入伸长生长阶段,插穗自我合成IAA 能力得以加强。

图3 金塔柏茎段不同发育期4 种内源激素含量的动态变化(n=3)Stage-1.扦插后1 d;Stage-2.扦插后35 d;Stage-3.扦插后50 d;Stage-4.扦插后60 d;Stage-5.扦插后70 d;下同Fig.3 Dynamic changes of endogenous hormones in cut⁃ting(sn=3)Stage-1. 1 d after cutting;Stage-2. 35 d after cutting;Stage-3. 50 d after cutting;Stage-4. 60 d after cutting;Stage-5. 70 d after cutting;the same as below
金塔柏插穗内源ABA含量呈现先上升后持续降低至平稳的趋势(见图3B)。扦插后,经过35 d的诱导,内源ABA 含量迅速积累,由0 d 的179.1 ng·g快速升高至376.8 ng·g,峰值出现在愈伤组织形成期;之后,随着不定根原基的形成与发育,ABA 含量急剧降低(99.9 ng·g),而随着不定根的表达与伸长,ABA 含量变化趋缓。而‘蝶叶’侧柏ABA 含量整体水平低于金塔柏,但变化趋势相同。分析认为插穗损伤后促进ABA 的快速积累,至愈伤组织形成达到高峰,随着根原基的形成及不定根的发育,ABA 的积累下降与消耗上升并存,从而维持在低水平,表明愈伤组织的形成过程是ABA的积累过程,而低水平的ABA 利于不定根形成与表达。
金塔柏内源ZT 在生根过程中表现为先下降后升高的变化趋势(见图3C)。其中扦插后1 d 插穗内ZT 含量最高(0.026 ng·g),随后急剧下降,在愈伤组织形成期(stage 2)达到最低水平(0.001 ng·g),表明愈伤组织的形成是一个内源ZT 大量消耗的过程;自愈伤组织形成后,根原基开始形成,插穗恢复合成ZT 的能力,但由于扦插后期愈伤形成与不定根发生并存,因此ZT 的合成与消耗同时存在,但合成逐渐高于消耗,因此后期呈现缓慢升高的趋势。‘蝶叶’侧柏内源ZT 含量的变化趋势与金塔柏前期相一致,‘蝶叶’侧柏后期ZT 含量变化趋势平稳,推测其插穗在该阶段形成较多愈伤组织,从而消耗较多ZT。
在整个不定根发生过程中,JA 含量呈现先升高后降低再略有升高的规律性变化(见图3D)。JA含量随着愈伤组织的形成快速积累,与ABA类似,同样在愈伤组织形成期达到峰值(87.8 ng·g),为根原基的形成与不定根的发生奠定了基础。愈伤组织形成后,随着根原基的发育与不定根的形成,JA 含量急剧下降(10.6 ng·g)。不定根原基向外发育突破皮层或愈伤组织形成不定根的过程中,插穗恢复合成JA 的能力,JA 含量开始有所上升。而‘蝶叶’侧柏在整个生根过程中的JA 含量变化与金塔柏相似,不同之处在于其后期JA 含量仍处于下降趋势。分析认为前期JA 的快速积累促进了不定根的形成,后期低水平的JA 含量利于二级侧根的形成。
在扦插生根的5个关键时期,金塔柏和‘蝶叶’侧柏插穗内源激素的比值变化趋势类似,但又有差异。IAA/ABA 值随发育进程呈先降低后升高的趋势,扦插后1 d,金塔柏IAA/ABA 值显著高于‘蝶叶’侧柏,随着愈伤组织的形成,IAA/ABA 值下降;随着根原基的形成与发育,IAA/ABA 值开始缓慢回升,但‘蝶叶’侧柏则略有下降(见图4A)。插穗IAA/ZT 值表现为先升高后降低至平稳的趋势,随着愈伤组织的形成,IAA/ZT值逐渐变大;随着根原基的形成与发育,IAA/ZT值急剧下降直至平稳,不同品种间差异不显著(见图4B)。IAA/JA 值的变化与IAA/ABA 相似,扦插后至愈伤组织形成,IAA/JA值急剧降低,在愈伤组织形成期达最低值,然后比值小幅度上升,但基本维持稳定(见图4C)。
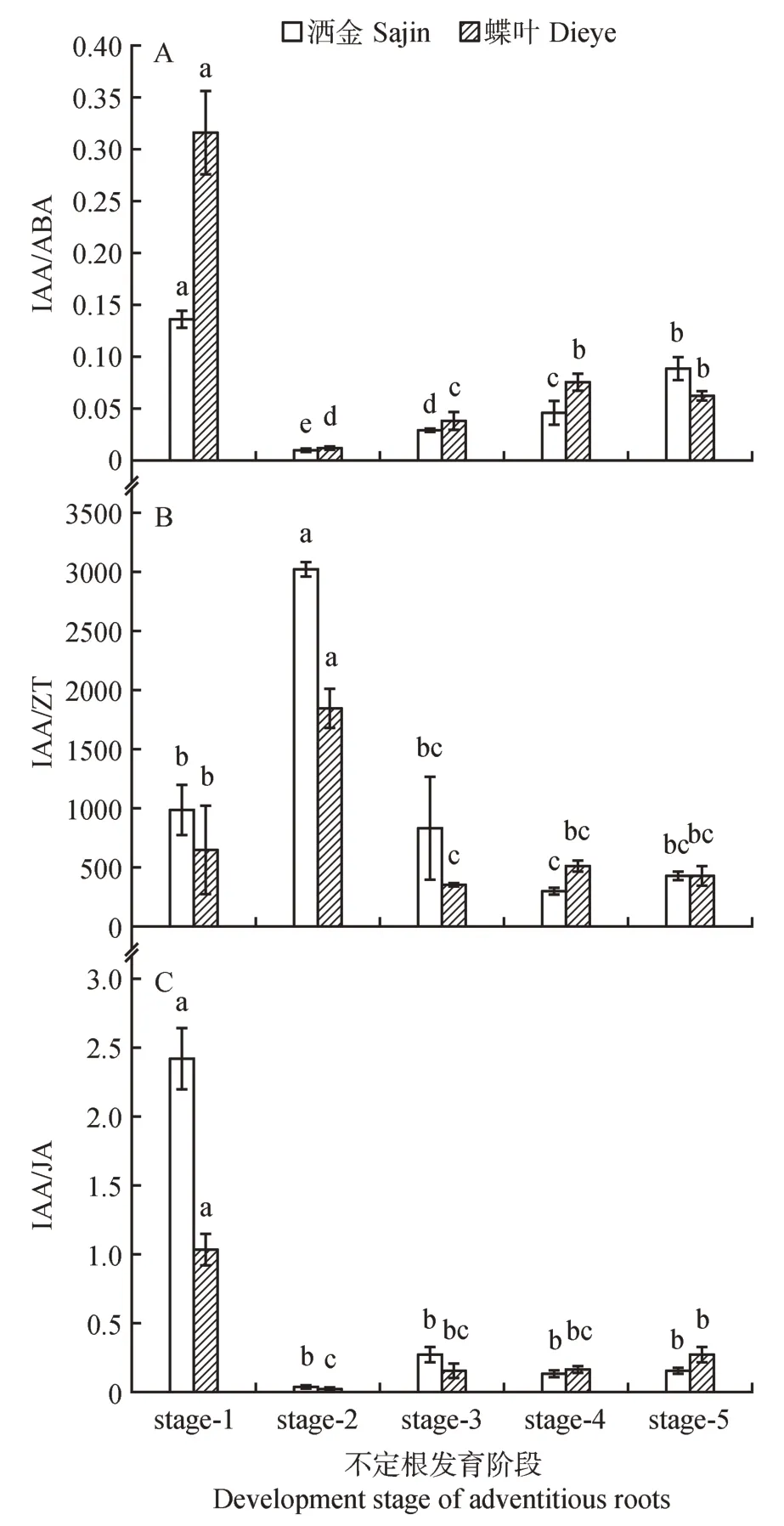
图4 金塔柏茎段不同发育期内源激素间比值及总激素含量的动态变化(n=3)Fig.4 Dynamic changes of endogenous hormone ration and total hormone content in cuttings(n=3)
3 讨论
植物扦插不定根的发生类型从形态学上依据不定根的伸出部位分为愈伤组织生根型、皮部生根型和混合生根型三种类型,依据不定根原基的起源部位,又分为单一位点发生和多位点发生。不定根的发生模式因树种不同而有所不同,有的树种属于单一皮部生根型且不定根原基起源部位单一,不定根根原基只从形成层与髓射线交叉处或木质部和韧皮部之间的维管形成层处发生,如欧美杂种山杨(×)、合欢();有的树种属于皮部生根型,但不定根原基可由初生韧皮部中的薄壁细胞、髓射线与形成层交叉处的部分薄壁细胞发育而来,属于非愈伤组织生根的多位点发生,如欧洲鹅耳枥()。有的树种属于单一位点对的愈伤组织生根型,扦插后先在形成层处形成愈伤组织,再由愈伤组织内分化出根原基,如薄壳山核桃()。有的树种可同时由愈伤组织和皮部诱导产生不定根,不定根原基起源为多位点发生,如桃()的不定根原基在皮层、韧皮部、木质部和愈伤组织中都可以形成,红缨海棠(‘Hongying’)的不定根原基诱导产生于愈伤组织、维管形成层、髓射线与皮层交界处、髓射线与环髓带交界处等部位。有的树种既有皮部生根又有愈伤组织生根,且皮部生根与愈伤组织生根相互抑制,如含笑();有的树种包含愈伤组织生根、皮部生根、愈伤组织和皮部混合生根型,大部分不定根从插穗基部愈伤和皮部的交界处伸出,如欧洲云杉()、圆 柏()、银 杏()。本课题组在前期研究中,认为侧柏扦插生根属于多位点发生模式,并在金塔柏插穗愈伤组织、髓射线与形成层的交界处2个部位观察到根原基形成,本试验通过连续组织切片观察,进一步发现,除了在愈伤组织、髓射线与形成层的交界处发现根原基形成外,不定根原基在髓射线、木质部、维管形成层、次生韧皮部、皮层中都可以形成,而且金塔柏扦插的生根类型包括愈伤组织生根、皮部生根和混合生根三种类型。
植物激素在不定根发生过程中起着重要作用,在解剖学的基础上研究植物激素与扦插生根的关系一直是不定根发生发育生理研究的热点,其中生长素、细胞分裂素、脱落酸、赤霉素、茉莉酸等作为不定根发生发育的诱导因子,与愈伤组织的诱导、根原基的发生、根的发育息息相关。生长素与生根关系密切,多数研究认为生长素含量与生根有直接关系,是调控不定根发生的关键激素,在激素互作调控不定根发生中起主导作用。但不同树种中生长素的调控作用有所不同,IAA 在楸子()扦插生根中只起协同作用,较高含量的IAA 抑制四倍体刺槐()嫩枝扦插生根,而较高浓度的内源IAA 含量利于杜鹃属根原基分化形成和生根。在本试验中,插穗经IBA 处理扦插后1 d 具有较高浓度的IAA 含量积累,随着愈伤组织的形成与根原基的发生发育而逐渐降低,不定根形成后逐步回升,说明愈伤组织与根原基的形成是一个消耗IAA 的过程,而扦插前期较高水平的IAA 积累有利于后期愈伤组织和根原基的诱导分化,后期不定根形成后促进IAA 积累;ABA 含量在扦插后初期较低,随着愈伤组织的形成而逐渐积累,然后在整个生根过程中持续下降至平缓,这有利于插穗内淀粉水解为糖,进而促进根原基的形成和插穗生根。细胞分裂素在不定根诱导的早期过程中发挥作用,在愈伤组织形成之前进行积累并维持在较高水平以调控愈伤组织的分化,本实验中金塔柏和‘蝶叶’侧柏插穗在扦插前期茎段内内源ZT 维持在较高水平,随着愈伤组织的形成而下降,愈伤组织形成后开始缓慢上升,进一步证实细胞分裂素可调控愈伤组织的形成。茉莉酸介导的受伤信号可以促进植物再生,本试验中茉莉酸在扦插前期水平较低,在愈伤组织诱导形成过程中积累达到最高水平,随着根原基的诱导与不定根形成而缓慢升高,该研究结果证实茉莉酸介导愈伤组织发育并促进不定根的发生,后期茉莉酸含量的提高则有助于不定根的形成与伸长,推测这也是‘蝶叶’侧柏生根率低于金塔柏的原因之一。不定根的发生发育是不同激素调控的复杂发育过程,植物激素间的动态平衡决定了不定根的发生,其中生长素、细胞分裂素、脱落酸、茉莉酸之间的平衡决定了再生器官的发育命运,IAA/ABA 值和IAA/JA 值下降、IAA/ZT上升促进愈伤组织形成,IAA/ABA 值上升、IAA/JA值维持在较低水平、IAA/ZT 比值下降促进根原基分化及不定根的发育。
猜你喜欢
新农民(2022年22期)2022-11-07
星星·诗歌原创(2021年3期)2021-06-01
健康博览(2021年3期)2021-03-22
特别健康·下半月(2020年10期)2020-10-20
农民致富之友(2019年27期)2019-10-11
特别文摘(2019年7期)2019-04-13
农民致富之友(2019年3期)2019-03-15
扬子江(2018年1期)2018-01-26
读写算·小学低年级(2017年7期)2017-08-11
农家顾问(2016年6期)2016-05-14
