
高山松自然地理分布、生长与生态适应性研究进展
2022-02-18 15:51杜燕包维楷
四川林业科技 2022年6期
杜燕, 包维楷
1. 中国科学院山地生态恢复与生物资源利用重点实验室,生态恢复与生物多样性保育四川省重点实验室,中国科学院成都生物研究所,成都 610041;
2. 中国科学院大学,北京 100049
高山松(Pinus densataMast.)隶属于松科(Pinaceae)松属(Pinus),是云南松(P. yunnanensisFranch.)和油松(P. tabuliformisCarrière)的天然杂交种,产于四川西部、青海南部、西藏东部及云南西北部,是我国西部高山地区的特有树种[1]。高山松是木本裸子植物中目前唯一被分子生物学证明的天然同倍体杂交种[2],是研究木本植物同倍体杂交种形成机制的模式物种之一,具有很高的科研价值。同时,高山松也是可用于植被恢复和绿化的优良树种[3],其枝干、针叶及松脂在畜牧、造纸和医学等行业具有较好的应用前景[4-7],具有较高的生态和经济价值。
本文作者系统查阅了1956—2021年发表的高山松个体研究相关文献资料,包括文献66篇,专著6本,研究资料集(刊)1本,行业标准1部,综述以下3个方面的研究现状:(1)高山松地理分布格局特征;(2)高山松物种的起源与演化;(3)高山松个体生长规律及其环境影响机制;并进一步梳理出当前需要聚焦的方向,为高山松的深入研究提供基础。
1 高山松的地理分布格局特征
根据相关植物志记录,高山松的分布区以四川西南部为中心,北达青海尖扎,南至云南丽江,西起西藏工布江达太昭,东界四川松潘贡冈岭,约为北纬27°~36°,东经92°~104°[1,8],而后续的文献[9]和标本记录(数据来源于中国数字植物标本馆https://www.cvh.ac.cn)表明其分布区域较之前的记录更广,西界桑日,在加查、隆子、墨脱等地也有分布。其在水平分布上并不连续,在然乌至怒江间有一中断[10]。海拔分布范围一般在2 600~3 500 m,是松属中分布海拔最高的物种[1],最高上限可达4 350 m,见于得荣,最低下限海拔1 700 m,见于宝兴赶羊沟[8]。在川西和滇西北地区,从东到西,由北向南,海拔分布范围逐渐升高[8,11-13]。
近年来,有研究对当前气候变化背景下高山松的适宜分布区进行了分析,结果均表明高山松的适宜分布区呈扩张趋势,Mao等[14]基于生态位模型,认为青藏高原地区的高山松可沿东南部的平行山谷进行扩张;陆双飞等[15]基于最大熵模型的预测结果表明,在2 070 RCP4.5 气候情景下,高山松适宜区和最适宜区面积分别增加了26.86%和19.74%,适宜区质心向西偏北的高海拔高纬度地区移动。
2 高山松物种的起源与演化
2.1 起源
高山松的起源存在着大量研究,获得了多方面的证据,结束了多年的争论。吴中伦[16]最早提出高山松起源的两个假设,一是高山松为云南松和油松的杂交种,二是高山松为云南松和油松的祖先,由其向南演变为云南松,向北演变为油松。综合针叶形态与解剖结构[17]、染色体构型[18]、线粒体与叶绿体DNA[19,20]、5S rDNA基因序列[21]、同工酶技术[22]、SSR标记技术[23]以及电阻抗测量[24]等多方面的相关研究证据,表明了高山松是油松和云南松的天然同倍体杂交种,证实了吴中伦的假设:高山松是云南松和油松的杂交种。进一步的研究表明,高山松起源于中新世晚期,最初的杂交起源地位于当前分布区的东北边缘,向西迁移扩散至青藏高原,群体分化开始于上新世晚期,青藏高原东南部的隆升促进了种的形成[25-28]。
2.2 进化机制
目前关于高山松物种进化机制的研究,实质上是探讨高山松为什么能分布在亲本种无法正常生长的高海拔生境。已有研究从苗期适应性、生殖隔离、基因与酶的结构和功能等方面开展了研究。苗木异生境萌发与栽培试验表明,在高山松生境下,高山松具有明显高于亲本的适合度优势,且亲本种不可能侵入高山松生境;高山松在亲本种生境下苗期均表现出正常的适应性,大部分性状居于亲本种之间,且不同种源间萌发与生长特征存在明显的分化,来自起源中心的种群适应性较好,表明生态选择在高山松的同倍性杂交物种形成中起到了关键作用[29-34]。从苗木生理生态特征看,高山松储备光能的能力远大于亲本种,而消耗光能的能力最小[35];在干旱和低温胁迫下,高山松在光能的吸收、传递和利用,水分利用效率和干物质积累等方面均表现出超亲优势[36,37];氮添加试验表明高山松的氮利用效率和光合氮利用效率均高于亲本种[38]。有研究表明,高山松与其亲本种尚未形成完全的生殖隔离,种间人工授粉成功率较高[39];在横断山区对云南松和高山松花期的观测发现,气象因子的不同导致云南松的散粉期明显早于高山松,花期不遇阻碍了两个物种间的基因交流,进而隔离开来[2,40]。还有研究以miRNA,DHAR等基因以及tau型谷胱甘肽转移酶的结构和功能探讨高山松的生态适应性和进化机制[41-44]。
3 生长规律及其环境影响机制
3.1 生长规律
3.1.1 高生长
陈起忠等[5,45-51]先后构建了高山松高生长拟合方程(见表1),蔡年辉等[30]建立了不同种源的高山松在1年内的高生长Logistics拟合曲线。陈端[3]发现林芝地区的高山松一年中高生长期在3—6月,约100 d。根据已有文献资料记载,高山松高生长在前5年比较缓慢,连年生长量在2.5~18.0 cm,6年开始增快,连年生长量在20年左右达到最高值,为50~61 cm,30年后处于相对稳定状态并开始减缓,170年时树高生长基本停止[12,46,53-56]。
3.1.2 径向生长
陈起忠等[45,46,56]等先后构建了胸径及其生长率拟合方程(见表2),蔡年辉等[30]建立了不同种源的高山松1年内的地径生长Logistics拟合曲线。陈端[3]发现林芝地区的高山松一年中径生长期在4~9月,约180 d。部分研究对高山松年均生长轮宽度进行了测定,在西藏地区和金川县分别为3.3 mm和2.14 mm[9,57],在香格里拉市高山松边材年均生长宽度为0.112 mm[58]。径向生长前期与高生长相似,连年生长量在20年左右达到最高值,为0.45~0.68 cm,70年后连年生长量迅速下降,径向生长可持续至170年以上[46,52,54,55]。

表 1 高山松高生长拟合方程Tab. 1 Fitting equation of height of Pinus densata

表 2 高山松径向生长拟合方程Tab. 2 Fitting equation of diameter at breast height of Pinus densata
3.1.3 根系生长
相关研究仅1例,郭立群[12]对滇西北1~10年生高山松苗木调查发现,在6年内,根径、根幅、根分支数数的增长缓慢,年均生长量分别约为0.16 cm,4.58 cm,2.75根,之后生长迅速加快,10年时连年生长量分别为2.00 cm,54.50 cm,23.50根;根深连年生长量在前5年逐渐增加,年均生长量为4.50 cm,第7年连年生长量可达7.50 cm,随后生长量缓慢下降。
3.1.4 树冠生长
相关研究仅1例,王小兰等[59]以林芝地区2041株样木构建了冠径-胸径二次函数关系:Cd=-0.001D2+ 0.244D+ 0.184(R2= 0. 866)。
3.1.5 生物量积累与分配
吴兆录等[5,51,60-63]先后构建了单木生物量回归方程,周丽等[34]建立了4年生苗木生物量估算模型(见表3)。郭立群[12]对滇西北1~10年生高山松苗木调查发现,4~6年以前地上部分生物量积累相对低于根系,之后地上部分生物量积累急剧增加。对林芝地区高山松解析木的分析发现,胸径在10 cm以内时,地上部分生物量积累缓慢,胸径达10 cm以后,地上部分生物量积累迅速提高;且随着立木胸径的增加,主干生物量所占地上部分生物量的比例增加,枝、叶所占比例下降;植株上留存的叶龄提高,一年生叶的比例逐渐下降,二年生叶的比例增高;主干生物量在垂直空间上由下向上递减,枝条和针叶生物量以树冠中部占比最大[5]。
3.1.6 材积生长
1978年,农林部颁布了含高山松在内的主要用材树种的二元材积表[64],曾伟生[65]基于新近采集的立木材积实测数据,对其进行了适用性检验,结果表明原有的高山松二元立木材积方程总相对误差和平均系统误差均超过3%,已不适用于目前的森林资源状况。陈起忠等[45,47,54,64,66,67]先后建立了材积、材积连年生长量和生长率相关模型(见表4 )。材积连年生长量持续的时间很长,在林芝和朗县等地90年时达到最高点,在波密持续上升的时间达到150年以上[54];在川西地区50~70年最高,为0.0074 m3[46]。
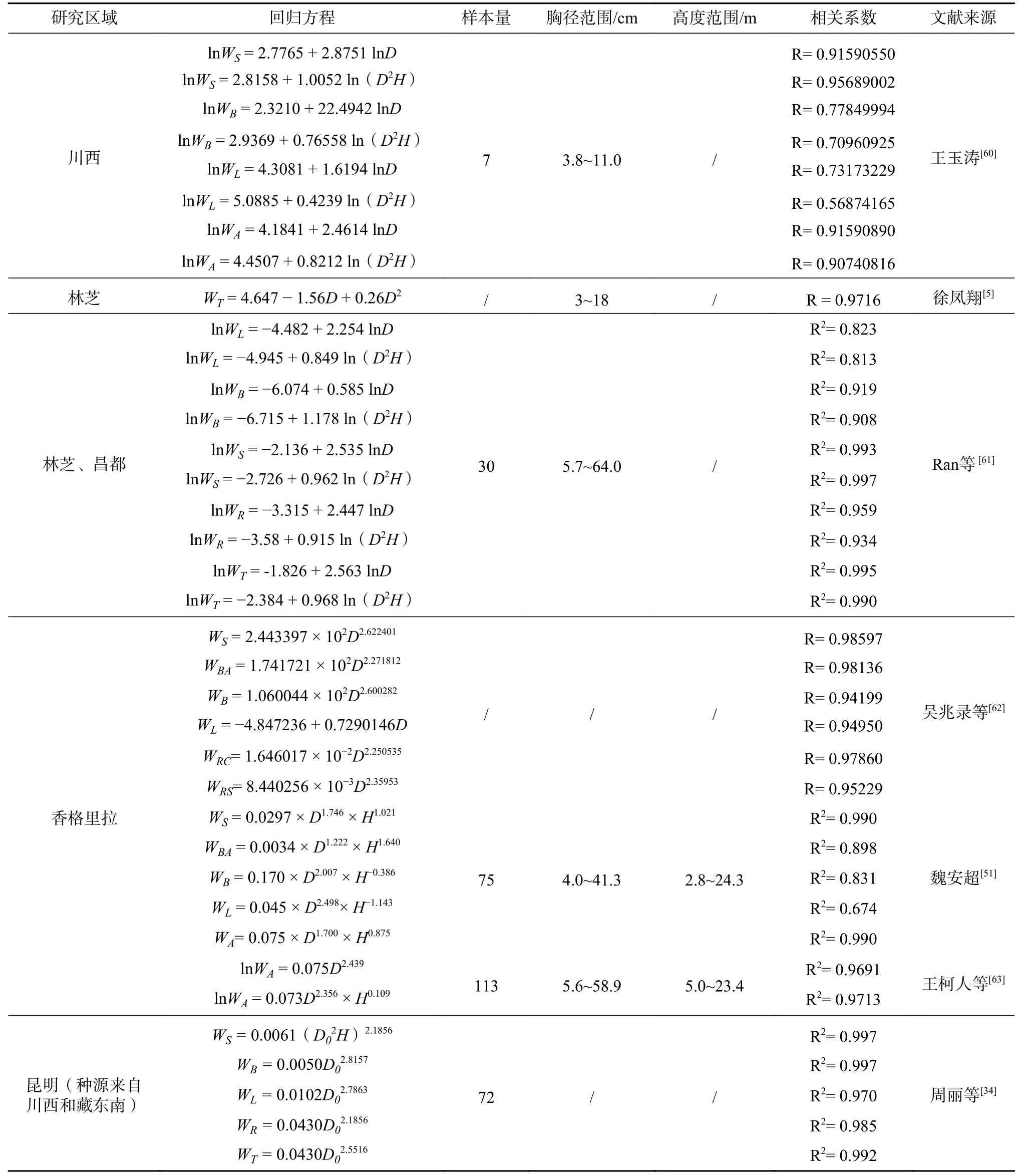
表 3 高山松单木生物量回归方程Tab. 3 Allometric equations relating individual biomass of Pinus densata

表 4 高山松材积拟合方程Tab. 4 Fitting equation of volume of Pinus densata
3.1.7 结实
根据资料记载,高山松结实早,且结实能力强,5~10年即开始结实,40~60年为盛果期,80~100年结实最好,200年时仍有结实能力;林分疏密度在0.4以下时,结实量较大,而疏密度0.6以上时结实量少[53,68]。
3.2 环境影响机制
3.2.1 气候
现有研究基于树轮宽度差值年表,采用滑动响应分析方法,分析高山松径向生长对气候的响应,结果表明其径向生长主要受生长季气候的影响,且在海拔分布上限,夏季气温是主要限制因子,海拔下限限制因子则为降水。在西藏南部地区主要受夏季降水的限制,年轮宽度与上年6—8月降水正相关[69];在金川县则主要受当年夏季气温和5月降水的影响,与当年5月降水和当年6—8月温度正相关,与当年6—7月降水负相关[57]。在石卡雪山海拔分布上限,7月温度是主要的限制因子,与当年7月平均气温和平均最高气温呈显著负相关,与降水未达到显著相关水平[70];在哈巴雪山海拔上限则主要受上年8月水热状况影响,与上年8月降水量显著正相关,与上年8月平均温显著负相关[71]。而对哈巴雪山海拔下限处的高山松研究表明,主要受当年5月水热状况影响,与当年5月降水量显著正相关,与当年5月平均最低气温和平均最高气温显著负相关[71]。还有研究[72]发现,在朗县地区,火干扰后高山松径向生长加快,且改变了对气候的响应模式,过火前幼树(DBH < 10 cm)径向生长与上一年11月平均最低气温显著负相关,成年树(DBH ≥ 10 cm)径向生长与当年9月平均最低气温和平均气温显著正相关,而过火后幼树和成年树的径向生长均与当年1月平均气温和平均最高气温显著负相关。
气候与高生长的研究仅1例,赵文智和荔克让[48]在雅鲁藏布江中游下段研究发现,阶地覆沙地高山松幼树高生长与4~6月降水量正相关,但不显著,而山坡覆沙地幼树高生长与4月平均气温呈显著正相关。
3.2.2 光照
高山松在4(6)年内耐荫蔽,对光照的需求随年龄的增加而递增[12,68]。徐凤翔[5]以更张和岗嘎林区60株高山松为研究对象,发现导致不同林分状况(林冠下、幼密林和空旷地)和不同更新年代的高山松生长差异的主导因子均为光照,在生长初期高生长量几乎相等,10年后空旷地生长量明显高于林冠下和幼密林,5年后更新年龄早的高生长领先于更新年龄晚的。郭立群[12](1982)对滇西北地区同一高山松林地上6种不同光照条件下高山松1~7年苗木高生长过程的调查发现,皆伐迹地空旷地高生长最快,年均高生长量可达14.16 cm,择伐迹地林窗中次之,为12.00 cm,其次为林窗边缘和母树树冠下弱度庇荫,分别为7.07 cm和6.69 cm,疏林林木间隙和林冠下生长较差,分别为6.11 cm和4.86 cm。
3.2.3 坡位
相关研究仅1例,徐凤翔[5]在更张和岗嘎林区研究发现,导致不同坡位高生长差异的主导因子为立地营养状况,坡下位的连年高生长量大于坡中。
3.2.4 地理空间格局
已有研究仅3例。徐凤翔[5]调查发现,从波密-更张-岗嘎,树高、胸径和材积的生长量均呈下降趋势,这是由于自东向西,降水量逐渐减少。毛建丰等[73]对香格里拉、林芝八一镇、林芝尼西乡、波密通麦镇、理县和小金6个样点的高山松球果13个种实性状的研究发现,其平均潜在繁殖能力随纬度的升高有下降的趋势。在海拔梯度上,陈甲瑞和王小兰[74]在林芝地区以海拔2 500 m为基点,自下而上分2 500~2 800、2 800~3 100、3 100~3 400 m共3个海拔梯度设置了调查样地,结果表明,随着海拔升高,高山松高生长相对减缓,胸径和冠幅生长速率逐渐增大,这便于提高同化能力,同时将更多的同化产物投入到与植物水分、养分输导有关的系统构建方面。
4 结论和展望
4.1 地理分布格局
关于高山松地理分布的资料年代均较早,受交通限制,可能仍有未知分布点。因此,需继续开展野外调查,明确其分布范围和边界,尤其是在与亲本种油松的分布交错地带白龙江流域,要注意物种的鉴定以免混淆,以形成完整的地理分布格局认识。
4.2 物种起源与演化
现有研究通过叶片形态和解剖结构以及分子生物学证据证明了高山松是油松和云南松的天然同倍体杂交种,推断出其起源和群体分化时间、起源中心以及迁移路线,并通过基因结构与功能、苗期生长性状、针叶形态解剖指标、光合与水分利用生理指标等探讨了其进化机制,结果表明高山松幼苗在形态建成、光能吸收利用、水分利用等方面具有超亲优势,能更好地适应低温、干旱的高海拔生境,生态选择是物种形成的主要驱动力,青藏高原东南部的隆升促进了种的形成。但目前对高山松的物种形成机制研究仍是不全面的,具体的起源地、迁移路线和演化规律尚不清楚。
4.3 生长规律及其环境影响机制
在生长规律方面,已有研究对高山松的高、胸径和材积生长过程进行了较为详细的论述,树高和胸径连年生长量均在20年左右达到最高值,材积连年生长量持续的时间很长,在50~90达到最高点,还构建了树高、胸径、生物量、材积等相关模型;冠幅和根系生长规律的研究较少;结实方面仅进行了果期年龄段划分,无结实量数据。对高山松冠幅生长规律、10年生以上个体根系和生物量生长规律尚无研究,高山松幼树(DBH≤ 6 cm)缺乏可靠的生物量估测模型。
目前已有的环境影响因子研究包括气候、光照、坡位、经度、纬度和海拔。气候因子的研究集中在径向生长方面,高山松径向生长对气候的响应存在“滞后效应”,在不同地区以及不同海拔段,影响高山松径向生长的气候因子及其响应模式不同。光照和坡位的研究集中在高生长方面,发现高山松苗木4~6年内耐荫蔽,而后随着年龄的增加对光照的需求增强;而由于立地营养状况的差别,坡下位的连年高生长量高于坡中。在小区域尺度上,高山松的生长量随经度的增加呈上升趋势,平均潜在繁殖能力随纬度的升高呈下降趋势。海拔的研究包括高、胸径、冠幅的生长,结果表明,随着海拔升高,高山松株高生长相对减缓,胸径和冠幅生长速率逐渐增大。当前研究仅对部分生态因子开展了生长过程差异的研究,并无机制方面的深入研究,各器官在生长过程中对不同生态因子的响应模式及其生理学机理、各器官间的权衡关系尚不清楚,在空间区域上的生长差异性缺乏大尺度的研究,高产区域未知。后续可对上述内容展开深入研究,以阐明高山松的生态适应策略。
猜你喜欢
江西农业(2022年8期)2022-05-13
林业科学研究(2021年6期)2022-01-05
内蒙古林业调查设计(2021年5期)2022-01-05
绿色科技(2021年21期)2021-11-26
林业科技情报(2021年2期)2021-07-13
内蒙古林业调查设计(2021年3期)2021-07-06
军事文摘(2018年24期)2018-12-26
现代园艺(2018年1期)2018-03-15
今日重庆(2016年2期)2016-08-11
中国新技术新产品(2011年5期)2011-05-09
