
胞外聚合物在蓝细菌钙化过程中的作用及其地质意义
2022-03-01 10:15钟怡江文华国陈洪德刘磊陈安清王兴龙王志伟白璇
沉积学报 2022年1期
钟怡江,文华国,陈洪德,刘磊,陈安清,王兴龙,王志伟,白璇
1.油气藏地质及开发工程国家重点实验室(成都理工大学),成都 610059
2.成都理工大学沉积地质研究院,成都 610059
0 引言
蓝细菌的化石记录长达35亿年[1],通过对其矿化特征及机理研究有望找到解锁地球演变之谜的重要“密码”。然而,受成岩和变质等地质过程的长期改造,甄别古老岩石中尺寸微小(介于微米与纳米之间)且不易与其它地质矿物分辨的微生物化石[2]是找到“密码”的关键过程。钙化作用(形成含钙碳酸盐,包括低镁方解石、高镁方解石、文石和白云石)是微生物矿化作用最广泛的方式[3-4],但是直到新元古代之前,钙化的蓝细菌化石都是及其稀少或缺乏的[5-6]。碳酸盐的饱和度一直很高的元古代为什么会如此缺乏钙化的蓝细菌,也是科学家为之不解的“前寒武纪之谜(Precambrian enigma)”[7]。
蓝细菌分泌到细胞外的胞外聚合物(extracellular polymeric substances,EPS)是其新陈代谢过程与外界沟通的必然通道,也是钙化作用发生的主要场所。蓝细菌是微生物席中EPS 的主要生产者[8-10],微生物碳酸盐岩就是钙化微生物席石化作用的产物[11]。蓝细菌的EPS 能够提供成核位置[12-13],影响钙化的过程[14]和产物[15-16]。蓝细菌EPS的钙化作用可发生在其新陈代谢过程中,同时也可与蓝细菌活着与否无关[17-20],甚至发生在蓝细菌死亡后EPS 降解的过程中[21]。大多数报道认为EPS 对蓝细菌的钙化起促进作用,但是在某些环境条件下EPS 也会阻止蓝细菌的钙化[22]。
因此,蓝细菌的EPS 对蓝细菌及微生物席的钙化起到了至关重要的作用[23]。本文选取蓝细菌及其产生的EPS 为主要剖析对象,旨在通过系统介绍蓝细菌及胞外聚合物的主要特征,回顾和总结前人在蓝细菌光合作用有关和降解过程中的钙化过程研究成果,重点剖析蓝细菌胞外聚合物在其钙化过程中的作用,旨在为探索与蓝细菌和微生物席钙化有关的系统科学问题提供借鉴和启示。
1 蓝细菌的主要特征
蓝细菌能像真核藻类一样进行光合作用和固氮作用[24-25],早期一直被认为是一种藻类,称之为“蓝绿藻”(blue-green algae)或者“蓝藻”(blue algae)。后来人们发现其既没有藻类生物所该有的叶绿体(只是能够合成叶绿素,确切的说是叶绿素a),也没有真正的细胞核,只是一种大型的单细胞原核生物(prokaryote),并且是真细菌的一种[26]。由于藻青素(phycobilin)和藻蓝素(phycocyanin)的存在使其呈现出蓝色,因此该种细菌被称之为“蓝细菌”[24,27]。
1.1 形态特征
按形态可将蓝细菌分为5大类群,包括29个属,并且细胞大小差异悬殊,最小的聚球蓝细菌属(Synechococcus)仅为0.5~1 μm,而巨颤蓝细菌属(Oscillatoria)可超过60 μm[28]。通常蓝细菌以单细胞(如球状蓝细菌,coccoid)的单列、多列组合、分枝状(图1)[29]或以席状聚集体的细胞集群形式生存,并且周围都会被其分泌的无结构分散黏液层(unstructured diffuse mucilage)、荚膜(capsule)或者鞘(sheath)所包裹。单列蓝细菌称之为毛状体(trichomes),一个或多个毛状体被鞘(sheath)包裹起来称之为丝状体(filament)(图2)[30]。鞘的存在是蓝细菌的一个分类标准[3]。

图1 球状蓝细菌的形貌特征(据Schopf[29])(a)球状Gloeocapsa sp.蓝细菌及包裹的鞘;(b)球状Entophysalis sp.蓝细菌聚集体Fig.1 Morphological features of coccoid cyanobacteria (after Schopf[29])

图2 丝状蓝细菌Arthrospira fusiformis 的形貌特征(据Sili et al.[30])(a)浸在多糖基质中的两个或多个缠绕活动的毛状体形成卵状蓝细菌;(b)两个单独的毛状体由黏液质的外套膜包裹,并外包着厚层的荚膜;(c)一簇典型的卵状毛状体和释放出来的成单独毛状体;(d)卵状和自由的毛状体浸在由印度墨汁反向染色的浓密黏质多糖中Fig.2 Morphological features of filamentous cyanobacteria Arthrospira fusiformis (after Sili et al.[30])
1.2 主要生存状态和环境
现代蓝细菌的生存空间极其广阔,在水中主要以浮游或底栖的形式存在,包括淡水或咸水,甚至超咸水环境等极端环境,在陆地上也是无处不在,甚至荒漠里也能找到它的踪迹。水中的蓝细菌可分为三种生态类型:1)造席类,岩石、沉积物和淹没植物表面微生物席的主要建造者;2)富营养类,沿海、富营养的河流和湖泊等富营养水体中繁盛;3)微蓝细菌(直径<2 μm),通常发育在开阔海洋和干净水体的湖泊[13]。但是蓝细菌最富集的地方还是浅水环境中的微生物席。微生物席从定义上讲就是真正的生物膜(biofilm)[31],是一种由微生物自身、微生物产生和分泌的胞外聚合物以及它们捕获或者沉淀的外来物质共同组成的席状物[11]。在所有的微生物席中,由蓝细菌主导的微生物席最被关注[11]。保守估计,蓝细菌的全球生物量将达到3×1014g 或者1×1015g[32]。相比其他真核藻类,蓝细菌对于高温、干旱、高盐度、紫外线等具有更高的耐受度[33-34],这正是蓝细菌能够在地质历史长河中存在和繁盛的重要原因。
1.3 地质历史中的蓝细菌及古生态作用
目前认为海洋中最重要的蓝细菌种属之一的Prochlorococcus于1988年便被最早报道[35]。在2.4 Ga之前,蓝细菌主要局限在淡水环境中,之后进入海水环境并呈多样化发展[36]。硅化的蓝细菌在元古宙沉积物中较为常见[37]。前寒武纪叠层石建造过程中,蓝细菌可能起到了重要作用[23,38]。虽然钙化的蓝细菌化石最早存在于1.2 Ga 的中元古代,但是直到542 Ma前寒武纪和寒武纪交界之后的古生代才呈幕式产出,之前一直很少被发现[38]。因此,Riding 将前寒武纪叠层石中缺乏钙化蓝细菌化石的现象称为“前寒武纪之谜(Precambrian enigma)”[7,39-40]。
蓝细菌通过光合作用在整个地球历史的生态演化和持续利用太阳能同化CO2转变为有机质方面发挥了极其重要的作用[13,41]。据估计,现代地球上一半的光合作用主要由蓝细菌完成[42],且其中超过一半的部分主要由两种海相蓝细菌Synechococcus和Prochlorococcus完成[43]。蓝细菌的光合作用是地质历史中地球大气圈氧化的主要原因[3,41,44-45],并积极推动了地球在2.0~2.5 Ga 期间(Great Oxygenation Event, GOE)以及540~850 Ma 期间(Neoproterozoic Oxygenation Event,NOE)的两次巨型氧化作用事件。
2 胞外聚合物(EPS)
2.1 微生物胞外聚合物的主要组成成分
胞外聚合物(extracellular polymeric substances,EPS)是有机高分子多聚化合物(分子量可达8 至大于10 000 kDa),不仅被包括光合自养和异养微生物[46]所合成并分泌到细胞外部,同时也偶尔发现被多毛类、真菌、珊瑚和植物根所产生[47]。EPS 的组成因其来源的微生物、生长媒介(特别是碳源)、生长阶段、温度及其他环境因素的不同而存在差异[48],总的来说,主要由多糖(碳水化合物)、蛋白质、糖醛酸、核酸、多肽类、非碳水化合物酸性基团(例如丙酮酸盐或琥珀酸盐)和腐殖质等组成,甚至包括硫酸盐和磷酸盐等无机化合物和细胞外DNA[49-53]。多糖和蛋白质是其主要成分,占总量的70%~80%[54]。
2.2 蓝细菌胞外聚合物的主要特性
蓝细菌是微生物席中EPS 的主要生产者[8-10],超过100 种(隶属于22 个属)蓝细菌都能产生EPS[54]。蓝细菌的EPS 可分为两种类型,一类是黏附在细胞表面部分,并按照结构形态可分为三种:1)鞘(sheaths),紧密包围在细胞或细胞群体表面薄且稠密的膜状物,具有一定的外形,在光学显微镜下不用染色也能看到(图1a);2)荚膜(capsules),与细胞表面紧密联系的厚层包裹体,黏度较大,轮廓光滑(图2b);3)黏液(slimes),细胞周围分散的黏液物质,不能反映细胞形态(图2d)[55-58]。另一类是可溶解释放到周围环境中的部分[13]。EPS 的不同物理状态是大分子之间的不同连接和相互作用造成的。鞘、荚膜和黏液的残余物为胶状或具有弹性,不容易被降解[59-61]。
蓝细菌的EPS 中带负电的酸性功能团能够吸附大量的单价或二价阳离子,并通过双配位桥的形成而利于提高黏性和保持结构的完整性[52]。上述酸性官能团主要包括羧基(R-COOH)、羟基(R-OH)、氨基(R-NH2)、硫酸基(R-O-SO3H)、磺酸基(R-SO3H)和巯氢基(-SH)[62],其中最重要的是羧基和硫酸基官能团[46]。
许多蓝细菌的EPS 包含与脂类有关的乙酰基、多肽类和像海藻糖和鼠李糖的脱氧糖,导致蓝细菌具有疏水性和乳化特征。大多数蓝细菌的EPS具有包含六种或以上单糖的异质多糖(HEPO)复合结构。正是这些大量的不同单糖产生的不同链接类型,使得EPS 具有多种不同的结构和构造,并呈现出复合的重复单元[13]。
包裹在细胞表面的EPS 由于能够成为细胞与其外的有机质、无机新陈代谢产物、捕食者、抗菌素和其他细菌之间的物理屏障[63],抵抗脱水作用、有机酸、噬菌体溶菌、吞噬作用、盐度、紫外线辐射、氧化压力等影响,并能够增加光照、营养摄入和固氮活动[64-65]。同时,蓝细菌的EPS 也有利于细胞聚集和生物膜积聚黏附在沉积物表面,使得各种新陈代谢作用可以在微空间中共存[65-66],从而促进微生物细胞在不同的沉积水动力环境下的稳定生存[67-69]和保护生物膜结构[13,48]。
2.3 降解过程中的蓝细菌胞外聚合物
蓝细菌一般分布在微生物席顶部,其产生的EPS被下部的异养细菌利用过程中发生生物降解。随着埋藏深度的增加,鞘会在生物降解过程中逐渐变成分散的多糖微结构(图3a~b,图3b左—右)[70],最终重新组织成由肿胀的节点相互连接且相对牢固的三维网状(蜂窝状或肺泡状)结构(图4a,b)[47,71]。这类结构被认为是由EPS中多糖链分子内非共价键结合导致坚硬的棒状单元调解而联合成的富含天冬氨酸、谷氨酸和糖醛酸[72]的大分子单元所致[73]。在含水阶段,三维网状结构呈现次球状,直径为几十纳米到几微米,壁厚度为几纳米到1 μm。随着水分减少而干燥,结构会坍塌破坏。这个结构形成了叠层石有机质和碳酸盐沉积的框架,且一直保存到沉积物中1 m深的地方[47,71]。Défargeet al.[47]列举了现代不同环境叠层石中均发育这类源于EPS 的三维网状有机结构,认为这类结构不仅是底栖微生物群体有机沉积,而且是地球表面所有生命有关沉积的重要部分。

图3 Kopare 叠层石中EPS 降解特征的TEM 微观照片(据Decho[70])超薄片进行了PA-TCH-SP(Thiery反应)处理,使得不溶多糖染色变暗;(a)蓝细菌折叠的丝状体和其多糖鞘,箭头指示枯萎细胞的细胞壁脱离了外部鞘,下部的丝状体显示了刚好切过多糖鞘的微纤维结构特征;(b)部分降解的蓝细菌鞘(左边是横切面)和荚膜(右边),可见多糖微结构网状物Fig.3 TEM micrographs of stromatolitic layers of Kopare (after Decho[70])

图4 沉积物中有机三维网状结构的冷冻电镜(Sryo-SEM)照片(据Défarge et al.[47])(a)Tuamoto群岛Hao环礁咸水湖中叠层石顶层;(b)Tuamoto群岛Moruroa环礁Brackish湖中叠层石层顶层Fig.4 Cryo-SEM micrographs of three-dimensional organic sediment framework (after Défarge et al.[47])
3 与蓝细菌有关的钙化作用
3.1 蓝细菌的CO2浓缩机制
水生生物光合作用至关重要的两个因素:一个是环境中可获取的溶解无机碳(DIC),另一个是在酶的作用下将这些无机碳进行初步固定的机制—二氧化碳浓缩机制(CO2concentrating mechanisms,CCMs)[74]。CCMs 在光合作用生物体中广泛存在,其作用是将CO2浓缩在细胞内,从而增加固碳量。虽然不同种类生物体的CCMs具有差异,甚至具体机制还未获得充分理解,但是总体过程具有一致性,包括HCO3-的传输、CO2的扩散和转化等主动的向细胞内的碳转移[75]和碳酸酐酶(carbonic anhydrase, CA)将HCO3-转换为CO2而为蓝细菌所用的过程(图5)。这一过程能够将细胞内CO2浓缩至细胞外的1 000倍[76-77],使得CO2在水中的扩散速度是空气中的1/104的情况下也能被蓝细菌有效利用[78]。
3.2 与光合作用有关的蓝细菌钙化作用
CCMs 导致钙化作用是多种蓝细菌的突出特征[3-4,23,74,79-80],碳酸盐饱和度的提高主要通过其中一种或两种途径:新陈代谢提高细胞周围EPS 或者蛋白质表层(S-layer)微环境中的pH值和钙化所需离子浓度[3,81-84]。pH 值的提高主要通过以下两个途经:1)依赖于羧酶体中碳酸酐酶的活动,消耗细胞溶质内H+(或者产生OH-),促进反应(1)向左进行,导致细胞溶质内OH-的增加,并输出或扩散到细胞外;2)通过光合作用电子在叶绿体类囊体中的传输(图5)和等离子膜中Ca2+或者H+的反向运输(输出Ca2+和输入H+),促进的平衡反应(2)和(3)向右边进行,增加细胞表面PH 值和的浓度。离子浓度提高主要通过EPS中的碳酸酐酶将周围环境进入的或细胞质溢出的CO2转变为和,而增加其中的浓度;细胞周围环境Ca2+和Mg2+的补充[86]和细胞内Ca2+的输出是Ca2+和Mg2+离子浓度提高的主要途经。
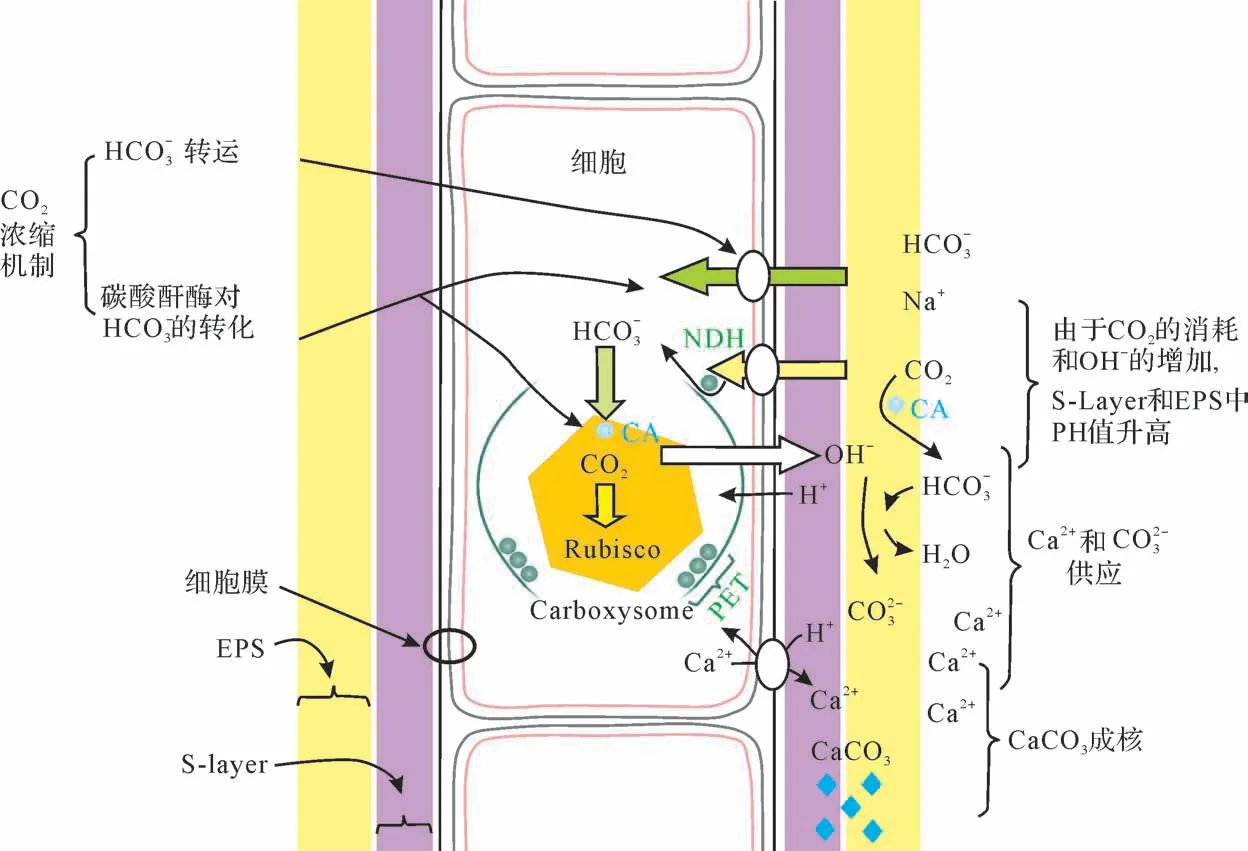
图5 活体蓝细菌的光合作用碳浓缩机制促进的鞘钙化综合模型(据Kamennaya et al.[13],Jansson et al.[41]和Riding[74],有修改)NDH:还原型烟酰胺腺嘌呤二核苷酸磷酸(NADPH)脱氢酶;CA:碳酸酐酶;PET:光合电子传输;Rubisco:二磷酸核酮糖羧化酶;Carboxysome:羧酶体Fig.5 Integrated model of in vivo cyanobacterial sheath calcification by CCM-enhanced photosynthesis(modified from Kamennaya et al.[13], Jansson et al.[41], Riding[74])
CCMs 导致蓝细菌的钙化主要是EPS 或S-layer内发生的碳酸钙沉淀,由于S-layer 不属于EPS,本文不展开论述。如果整个鞘被碳酸钙晶体侵染,鞘的外形就能被保存下来,保留蓝细菌管状或灌木状等形态[11],这便是我们常说的“蓝细菌化石”,但它只是保持了细胞外形的EPS 钙化产物而已,鞘内的单细胞或毛状体并没有发生钙化而是降解消失[11,74,87]。部分的EPS 钙化后会从细胞表面脱落,而且会有新的EPS产生。如果蓝细菌细胞完全被CaCO3包埋,由于不能摄取营养物质而死去的现象也时有发生[86]。图6 展示了现代美国巴哈马滩中叠层石内活着的蓝细菌Dichothrix正在发生鞘的钙化现象[88]。如果只是在细胞附近沉淀了一些孤立的晶体,就无法形成一个可保留的结构,而是随着EPS 的溶解而扩散到外部空间,以外来颗粒(如“白垩”)的方式释放,在湖底或者海底堆积形成大量微米级碳酸盐泥的沉积物[11,87,89]。白垩化现象是快速的大规模细粒碳酸钙和有机质的混合沉积,能够导致整个湖泊水体变成牛奶状[86,90],例如北美密西根湖就曾经发生过此类现象[81,91]。但是,现代海洋白垩化现象是如上所说浮游蓝细菌钙化还是灰泥的重新悬浮所致还存在较大的争议[92-94]。

图6 美国巴哈马滩叠层石内蓝细菌Dichothrix 鞘的钙化特征(据Planavsky et al.[88])(a,b)钙化的蓝细菌丝状体;(c)钙化蓝细菌丝状体的SEM照片,细粒的文石沉淀在蓝细菌细胞/毛状体(T)周围黏液状的鞘(S)内Fig.6 Calcification of cyanobacterial sheath of Dichothrix in a stromatolite layer in the Bahamas (after Planavsky et al.[88])
蓝细菌通过光合作用导致鞘内发生钙化的两个条件是低的溶解无机碳和高的钙离子浓度[3,95-96],且钙化现象主要发生在现代淡水环境中,很少发生在海水[97]或超咸水环境中[14,98-100]。需要引起注意的是大多数蓝细菌都具备CCMs的能力[101],但不是所有都存在钙化现象,特别是海洋环境中,同时也有报道蓝细菌在没有光合作用的情况下也钙化的现象[102]。显然蓝细菌的钙化作用机理仍然是值得深入探讨的问题。

钙化作用对蓝细菌来说是把“双刃剑”,既能保护细胞,例如使生活在浅水或高海拔地区的蓝细菌免受高强度光照损害其光合作用机能,同时缓冲细胞周围微环境由于光合作用升高的PH值,从而有利于高速率的光合作用[102],也能导致蓝细菌过度钙化被包埋,特别是因钙化加重而使浮游类沉没于有利光照带以下而死亡[84]。
2012年Science上报道了活着的蓝细菌细胞内形成不定形富钙碳酸盐[103](图7)的认识打破了人们对蓝细菌只有细胞外钙化唯一方式的固有认识,并且后来的研究显示多达68 种蓝细菌具有这一能力,并认为之前的研究忽视了这一重要现象[104-106]。虽然人们对蓝细菌细胞内钙化现象的机理认识还不清楚,目前也没有找到地质证据支持地质历史时期的蓝细菌也存在这一特性,但是这一进展开启了蓝细菌钙化作用研究的新领域,可能挑战着蓝细菌钙化作用的地质记录[107]。

图7 蓝细菌细胞内不定形碳酸钙沉淀的SEM 图像(据Couradeau et al.[103])Fig.7 SEM micrographs of amorphous CaCO3 incorporated in a cyanobacterium cell (after Couradeau et al.[103])
3.3 蓝细菌死后的钙化作用
成熟的微生物席通常是由高度有序组织的多个层状微生物群落组成的有机共同体,包括顶部的蓝绿色蓝细菌层(生氧光合细菌)及向下紧邻的棕色需氧异养细菌、中部的微红—粉红色紫色硫细菌层(不生氧光合细菌)、中下部的绿色硫细菌层(厌氧光合细菌)以及下部的硫酸盐还原细菌层(厌氧异养细菌)[1,108-109](图8a,b)。蓝细菌是共同体中有机质(主要是EPS)的主要生产者,其他异养微生物均对EPS具有生物降解作用。

图8 巴西东南部Vermelha 潟湖内叠层石沉积特征(据Vasconcelos et al.[108];Spadafora et al.[109])(a)潟湖内底部发育穹窿状叠层石和从水中捞出的叠层石单体(高10 cm,宽30 cm)[109];(b)叠层石顶部发育的微生物席微剖面,碳酸盐(白色层)和未石化的有机质层间互产出[108]。左边标尺显示剖面层厚大概3 cm,右边显示采样位置;(c)层1中的钙化蓝细菌丝状体沉淀方解石[108];(d)和(e)层2和层3中的球状和椭圆状的高镁方解石[108]Fig.8 Sedimentary features of stromatolite in Vermelha Lagoon, southwestern Brazil (after Vasconcelos et al.[108];Spadafora et al.[109])
蓝细菌死后的钙化作用主要是微生物降解过程中及降解后的钙化作用[17-20,110],其中前者是微生物席内钙化作用的主要方式。这两种钙化作用是典型的有机矿化作用,分别称之为生物诱导的矿化(biologically-induced mineralization)和生物影响的矿化(biologically-influenced mineralization),与生物控制的矿化(biologically-controlled mineralization)差异明显[17-20]。生物控制的矿化作用强调生物的活动直接控制了矿物的成核、生长、形态和最终位置,形成生物外部或内部的骨骼[111-113];生物诱导和生物影响的矿化作用则强调有机质对矿物形成过程的作用,其中前者必须是生物体存活时其新陈代谢活动改变了生物有机质中矿物的形成条件(碱度、饱和度等),因此是一种生物活动的主动过程,而后者主要是源于生物(生物体分泌或分离开,或者遗迹和死亡有机体的副产物[110])或非生物的有机物质对矿物形成(晶体形态和组成)的影响作用,因此是一种被动过程[46]。因此,之前介绍的与蓝细菌光合作用有关的钙化应该属于生物诱导的钙化作用。
微生物席共同体建造生长过程中不同功能微生物群落代谢过程能对碳酸钙产生沉淀和溶解不同作用,大致可以分为6 种主要的化学代谢反应,光合作用(包括生氧和不生氧)和硫酸盐还原反应通过增加微环境碱度(pH值)促进沉淀,而呼吸作用、硫化物的氧化反应和发酵作用产生的碳氢酸和有机酸则很可能会导致碳酸钙溶解作用的发生[11,46,114-117](图9)。因此,碳酸钙沉淀程度取决于以上代谢作用导致的溶解和沉淀之间的平衡,且蓝细菌EPS 的降解过程起到了重要作用[109]。

图9 微生物席垂向微环境梯度中细菌群落的活动与碳酸钙的沉淀—溶解作用(据Dupraz et al.[46],Visscher et al.[116],Visscher et al.[114],Kazmierczak et al.[118],有修改)(a)微生物席中氧气、硫化物和pH 值在24 h内的变化,剖面I 和II 分别为下午2 点和早上3 点的“地球化学快照”,可以最显著地反映出微生物席在白天和黑夜之间的不同;当黑夜降临时,由于光合作用的停止和异养细菌的消耗,微生物席很快就转变为缺氧环境[46,116];(b)发生在一个典型的微生物席中的6 种主要化学代谢反应,这些反应按对碳酸钙沉淀作用的影响分类;光合作用和硫酸盐还原反应通过增加微环境碱度而促进沉淀,而呼吸作用、硫化物的氧化反应和发酵作用则很可能会导致溶解作用的发生;当黑夜降临时,需氧细菌的活动逐渐停止,厌氧的异养微生物的活动开始占据主导地位;碳酸钙的净沉淀量取决于不同类型的微生物的新陈代谢活动及其时空分布及变化[46,118]Fig.9 Activity of microbial communities in a microbial mat under vertical microenvironment gradients leading to carbonate precipitation and dissolution (modified from Dupraz et al.[46]; Visscher et al.[116]; Visscher et al.[114] ; Kazmierczak et al.[118])
在海洋和超咸水微生物席中,硫酸盐还原细菌在整个的微生物群落导致碱度变化而促进碳酸钙沉淀过程中起到关键作用[46],超过90%的缺氧呼吸作用都是硫酸盐还原细菌完成[114]。硫酸盐还原细菌生物降解EPS能够释放其中的Ca2+和Mg2+等离子,同时消耗了SO2-4和有机碳化合物,提高环境碱度[14,46,115],从而产生生物诱导的钙化作用。EPS 降解后的钙化作用主要发生在微生物席底部,微生物席的这部分环境更为封闭,EPS在上部微生物降解带内被硫酸盐还原细菌、产甲烷细菌等“终极”细菌降解后残余的EPS能为碳酸盐矿物的沉淀提供成核中心,同时其离子束缚能力在持续的离子供给条件下达到饱和,能产生自由的Ca2+和Mg2+等离子,影响微生物席底部的微环境地化性质,进而在残余的EPS 内形成钙化作用的场所,促进了碳酸盐的沉淀[11,46,115-116],产生生物影响的钙化作用。碱性水体季节性上涌后的蒸发作用能形成以上条件产生钙化作用[117]。
大量研究显示蓝细菌死亡后的有机物质在降解过程中,由于异氧微生物的作用形成有别于蓝细菌活体钙化矿物的半球状镁方解石/文石[108](图8c~e)、亚微米级的不定形文石颗粒[118-119]、纳米级球状镁方解石[99]、哑铃状高镁方解石[120-122],并且在已成鞘的丝状体中,鞘的降解导致了厚度和连续性都可变的不规则的外壳[120],鞘的形态和大小都发生了变化[27]。这些现象的普遍揭露甚至使得Altermannet al.[1]认为海相环境下蓝细菌席的钙化主要是被动过程,蓝细菌的直接钙化潜力和光合作用对钙化过程的影响很微小,碳酸钙的沉淀主要是通过在黏质鞘和EPS 里的自溶和菌溶作用形成。
研究者注意到白云石的形成,包括成核和稳定生长过程都与蓝细菌的EPS 降解有很大的关系[123]。在氨基酸和源于氨基酸的有机质降解带内非常高镁含量的方解石和不完全有序的钙白云石等富镁碳酸盐沉淀,形成白云石的先驱矿物[116,124-125]。值得重视的是,硫酸盐还原菌[126],适度适盐需氧细菌[127]和产甲烷古细菌[128]等产生的EPS也是控制白云石沉淀的重要因素[129]。
4 蓝细菌的EPS 在与其有关钙化过程中的作用
蓝细菌新陈代谢过程中产生的EPS 形成了与外界物质交换的通道,并提供了微生物席内异养生物重要的营养供给。Shearmanet al.[130]于1965 年第一次提到了EPS 在蓝细菌钙化中的作用。Merz-preiß[3]在2000 年注意到自然条件下分泌鞘的蓝细菌才钙化,而缺乏鞘的蓝细菌不钙化的现象,但也并不意味着所有在碳酸盐丰富环境和具有鞘包裹的蓝细菌都会有钙化现象,并引起了广大学者关于EPS 与钙化作用关系的研究。
微生物席内异养细菌,例如硫酸盐还原细菌的EPS 生产潜力也被证实[131-132],上述适度适盐需氧细菌[127]、产甲烷古细菌[128]等其他微生物的EPS 在钙化中的作用也受到重视。本文仅以微生物席内最重要的EPS 生产者蓝细菌为论述对象,介绍对与其有关的微生物钙化作用的影响。
4.1 蓝细菌结壳作用
(1)带负电性 EPS形成蓝细菌表面带负电的微环境,能够从周围水环境中黏合Ca2+和Mg2+等二价离子并减少扩散速率[93],改变矿物成核和晶体生长的动力学路径[48],有利于碳酸盐的饱和而促进CaCO3的成核生长。EPS 中带负电的有机物质(例如多糖)控制了不定型碳酸钙前驱的稳定性[133]。
(2)提供成核位置 细胞表面EPS中的谷氨酸、天冬氨酸或者羧基、磺酸基等束缚Ca2+或者Mg2+的有序负电酸性功能团能成为初始的CaCO3成核位置[134],同时EPS 还能降低方解石成核核心的界面自由能[84],而促进蓝细菌的钙化作用。大多数蓝细菌的鞘或者荚膜中碳酸钙的饱和度都很高,但是方解石在多糖中的成核速率是由其酸度决定的。现代泉华中主要由丝状蓝细菌产生酸性外聚物提供一致的显微成核位置,形成层状结构的叠层石;球状和丝状蓝细菌产生酸性或非酸性的外聚物,提供非均一的显微成核位置,形成了凝结结构的凝块石[134]。
(3)高含水特性 有利于离子的聚集和浓缩[52],促进Ca2+和Mg2+的吸附聚集。高含水的EPS 中部分有机大分子由于附着在晶体表面而限制晶体生长,但是如果这些大分子稳固的附着在固体的基质上时就有利于晶体的结核生长[135-137]。
4.2 蓝细菌降解过程中的钙化
EPS 的内部特性(例如物理化学特性)通过影响饱和度和可用的成核位置对蓝细菌降解过程中的钙化作用起到了阻碍和促进碳酸钙形成的双重作用[46,138]。初始生成的EPS 由于对阳离子的吸附而阻碍钙化作用,但生物(生物降解作用)或非生物(水解和紫外线的光解作用)的作用能够改变EPS 对钙化的阻碍作用[14,139]。
(1)阻止碳酸钙的沉淀 酸性多糖等有机高分子能够通过他们所组成的自由羧酸盐、酸性氨基酸(例如天冬氨酸和谷氨酸)和(或)硫酸盐基团来吸引和黏结二价阳离子,如Ca2+、Mg2+、Sr2+,并与其发生反应[17,70],但聚阴离子(能够发生离子聚合反应的阴离子)与Ca2+的结合能阻止其与CO23-的成核和结晶生长。EPS 的离子吸附能力导致自由的Ca2+从周围溶液中移走而被消耗,从而限制了周围环境中钙碳酸盐的沉淀[46]。具有丰富糖醛酸的多糖通过黏合Ca2+对钙化晶体成核和生长具有调节功能,否则不受控制的钙化会导致细胞的损伤[1]。因此,在碱性水体中即使对于文石或方解石是超饱和的,由于没有自由的Ca2+而不能沉淀[46],形成生物限制的“抗钙化作用”[136]。
(2)促进碳酸钙的沉淀 由于异氧细菌以蓝细菌的EPS 多糖为食物,细胞溶素作用以及自动分解和水解作用使得EPS 和多糖-蛋白质复合物中的Ca2+、Mg2+或者释放,促进碳酸盐过饱和以及沉淀,产生生物诱导的矿化作用,并且由此产生具有一定的离子有序度的CaCO3及其他矿物的微小不定型前驱[140-142]。通过现代研究实例证明,通过光合作用产生的总碳中有3%~4%来源于生成的EPS,之后在24 小时内有40%~60%的新生成EPS 会被降解,净EPS产率较低且维持在产生与消耗的动态平衡中[65]。新生成的EPS 通过络合钙离子并不能直接形成钙化,当离子络合达到饱和[117],或者当EPS通过微生物或化学过程降解时,大量的钙离子被释放,形成自由的Ca2+、Mg2+或者HCO-3,促进钙化的产生[14]。
4.3 控制钙化过程及产物
(1)控制矿物的类型 蓝细菌微生物席中CaCO3成核实验证实EPS在钙化层引导了文石晶体的形成;在非钙化层引导了方解石的形成。EPS 的生物地球化学组成,特别是小的蛋白质影响了CaCO3的矿物形成[15-16]。EPS中部分的溶解有机碳影响了Ca2+的络合作用,从而降低了碳酸盐的过饱和,使得溶解度低的方解石沉淀而不利于溶解度高的球霰石沉淀,并且EPS中的有机分子能阻止球霰石向方解石转化[143]。
(2)控制矿物的形态 由于EPS 有机分子吸附在晶体表面并影响表面活性,同时胶状的特性影响钙离子的扩散、吸收、络合[46],从而限制了晶体的快速生长,形成球状、哑铃状或光滑的菱形晶体表面[144]。EPS 能使得更小、更多和更圆的晶体形成,同时还能促进基于聚集的晶体生长机制[48,145],形成球形小晶体聚集体[146]。但是需要注意的是,具有胶状特性的非生物的硅胶中也能形成球状、哑铃状、麦穗状束或菱形的碳酸盐矿物[147]。因此,化石记录中球状或哑铃状的形状不是EPS有关的有机矿化直接证据[148]。
(3)控制矿物的稳定性 细胞外内层紧密结合的EPS 的数量、有机地化组成和结构控制了其中沉淀的不定形碳酸钙先驱矿物形成后向热力学更稳定方解石的转化过程,有机大分子能稳定不定形碳酸钙,形成蓝细菌免受细胞外不受控制碳酸钙沉淀包埋的“保护机制”[84]。
(4)提供钙化稳定结构和晶体生长空间 蓝细菌降解过程中,重新组织成相互连接且相对牢固的三维网状结构能够为钙化形成叠层石提供稳定的框架。同时形成的局限空间也能限制碳酸钙矿物晶体的生长,约束晶体的大小[47]。
4.4 控制白云石的形成
自从Vasconceloset al.[149]在巴西LagoaVermelha海岸潟湖发现了与硫酸盐还原细菌共生的铁白云石,且通过模拟实验证实嗜硫细菌明显诱导了白云石的形成后,地质学家普遍认为新陈代谢活动是低温白云石沉淀的关键因素,“微生物白云岩”的概念开始盛行。但是随后的实验和实地考察中发现在没有微生物参与或新陈代谢不活跃的情况下,一些有机大分子、尤其是羧基或羧基化聚合物仍然能够诱导白云石的沉淀,说明起作用的并非微生物的生理活性[150-152]。值得注意的是包括蓝细菌在内的各种微生物的EPS 是白云石成核和生长的主要场所,为白云石成核提供了有机模板和有利的化学微环境,且EPS 中有机大分子能够减缓碳酸盐矿物从晶核状态向化学计量晶体转化的进程[153]。
(1)白云石初始成核:低温情况下,不同降解速率的EPS 内重新排列的酸性功能团组,可能导致糖类和氨基酸中带负电的配体具有不同的调整几何结构和吸附Ca2+和Mg2+的特征[69,116,139,142,154-155],使其能够成为白云石初始成核模板[20,156]。降解过程中EPS 中逐渐释放到周围溶液中的离子沉淀成不定形碳酸盐矿物[141],可能阻碍了一定程度的阳离子排列[142]。一旦有利的物理化学条件在稳定情况下存在,不定形碳酸盐矿物便能导致白云石的生长[156]。
实验证实形成稳定的超级核心(比临界尺寸大的团簇)很关键,因为他们能自发生长[157]。EPS 中羧酸盐有机物质和细胞壁中的羧基具有使镁脱水的功能是成核的主要机制[151-152]。死亡细胞由于排出水分子而使EPS 中羧基官能团吸附的[Mg(H2O6)2+]形成[Mg(H2O)5(R-COO)]+复合体,随后CO2-3替代水分子在细胞壁外形成表层的MgCO3(H2O)4(R-COO)复合体,并在碱性条件下成为附着的Ca2+和另外的CO2-3成核模板成为团簇。这个机制需要稳定提高的PH值而保持溶液中相对高的CO2-3离子活性[158]。
(2)白云石的稳定生长:EPS中的有机大分子在降解和浅埋藏的成岩过程中持续提供的离子保持有利于白云石形成的Mg∶Ca 离子比例,持续保持浅埋藏白云石的沉淀反应[123]。EPS 中羧化的高渗透性复合物对硫酸盐还原和产甲烷微生物至关重要[159],且对生物反应的金属离子进入浅埋藏成岩作用的贡献对白云岩成核后的继续稳定生长具有重要作用[123]。因此,在地表下埋藏沉积物中EPS 的降解过程可以控制成岩白云石的形成。值得注意的是,大陆边缘埋藏于几百米下的腐泥中通过16S RNA分析发现具有和现代潮缘环境非常相似的微生物群体[160-162],而且新近系与EPS共生的深海细晶白云石的存在是否预示更深的成岩白云石与EPS有关呢?
5 地质意义及展望
5.1 地质意义
蓝细菌钙化化石及生物(有机)矿物可以记录整个地质历史时期新陈代谢演化过程[108]。蓝细菌的钙化产物是其与地球系统协同演化留下的宝贵地质记录,为我们打开了一扇窥探地球演化历史及生命起源和演化奥秘的窗口。蓝细菌的EPS在与其有关的微生物钙化过程中作用研究的地质意义体现在以下三个方面:
(1)蓝细菌钙化化石本质为细胞外鞘的钙化作用的结果,微生物碳酸盐岩就是微生物席内EPS 有机矿化作用的产物。在完整微生物化石缺失的情况下,钙化的EPS 也记录着微生物与环境协同演化的重要信息。以蓝细菌EPS与微生物席钙化之间的关系为主要线索,探寻岩石记录中微生物化石、生物(有机)矿物和EPS矿化产物与EPS之间的成因联系,将有望获得微生物地质记录形成原因的新认识。
(2)由现代蓝细菌EPS 的属性差异控制不同微生物碳酸盐形成的研究结果引申出蓝细菌进化和绝灭事件过程中EPS属性变化控制了元古代和寒武纪界线凝块石和钙化蓝细菌的爆发事件的重要推论[134],为我们提供了一个从地质事件中蓝细菌EPS属性变化的角度探索地质历史时期微生物碳酸盐岩的形成和演化的新视角。
(3)通过对比分析能够建立蓝细菌EPS 属性的变化与早期地球环境的一些边界条件之间的关系,有望搭建微生物新陈代谢过程与古环境、古气候协同变化之间的重要桥梁。探索古环境和古气候波动过程中蓝细菌EPS 的变化响应及其导致的化石、生物(有机)矿物和EPS残余物及矿化产物的相应变化可能是我们获得微生物新陈代谢活动与地球系统协同演化过程认识的新思路。但是,如何从矿化记录中提取物理和化学信息重构EPS 的变化,反演微生物矿化过程是我们面临的一个重要难题。
5.2 研究展望
蓝细菌EPS 在微生物钙化过程中的作用研究日益受到地质微生物学、微生物工程学和仿生学等不同领域研究者的重视,但只是触及到冰山一角,关键机理问题还不得而知。受复杂成岩作用和变质作用影响,EPS 残余物及矿化产物将受到极大的改变,而且在取样过程中EPS的残余记录仍然会发生改变甚至完全丢失。因此,在地质记录中探寻蓝细菌EPS的变化及其与微生物钙化作用的关系显得尤为困难。针对上述困难和问题,就本文讨论内容提出以下研究展望:
(1)充分利用好“将今论古”这把金钥匙,加强现代各类环境中蓝细菌EPS和与其有关的微生物钙化成因联系和实验模拟研究,以期获得地质记录的现代对等物和各类成因指标。同时,也要参考地质时期的环境、气候、构造、生命演化等变量的影响,建立上述变量和蓝细菌EPS有关的微生物钙化之间成因联系,并谨慎使用“将今论古”这把钥匙。
(2)除蓝细菌以外的各类微生物同样也具有产生EPS的潜力[131-132]。它们在个体的矿化和微生物席的矿化过程中EPS 发挥的作用同样应受到重视,并注意它们与蓝细菌EPS的识别和区分。
(3)微生物地质学家们在探寻地质记录中微生物化石的同时,应注重EPS 残余物和矿化产物的特征分析,建立EPS 与微生物矿化以及生物(有机)矿物之间的成因联系。
(4)现代沉积和模拟实验分析均已揭示白云石的形成与微生物EPS 有很大关系[150,158]。加强地质历史中白云石分布与成因和微生物EPS之间的成因联系研究,有望获得白云石成因的新线索。
(5)充分利用现代高精度实验技术和手段进行地质样品中EPS残余物和矿化产物原位微区取样和物理和化学信息的提取。原位微区取样技术主要是扫描电镜联用的聚焦离子束,实验过程要注意镓离子和剥蚀对象的污染和对样品的损害[163]。原位的地球化学分析包括扫描电镜和拉曼光谱联用[164]、透射电镜及选区电子衍射[165]、扫描透射X射线显微镜及X射线吸收谱[166]、纳米二次离子质谱技术等[167],能够获得显微成像、有机分子、晶体学、元素及同位素等原位信息。
(6)加强EPS的埋藏学分析,包括岩石记录中的EPS 残余物和EPS 的埋藏实验模拟两个方面。EPS的埋藏多样性与微生物席的沉积记录存在最为紧密的成因关联[168]。EPS 残余物分析包括保存形态、物质化学组分和生物分子等方面,目的是还原EPS 的物理和化学属性。同时,EPS的埋藏实验模拟主要是分析EPS 在埋藏过程中的物理和化学性质变化,为岩石记录中残余EPS的属性反演提供模拟参数。
致谢 感谢两位审稿专家给出的意见和建议。
猜你喜欢
中国高新科技(2022年16期)2022-12-07
河北地质(2022年2期)2022-08-22
睿士(2021年5期)2021-05-20
河南冶金(2021年1期)2021-04-27
睿士(2020年5期)2020-05-21
青少年科技博览(中学版)(2020年1期)2020-04-21
作文评点报·低幼版(2019年46期)2019-12-30
意林·全彩Color(2019年11期)2019-12-30
睿士(2019年9期)2019-09-10
Coco薇(2015年12期)2015-12-10
