
不同种源银柴胡及其伪品的ⅠTS2序列分析与鉴别△
2022-02-18 08:23李振凯冯璐杨燕宋乐王红李彦青高跳彭励
中国现代中药 2022年12期
李振凯,冯璐,杨燕,宋乐,王红,李彦青,高跳,彭励
宁夏大学 生命科学学院,宁夏 银川 750021
中药银柴胡是我国传统医学中常用的一种清虚热、除疳热的根类药材,临床中多用于治疗阴虚发热、骨蒸劳热、小儿疳热等证,现代医学发现银柴胡还具有抗炎、抗过敏、抗癌等功效,具有良好的药用价值[1]。20 世纪60 年代,银柴胡作为首批中药材入选《中华人民共和国药典》(以下简称《中国药典》)1963 年版[2],并一直收录至今,其基原植物为石竹科繁缕属植物银柴胡Stellaria dichotomaL.var.lanceolataBge.,至今无新的品种纳入。
宁夏为银柴胡的道地产区,自20世纪80年代引种驯化成功后,已有40 年的银柴胡种植历史。目前,宁夏同心县已建成了全国最大的银柴胡种植基地,获得了“同心银柴胡”国家农产品地理标志认证。但是在银柴胡由野生转为栽培的过程中,其生境和种植方式产生了显著的变化。尤其是农户长期自繁、自用,导致银柴胡种质出现不同程度的退化。调查中发现,部分产地植株出现形态矮化、叶片变厚、种子千粒质量下降、抗逆性变弱等现象。目前,生境的变化或栽培技术的不同是否引起了栽培银柴胡种质的改变尚未见报道。有关银柴胡伪品从古至今均有记载。清代《本草崇原集说》中记载:“今市肆中另觅草根白色而大,不知何种名银柴胡,此伪充也,不可用。”[3]在近现代临床用药和研究中,不仅发现霞草Gypsophila oldhamiana Miq.、旱麦瓶草Silene jenisseensisWilld、女娄菜Silene apricaTurcx.ex Fisch.et Mey.、灯芯草蚤缀Arenaria dahuricaFisch.ex Ser.、蝇子草Silene gallicaLinn.、石竹Dianthus chinensisL.等银柴胡近缘植物的根伪充银柴胡的现象,还有未定名银柴胡伪品的记录[4-5]。因此,实际栽培生产中银柴胡品种退化的现象是否与种质资源差异、伪品混入有关均有待于进一步验证。
中药材DNA 条形码是一种基于分子生物学的新型中药材鉴别方法[6-7],其鉴定的准确性和可靠性已被大量研究结果所证实。《中国药典》2015 年版起收录中药材DNA 条形码分子鉴定指导原则,成为鉴别药材真伪和保证中药材临床应用准确、安全、有效的技术手段。陈士林[8]通过对10 批银柴胡药材的核糖体DNA内转录间隔区2(ⅠTS2)序列进行分析,明确了以228 bp 的ⅠTS2 序列为主体的标准银柴胡DNA条形码,为银柴胡的分子鉴定提供了依据。
本研究以标准银柴胡DNA 条形码ⅠTS2 序列为鉴定依据,对收集的包括不同产地、不同生活型(野生、栽培)的17 份银柴胡和2 份伪品进行ⅠTS2序列分析,以期对不同种源银柴胡及伪品进行鉴别,为银柴胡种质资源多样性研究、良种繁育及从源头保证药材生产的安全有效提供参考。
1 材料
1.1 仪器
JXFSTPRP-CL 型全自动样品研磨仪(上海净信实业发展有限公司);MiniSpin plus 型高速离心机(德国Eppendorf公司);MⅠULAB型恒温金属浴(杭州米欧仪器有限公司);MyCycler 型聚合酶链式反应(PCR)仪(美国Bio-Rad公司)。
1.2 试药
银柴胡种子分别采集自不同产地银柴胡植株,经宁夏大学生命科学学院彭励教授鉴定为银柴胡Stellaria dichotomaL.var.lanceolataBge.的种子。其中1份银柴胡种子(S16)为航天诱变后的F1代种子。所有银柴胡种子于2021 年5 月种植于相同试验田中,期间采用相同的田间管理,在生长10 周后,取其叶片用于DNA 提取和测序分析。2 份伪品于2020 年5 月采集植株,经彭励教授鉴定为霞草Gypsophila oldhamianaMiq.和旱麦瓶草Silene jenisseensisWilld.,将其移栽至与种植银柴胡相同的试验田中,进行相同的田间管理,同银柴胡样品采集同一时期采集新鲜叶片用于DNA 提取和测序分析(表1)。通过《中国药典中药材DNA条形码标准序列》获得了银柴胡的标准序列[8];霞草、旱麦瓶草、女娄菜、灯芯草蚤缀、蝇子草和石竹的ⅠTS2 序列来源于GenBank数据库,序列信息见表2。
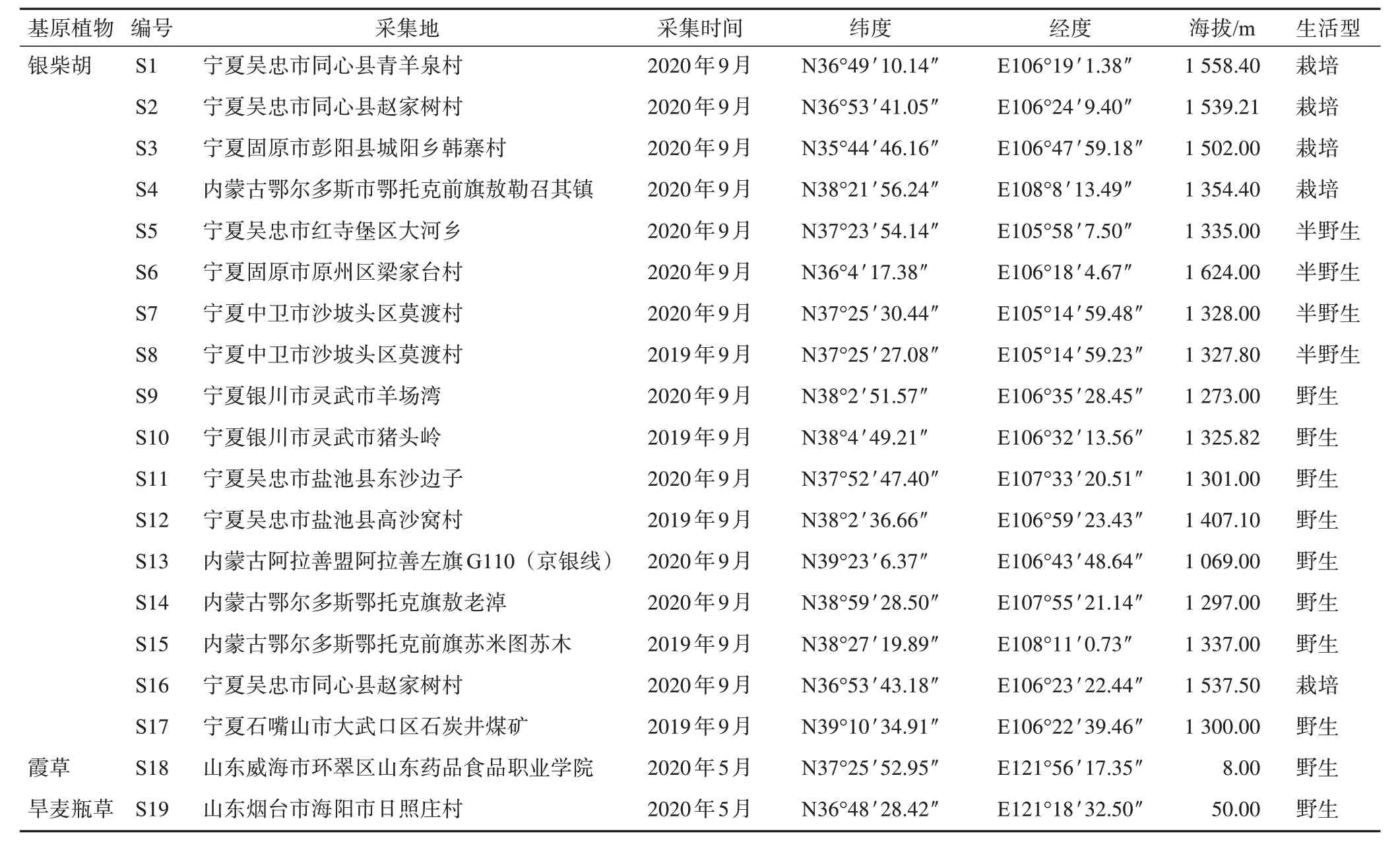
表1 银柴胡及其混伪品植物样品采集信息
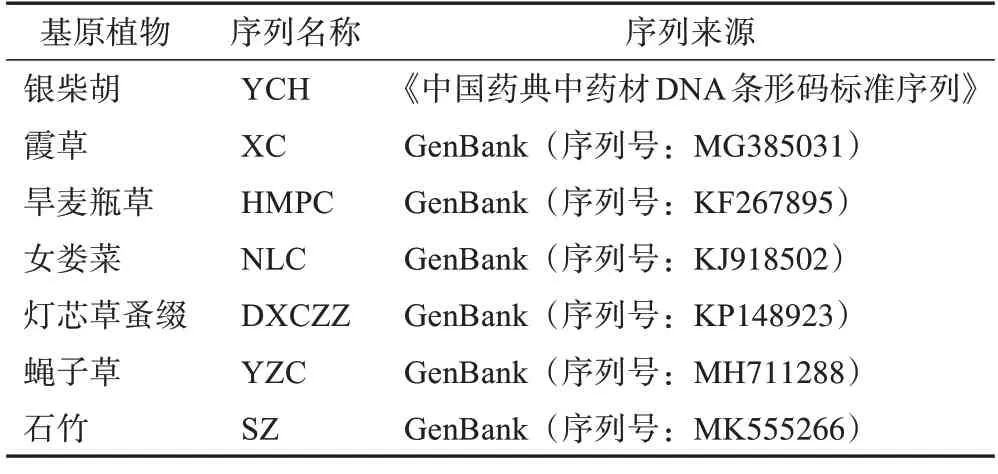
表2 银柴胡及其混伪品的ITS2检索序列信息
Ezup 柱式植物基因组DNA 抽提试剂盒 [批号:B518261,生工生物工程(上海)股份有限公司]。
2 方法
2.1 DNA的提取
随机从每份样品中取新鲜叶片1~2片,每个样品3个重复,分别加入液氮充分碾磨并离心,采用Ezup柱式植物基因组DNA抽提试剂盒提取总DNA。
2.2 PCR扩增及测序
采用《中国药典中药材DNA 条形码标准序列》中ⅠTS2 通用引物:正向引物为5′-ATGCGATACTTGGTGTGAAT-3′,反向引物为5′-GACGCTTCTCC-AGACTACAAT-3′。PCR反应体系(20 μL):dNTP mix(2.5 mmol·L-1)4.0 μL,正、反向引物(10 μmol·L-1)各2.0 μL,Phusion DNA 聚合酶(2 U·μL-1)0.5 μL,DNA 模板1.0 μL,5×Phusion HF Buffer 10 μL,双蒸水补足至20 μL。PCR 扩增条件:预变性98 ℃,3 min;变性98 ℃,10 s,退火58 ℃,30 s,延伸72 ℃,30 s,循环30 次;延伸72 ℃,8 min,16 ℃保存。将所得的DNA 扩增产物一并送生工生物工程(上海)股份有限公司进行DNA序列双向测定。
2.3 序列处理与分析
将所得序列一并导入ⅠTS2 数据库(http://its2.bioapps.biozentrum.uni-wuerzburg.de/)进行序列注释,去除两端(5.8S和28S rRNA)区段,得到完整的ⅠTS2 序列。利用MEGA 8.0 软件对所得序列进行数据分析,计算鸟嘌呤(G)+胞嘧啶(C)占比,选择邻接(neighbor-joining,NJ)法构建系统发育树,以Kimura 2-parameter(K2P)遗传距离模型计算得各样本遗传距离值。将得到的ⅠTS2 序列导入美国国家生物技术信息中心(NCBⅠ)数据库(https://blast.ncbi.nlm.nih.gov/Blast.cgi)进 行BLAST 分析,导入“中药材DNA 条形码鉴定系统”(http://59.108.50.222/china/index.php? optionid=174#this)进行物种鉴定。利用ⅠTS2数据库预测ⅠTS2序列的二级结构。
3 结果与分析
3.1 不同种源银柴胡ⅠTS2序列一级结构分析
通过对17份银柴胡样品的ⅠTS2序列进行测序分析,共得到3 种单倍型序列。如图1 所示,样品S1~S3、S5~S8、S10、S12~S16 拥有相同的ⅠTS2 序列单倍型,为方便描述,将该序列命名为YCH1;样品S4、S9、S11 的ⅠTS2 序列相同,命名为YCH2;样品S17 的ⅠTS2 序列与其他样品均不相同,命名为YCH3。其中YCH1 片段大小为228 bp,G+C 占比为57.46%;YCH2 片段大小为228 bp,G+C 占比为57.02%;YCH3 片段大小为224 bp,G+C 占比为55.36%。参照《中国药典中药材DNA 条形码标准序列》中公布的银柴胡标准ⅠTS2序列YCH,对所有序列进行比对分析。结果显示,YCH1和YCH2单倍型与YCH 一致,2 个单倍型存在的3 个变异位点均为YCH 公布的3 个变异位点,分别为89 bp处的C-胸腺嘧啶(T)变异位点、176 bp 处的T-C 变异位点和179 bp处的C-腺嘌呤(A)变异位点。YCH3单倍型与YCH比对含有4个碱基缺失,分别为61 bp处的C 缺失、102 bp 处的A 缺失、112 bp 处的C 缺失和172 bp 处的G缺失;另外在标准序列的3个变异位点上,89 bp位点为C、176 bp位点为T、179 bp位点为C,与YCH1单倍型的变异位点碱基相同。
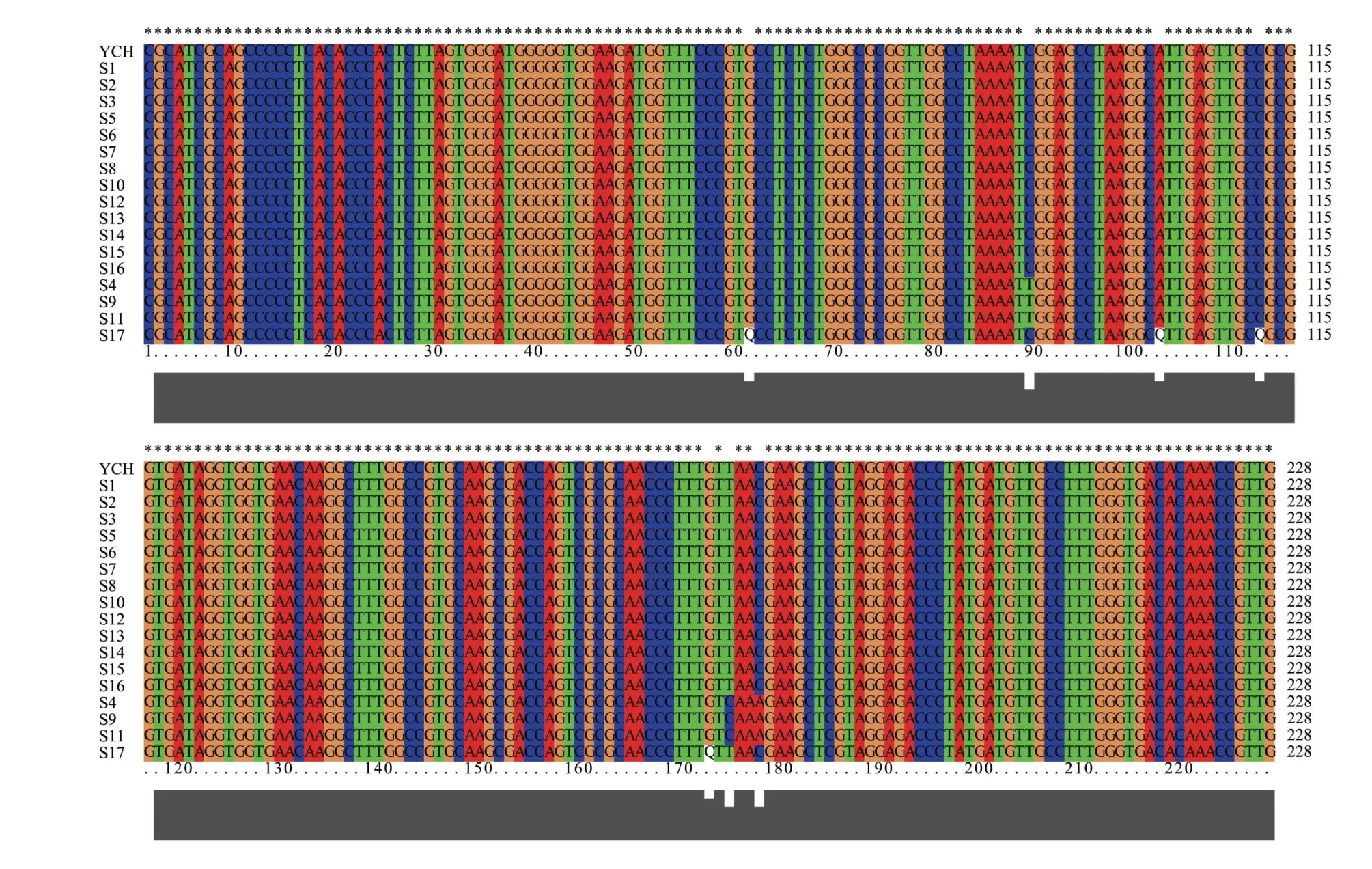
图1 17份银柴胡样品ITS2序列对比
3.2 银柴胡与伪品基原植物ⅠTS2序列对比分析
通过检索GenBank 数据库和《中国药典中药材DNA 条形码标准序列》获得了霞草、旱麦瓶草、女娄菜、灯芯草蚤缀、蝇子草和石竹等银柴胡伪品的6条ⅠTS2序列,分别为XC、HMPC、NLC、DXCZZ、YZC 和SZ,同时通过对霞草和旱麦瓶草的ⅠTS2 序列进行测序,得到2 条单倍型序列XC1 和HMPC1。进一步对银柴胡及6 种伪品ⅠTS2 序列的一级结构进行比对分析。结果如图2 和表3 所示,相较于3 种单倍型银柴胡ⅠTS2 序列与YCH 的差异,来自于6 种伪品基原植物的ⅠTS2 序列在序列长度、G+C 占比及变异位点上均存在较大的差异。

表3 不同单倍型银柴胡及伪品基原植物ITS2序列特征
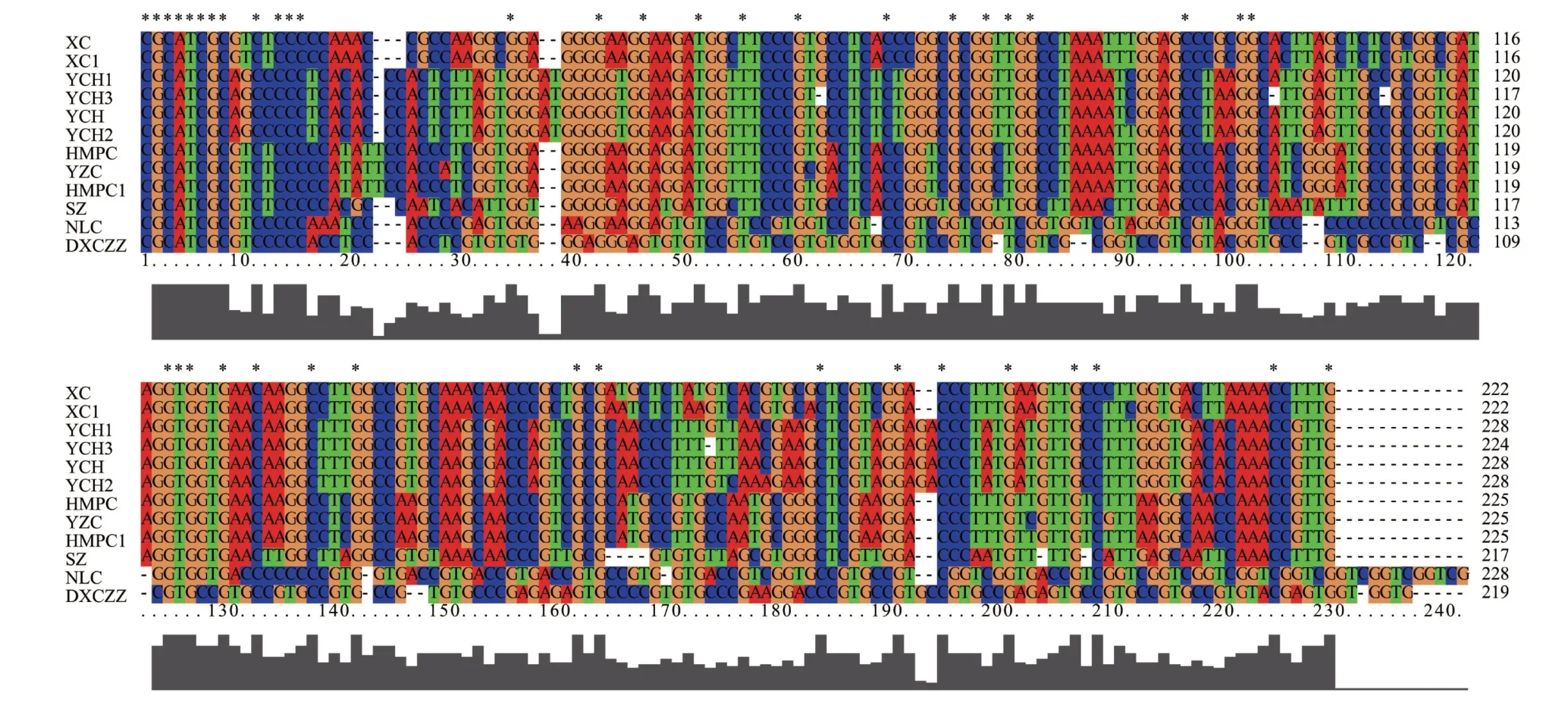
图2 不同单倍型银柴胡及伪品基原植物ITS2序列
将获得的3 种单倍型银柴胡ⅠTS2 序列和6 种伪品的ⅠTS2 序列导入NCBⅠ数据库,以YCH 为对照进行BLAST 分析,结果显示YCH1 与标准序列相似度为100%,YCH2 为98.68%,YCH3 为98.25%,而6 种伪品基原植物ⅠTS2 序列均未得到比对结果。进一步将以上序列分别导入“中药材DNA 条形码鉴定系统”进行物种鉴定,结果显示来自银柴胡的3 种单倍型序列均被鉴定为银柴胡,其中YCH1和YCH2序列与数据库中银柴胡ⅠTS2 相似性最高均为100%,YCH3 序列相似性最高为98.20%,其余6 种伪品基原植物ⅠTS2序列鉴定结果均非银柴胡。
3.3 遗传距离及系统发育树分析
基于K2P 模型计算不同单倍型银柴胡及伪品基原植物ⅠTS2 序列遗传距离,结果见表4。银柴胡的YCH1单倍型序列与YCH一致,遗传距离最小,为0;次之为YCH2 单倍型,遗传距离为0.014;YCH3 遗传距离最大,为0.936。而伪品基原植物ⅠTS2 序列与YCH 均表现出较大的遗传距离,其中旱麦瓶草和蝇子草ⅠTS2 序列(HMPC、HMPC1 和YZC)遗传距离相对较小,分别为1.610、1.621 和1.610,其余植物ⅠTS2 序列的遗传距离均超过1.800。进一步构建不同单倍型银柴胡及伪品基原植物ⅠTS2 序列的NJ系统发育树(图3)。银柴胡、旱麦瓶草和蝇子草聚为1 个大的分支,霞草、石竹、女娄菜和灯芯草蚤缀聚为1个大的分支,其中YCH1和YCH2单倍型序列与YCH聚为1个小的分支,又与YCH3同为1个分支,此外HMPC1 与HMPC、XC1 和XC 各聚为1 个分支。综上,基于K2P 模型计算的ⅠTS2 序列遗传距离和NJ系统发育树表现出一致的特征,说明不同单倍型银柴胡及伪品基原植物间存在不同的亲缘距离。
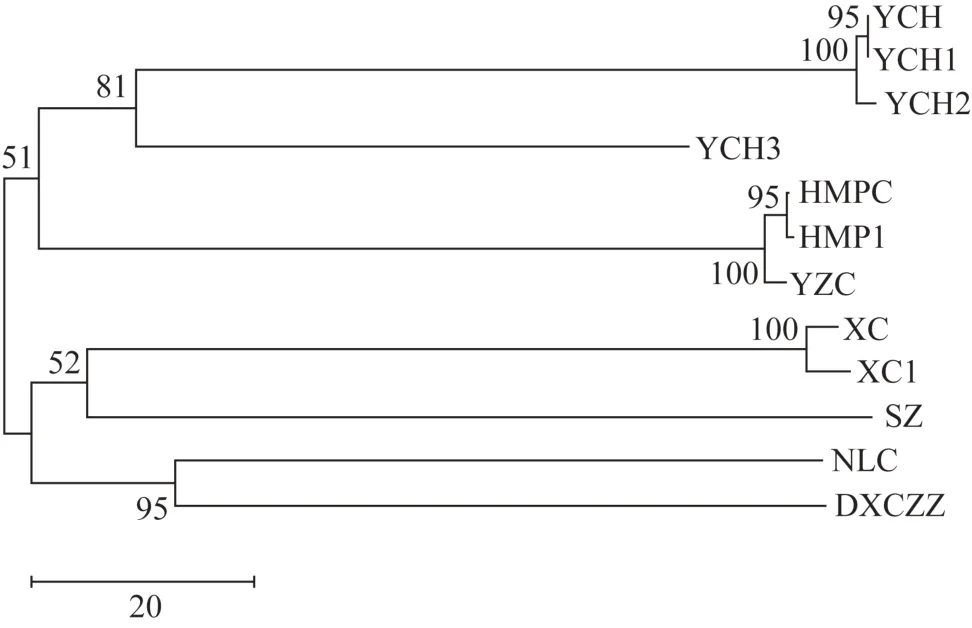
图3 不同单倍型银柴胡及伪品基原植物ITS2序列NJ系统发育树
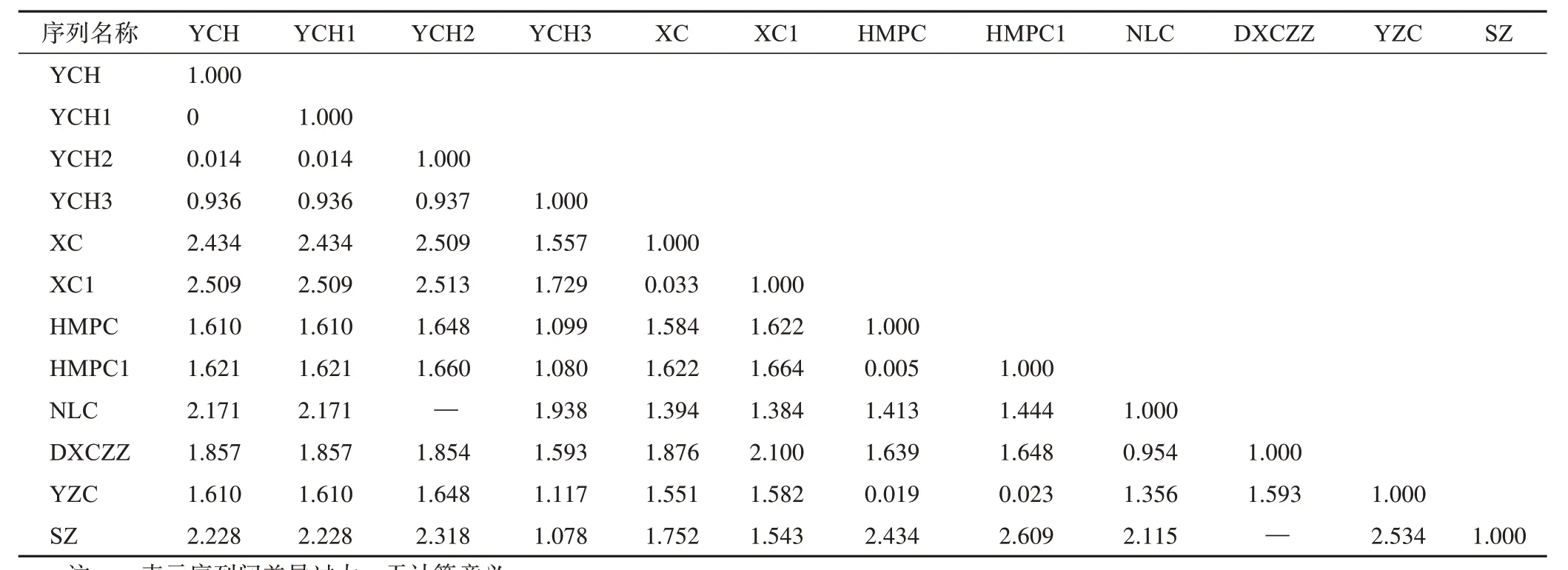
表4 不同单倍型银柴胡及伪品基原植物ITS2序列遗传距离
3.4 ⅠTS2二级结构分析
将得到的不同单倍型银柴胡及伪品的ⅠTS2 序列分别导入ⅠTS2 数据库,进行二级结构预测,成功获得了YCH1、YCH2、YCH3、XC、HMPC、YZC、SZ的ⅠTS2序列二级结构,未成功预测出女娄菜和灯芯草蚤缀可靠的二级结构。预测结果如图4 所示,7 个序列单倍型的ⅠTS2 二级结构均为典型的被子植物ⅠTS2 序列一环(主环)四臂(4 个螺旋区)结构,但是不同序列在部分茎臂和环上存在一定差异。其中与YCH 相似度最高的YCH1 和YCH2 单倍型具有完全一致的二级结构,而相似度略低的YCH3 单倍型的Ⅱ臂多1 个内环,在Ⅲ臂的中段少了1 个内环,多了1 个凸环。其余4 个伪品基原植物的ⅠTS2 序列与3 种单倍型银柴胡ⅠTS2 序列在中心环结构、茎臂长度及内环和凸环数量、结构上均表现出显著差异。
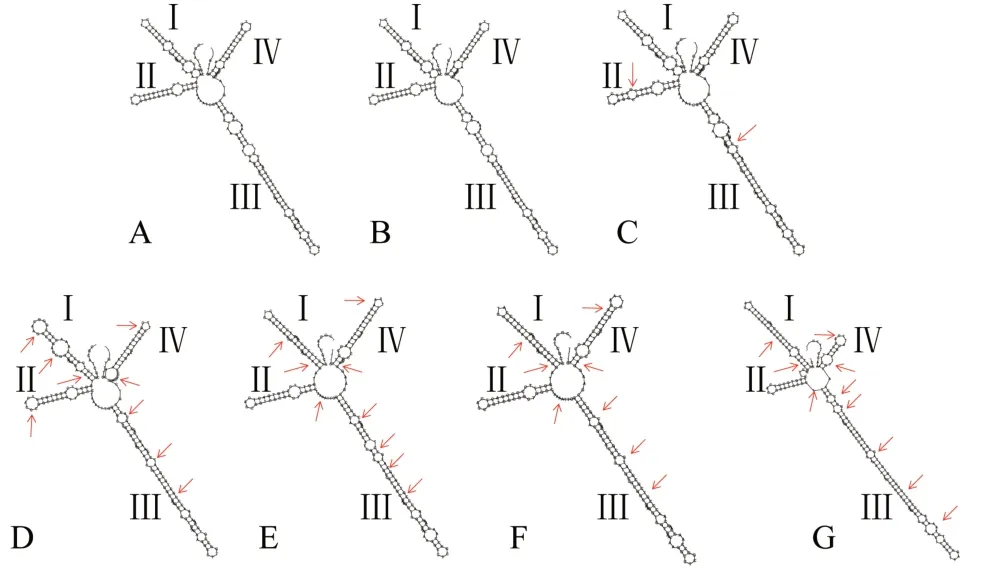
图4 不同单倍型银柴胡及伪品基原植物ITS2序列二级结构
4 讨论与结论
近年来,随着分子鉴定和DNA 条形码技术的不断成熟,ⅠTS2 序列在中药材尤其是植物类中药材鉴别分析中得到了广泛的应用和可靠的效果,并成为中药材DNA条形码建立的主体序列之一[7]。《中国药典中药材DNA 条形码标准序列》公布了包括银柴胡ⅠTS2序列DNA条形码在内的500余种中药材的DNA条形码,为中药材提供了可靠鉴定依据的同时,也为中草药基原植物、种子真伪鉴别和种质资源分析等相关研究提供了有效的分析手段。但在已有研究报道中,学者普遍关注银柴胡的ⅠTS 序列,对于ⅠTS2序列的应用和研究较少,如李军等[9]比较了4种DNA条形码序列ⅠTS、psbA-trnH、rbcL和matK在银柴胡药材及混伪品中的鉴别效果;孟祥善等[10]利用ⅠTS 序列分析了银柴胡种质资源的多样性。而ⅠTS2序列相较于ⅠTS 序列更易获得,承受的选择压力较小,相对变化较大,且具有相对保守的二级结构,在物种鉴定、遗传物质分析中具有更大的优势[11]。
本研究通过对17份不同种源银柴胡ⅠTS2序列进行测序和分析,共得到3条不同序列单倍型(YCH1,YCH2 和YCH3)。通过序列比对分析,发现表现为YCH1 或YCH2 单倍型的16 份银柴胡种质ⅠTS2 序列与标准银柴胡DNA 条形码ⅠTS2 序列高度一致。另有1 份样品为YCH3 单倍型,其ⅠTS2 序列与银柴胡标准序列的相似度达到98.20%,但其存在4 个碱基缺失,且与另外2 个单倍型具有较大的遗传距离(0.936~0.937)。此外,ⅠTS2 序列在细胞内以二级结构的形式发挥功能,这种结构蕴含着更为丰富的遗传信息[12]。进一步的二级结构分析显示,YCH1和YCH2 单倍型具有相同的ⅠTS2 序列结构,但YCH3单倍型存在多处的结构差异,说明YCH1 和YCH2可能具有相同的遗传信息和生物功能,但YCH3 可能存在一定的差异。YCH3 单倍型来源于宁夏石嘴山市大武口区石炭井煤矿采集的银柴胡植物样品,在本课题组前期的银柴胡资源调查中发现,该采集地与其他采集地的生境有显著的差异,表现为高含盐量(全盐质量分数为297.74 g·kg-1),pH为7.29,而非其他产地的碱性土壤,土壤质地以砂砾为主,与其他产地中砂土或粉砂土的生境也有显著的差异。大量的研究证实,植物长期生长在特定环境中,受环境中各种生态因子的综合影响可能引起植物DNA突变、染色体结构和数量畸变等遗传物质的改变,进而改变新陈代谢调控酶,使得次生代谢的产物发生变异[13-15]。因此是否有更多遗传物质的变化,亦或是有新品种的产生,以及这些变异是否会引起药材成分和功效的变化尚需进一步关注和深入研究。此外,对6 种伪品基原植物ⅠTS2 序列的鉴定分析也进一步证明其与正品银柴胡存在遗传物质的本质差异,其ⅠTS2 序列不仅在一级结构上与正品银柴胡不同,二级结构也与银柴胡存在显著差异,因此,可以采用ⅠTS2技术进行银柴胡及其伪品的鉴定。
道地药材是指经过中医临床的长期验证,具有特定的种质、特定的产区和加工方法,质优效佳、具有较高知名度的药材[16-17]。其形成过程往往是特定基因型在特定生境下次生代谢过程关键酶基因表达受到时空差异等复杂调控的结果[18]。因此,具有道地性的种质资源被认为是决定道地药材品质的关键因素[19-20]。通过本研究可以发现,目前宁夏栽培银柴胡与野生银柴胡保持着一致或十分接近的ⅠTS2 序列特征,初步可以判断其具有相似的种质特性,为宁夏银柴胡道地性形成机制研究提供了有效的种质资源依据。此外,田间生产中出现的品种退化现象可能是田间管理方法或生境差异导致,因此建议进一步建立健全规范的银柴胡良种繁育技术,制定相关的操作规程。本研究结果对银柴胡真伪鉴别、种质资源筛选、新品种选育及宁夏银柴胡道地产区建设有积极的借鉴意义。
猜你喜欢
少年文艺·开心阅读作文(2021年8期)2021-09-05
世界科学技术-中医药现代化(2021年10期)2021-03-02
世界科学技术-中医药现代化(2020年2期)2020-07-25
小学科学(学生版)(2019年5期)2019-05-21
少儿美术(快乐历史地理)(2019年11期)2019-04-20
天然产物研究与开发(2018年7期)2018-08-21
中成药(2017年9期)2017-12-19
中国中药杂志(2017年13期)2017-07-31
中国调味品(2017年2期)2017-03-20
中国钱币(2016年4期)2016-05-17
