
基于Ecopath 模型的七连屿礁栖性生物的生态承载力分析
2022-02-17 09:40洪小帆陈作志张俊江艳娥龚玉艳蔡研聪杨玉滔
热带海洋学报 2022年1期
洪小帆, 陈作志, 张俊, 江艳娥, 龚玉艳, 蔡研聪, 杨玉滔
1. 中国水产科学研究院南海水产研究所, 农业农村部外海渔业开发重点实验室, 广东省渔业生态环境重点实验室, 广东 广州510300;
2. 南方海洋科学与工程广东省实验室(广州), 广东 广州 511458;
3. 上海海洋大学海洋科学学院, 上海 201306
中国南海位于具有世界最高生物多样性的“珊瑚金三角”区域的北缘, 珊瑚礁海域分布面积广阔,具有丰富的珊瑚礁生物资源(黄晖 等, 2013)。近年来, 由于环境变化和人类活动的影响, 不仅导致西沙群岛七连屿珊瑚礁出现珊瑚白化、覆盖度下降等问题, 礁栖性鱼类数量及密度也呈下降趋势, 甚至部分珊瑚礁鱼类出现了小型化和性早熟的现象, 严重损害了珊瑚礁生态系统的健康和食物产出功能(洪小帆 等, 2020; 张俊 等, 2020)。2006 年国务院颁发了《中国水生生物资源养护行动纲要》, 把水生生物增殖放流作为养护水生生物资源, 提高生物多样性和生态系统健康的重要措施之一。开展珊瑚礁增殖放流工作, 不仅可以有效提升珊瑚礁的生物量和种群结构, 部分固礁或护礁生物(砗磲等双壳类)的增加也有助于珊瑚礁的修复, 提高生态系统的健康水平。然而, 增殖放流不仅要恢复所放流物种的种群数量, 也要求保证不破坏放流水域的生态系统,维持生态系统的平衡。因此, 生态承载力的评估是科学实施增殖放流的基础和前提。
Ecopath 模型依据热力学守恒定律作为运行原理, 通过构建食物网对生态系统进行简化, 以建模的方式量化生态系统的各类特征参数, 反映某一时期特定生态系统的特征和营养关系(仝龄, 1999), 其不仅能够显示食物网物质平衡的静态状况, 同时也是作为生态系统动态模型的一个起始点(Christensen et al, 1992)。Ecopath 模型不仅能用以评估生态系统的结构、功能以及能量流动等特征, 同时也具有在维持生态系统平衡的条件下评估模型中功能组生态容量的功能, 并被广泛应用于渔业资源增殖承载力或生态容量评估(Xu et al, 2011; 黄梦仪 等, 2019)。因此, 本文以2019 年在七连屿海域开展的渔业资源和生态环境调查数据, 运用 Ecopath with Ecosim(EwE) 6.5 软件构建七连屿珊瑚礁生态系统的Ecopath 模型, 分析七连屿珊瑚礁生态系统的结构和功能, 评估七连屿珊瑚礁生态系统中部分功能组的生态承载力, 以及当前条件下珊瑚礁生态系统中各群落的增殖潜力, 旨在为南海珊瑚礁生态系统的健康和渔业可持续产出管理提供理论依据。
1 材料和方法
1.1 研究区域
七连屿位于中国南海宣德群岛(隶属西沙群岛)的东北部(16°55′~17°00′N, 112°12′~112°21′E) (图1),由若干个岛礁和沙洲组成, 岛屿和沙洲陆地总面积超过 1.32km2, 礁盘总面积约为 25km2(周晓刚,2012)。遥感分析的结果显示七连屿中北岛所在的礁盘的大部分区域水深不过超过20m(张靖宇, 2015);七连屿海域年平均表层海水温度为26.8℃, 受热带季风气候影响, 灾害性天气频发, 降水多集中于夏秋季节(5~11 月)。潮汐属于不正规全日潮, 海流以风生流为主, 流向随季风风向而变化。七连屿岛礁及邻近海域拥有鱼类、甲壳类、爬行类和藻类等丰富的生物资源, 是中国现存最大的绿海龟(Chelonia mydas)产卵场(张婷 等, 2020), 鱼类的物种多样性较高且生物分类学分布较为合理(李媛洁 等, 2020),同时也是我国珊瑚礁渔业的传统渔场, 具有重要的社会经济和生态环境保护价值。
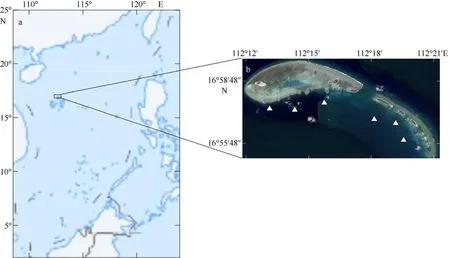
图1 调查区域及站点位置图a 基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2016)1665 号的标准地图制作; 图b 中白色三角为生物采样站点Fig. 1 The study area and the locations of the stations. White triangle markers are the stations of sampling
1.2 数据来源
本文所采用的基础数据来源于“南锋号”调查船于2019 年春季(2019 年5 月)和秋季(2019 年9 月)航次在七连屿珊瑚礁海域开展的渔业资源和生态环境调查。调查内容包括游泳动物、浮游生物、底栖生物以及其他环境要素采样, 采样及调查方法根据《海洋监测规范》(GB17378-1998)和《海洋调查规范》(GB-12763.91)进行。生物样本带回实验室鉴定和分析。
1.3 研究方法
利用EwE 6.5 软件基于七连屿周边礁盘构建珊瑚礁生态系统Ecopath 模型(Christensen et al, 2005)。在Ecopath 模型中, 需要综合考虑具有相似营养级、生活史和生态位特征的功能组(Pauly et al, 2000),且所有的功能组必须能够代表生态系统中能量和物质流动的基本过程(Polovina, 1983; Christensen et al,1992)。Ecopath 主要由两个方程构成, 方程(1)定义每个功能组的输入与输出保持平衡, 确保生态营养效率不超过1; 方程(2)定义功能组的热力学守恒。方程(1)、(2)表达式如下:
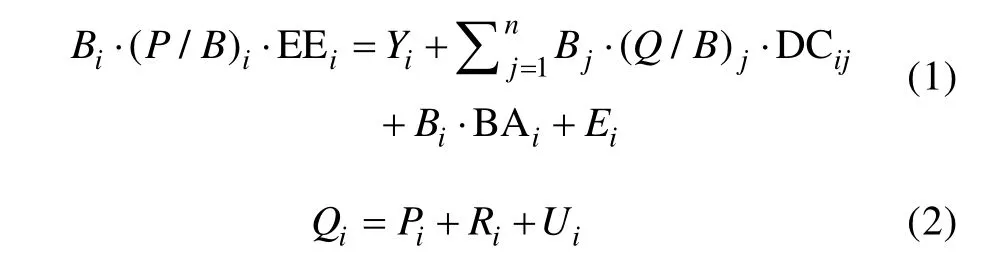
式中,Bi代表功能组i的生物量, (P/B)i代表功能组i的生产量与生物量比例, EEi代表功能组i生态营养级效率,Yi代表功能组i的渔获量, (Q/B)i代表功能组i消费量与生物量比例, DCij代表功能组i占功能组j食物构成的比例, BAi代表功能组i的生物累积率,Ei代表功能组i的净迁移量,Qi代表功能组i的消费量,Pi代表功能组i的生产量,Ri代表功能组i的呼吸量,Ui代表功能组i的未消化食物量(Christensen et al, 2008)。
1.4 功能组划分
模型功能组的划分须要充分考虑不同物种的栖息生境和食性随生活史变化而产生的转变, 功能组可由单个物种或多个物种组成(Walters et al, 2008)。根据珊瑚礁生态系统中生物的生态学、生物分类学或食性组成特征, 以及七连屿珊瑚礁海域物种分布的基本情况(孙典荣 等, 2005; 李元超 等, 2017; 李媛洁 等, 2020), 同时结合其他具有相似生态系统特征的珊瑚礁海域模型, 将七连屿珊瑚礁生态系统模型划分为21 个功能组, 基本覆盖该生态系统各营养级的能流流动过程。模型功能组包括初级生产者[浮游植物、小型底栖藻类、大型底栖藻类、碎屑(有机物质颗粒)]、珊瑚礁鱼类(杂食性鱼类、珊瑚食性鱼类、植食性鱼类、小型肉食性鱼类、中型肉食性鱼类、大型肉食性鱼类)、大型海洋动物(软骨鱼类、海龟)、无脊椎动物[浮游动物、甲壳类、双壳类、珊瑚、小型底栖动物、棘冠海星(Acanthaster planci)、法螺(Charonia tritonis)、其他棘皮动物、其他软体动物](表1)。

表1 七连屿珊瑚礁海域生态系统功能组及主要种类Tab. 1 Functional groups and main species checklist based on the Ecopath model in Qilianyu Islands
1.5 功能组参数来源
本文Ecopath 模型中, 系统中能量的流动以生物量形式(生物湿重, 单位为t·km–2)进行表示, 时间限制为1a。生物量主要通过调查数据进行估算, 其中浮游植物生物量根据南海岛礁海域叶绿素a浓度进行换算(Ke et al, 2018); 珊瑚礁鱼类采用地笼网、手钓等作业方式进行采集, 并结合声学探测技术估算各珊瑚礁鱼类功能组生物量(Zhang et al, 2016);珊瑚、棘冠海星和法螺的生物量因缺少相关实地调查数据, 故参考其他珊瑚礁生态系统的模型参数设置而提前设定EE 值进行推算; 有机碎屑生物量根据Paulya 等 (1993)提出的经验公式进行计算得出;浮游动物、甲壳类、小型底栖动物、软骨鱼类、棘皮动物和软体动物等功能群的生物量参考周边海域的相关研究调查结果。鱼类功能组的P/B值与Q/B值分别根据Pauly(1980)和Palomares 等 (1998)提出的经验公式进行推导, 同时以与七连屿珊瑚礁海域纬度相近或生态系统特征相似的模型作为主要参考对象, 并且根据实际情况进行相应的调整。食物组成矩阵 (Diet composition)通过结合胃含物分析与Fishbase (https://www.fishbase.org)的历史记录完成构建。渔获量统计数据根据渔民提交的生产数据以及通过走访调查和座谈得到的相关信息进行综合估算得出。
1.6 Ecopath 模型的调试与敏感性分析
本文采用PREBAL 预检验对B、P/B、Q/B和P/Q(P/B与Q/B的比值)进行模型平衡前检验, 以检查模型的一致性(Link, 2010), 检验的指标包括: 生物量分布、生物量比值、“重要比例”分布、“重要比例”比值、整体的生产和消耗。根据质量守恒和热力学守恒原则, 对预检验之后的Ecopath 模型通过一系列逻辑约束进行调整, 以达到生态学和热力学平衡状态(Heymans et al, 2016), 需要调平的参数如表2 所示。

表2 模型平衡的生态学和热力学原则Tab. 2 Ecological and thermodynamic principles of model balance
应用敏感性分析以检测模型各功能组基本输入参数的细微变化对输出参数的影响水平, 输出参数对于输入参数的敏感性取决于功能组之间的关联程度。拟定基本参数(B或EE)以10%的步长发生变化,改动范围为±50%, 以测试输出参数对于输入参数不同波动变化的响应幅度。敏感性分析的计算方式为: (估算参数-原始参数)/原始参数(Christensen et al,2005)。
1.7 生态承载力评估
本文参考生态承载力的相关研究(Christensen et al, 1998; McKindsey et al, 2006), 并根据研究的主要任务, 将生态承载力(Ecological carrying capacity)定义为在未显著改变生态系统的主要能量流动和食物网结构的前提下, 生态系统所能承受的目标种增殖或衰减的最大承重水平(刘岩 等, 2019)。根据Ecopath 模型原理, 功能组生物量的变化均会对其自身EE 以及与其具有密切营养联系的功能组参数产生影响。
本文结合近年来有关七连屿珊瑚礁生态系统的相关调查和研究结果, 从生态和经济方面进行综合考虑, 对七连屿珊瑚礁生态系统中具有护礁和固礁作用的双壳类和珊瑚, 以及具有重要经济价值的法螺、大型肉食性鱼类和植食性鱼类等5 个功能组作为目标功能组进行生态承载力推算; 在模型调试平衡之后, 通过10%的步长比例逐步增加或减少模型中目标功能组的生物量, 在反复迭代模拟的过程中直至使模型中任意功能组首次出现EE≥1 时, 则确定此时目标功能组的生物量为生态系统对目标功能组的生态承载力。此外, 根据各功能组的生物学特征将模型中大部分功能组归类为两个主要珊瑚礁生物群落——珊瑚礁鱼类群落(杂食性鱼类、珊瑚食性鱼类、植食性鱼类、小型肉食性鱼类、中型肉食性鱼类、大型肉食性鱼类)和珊瑚礁无脊椎动物群落(浮游动物、甲壳类、双壳类、珊瑚、小型底栖动物、其他棘皮动物、其他软体动物), 分别以10%的步长比例逐步增加各群落的总体生物量(群落内各功能组的生物量按相同比例增加), 分析不同增殖模式下珊瑚礁鱼类群落的最大生态承载力。
2 结果
2.1 Ecopath 模型的数据敏感性分析
敏感性分析显示, 各功能组的EE 与生物量的数值变化关系最为敏感(图2), 相较于其他敏感性关系, 浮游植物功能组的EE 数值对其B数值减少时的敏感性最高, 而其他软体动物功能组的EE 则对其B数值增加时的敏感性最高; 软骨鱼类功能组的EE数值对其B数值的变化没有任何敏感性响应。整体分析显示, 当各功能组输入参数数值的变化率为–50%时, 模型输出参数的平均响应幅度达到最大(91.52%); 当各功能组输入参数数值的变化率在0~50%时, 则模型输出参数的平均响应幅度较小。
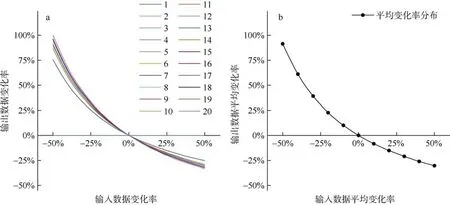
图2 七连屿珊瑚礁生态系统Ecopath 模型敏感性分析结果图a 为各功能组的敏感性分析结果, 功能组序号与表1 相对应; 图b 为模型输出数据的平均变化率Fig. 2 Sensitivity analysis of Ecoapth model for coral reef ecosystem of Qilianyu Islands. a: Parameter sensitivity of each group; b: average response of each parameter
2.2 营养结构和能流分布
七连屿珊瑚礁海域Ecopath 模型的基本输入和调试输出结果如表3 所示, 各功能组营养级(Trophic level, TL)符合生态学基本规律, 营养级范围在1.00~3.81 之间。其中, 软骨鱼类具有最高营养级(3.81), 其次为大型肉食性鱼类(3.60); 珊瑚礁鱼类功能组营养级范围为2.18~3.60, 平均营养级为2.97。
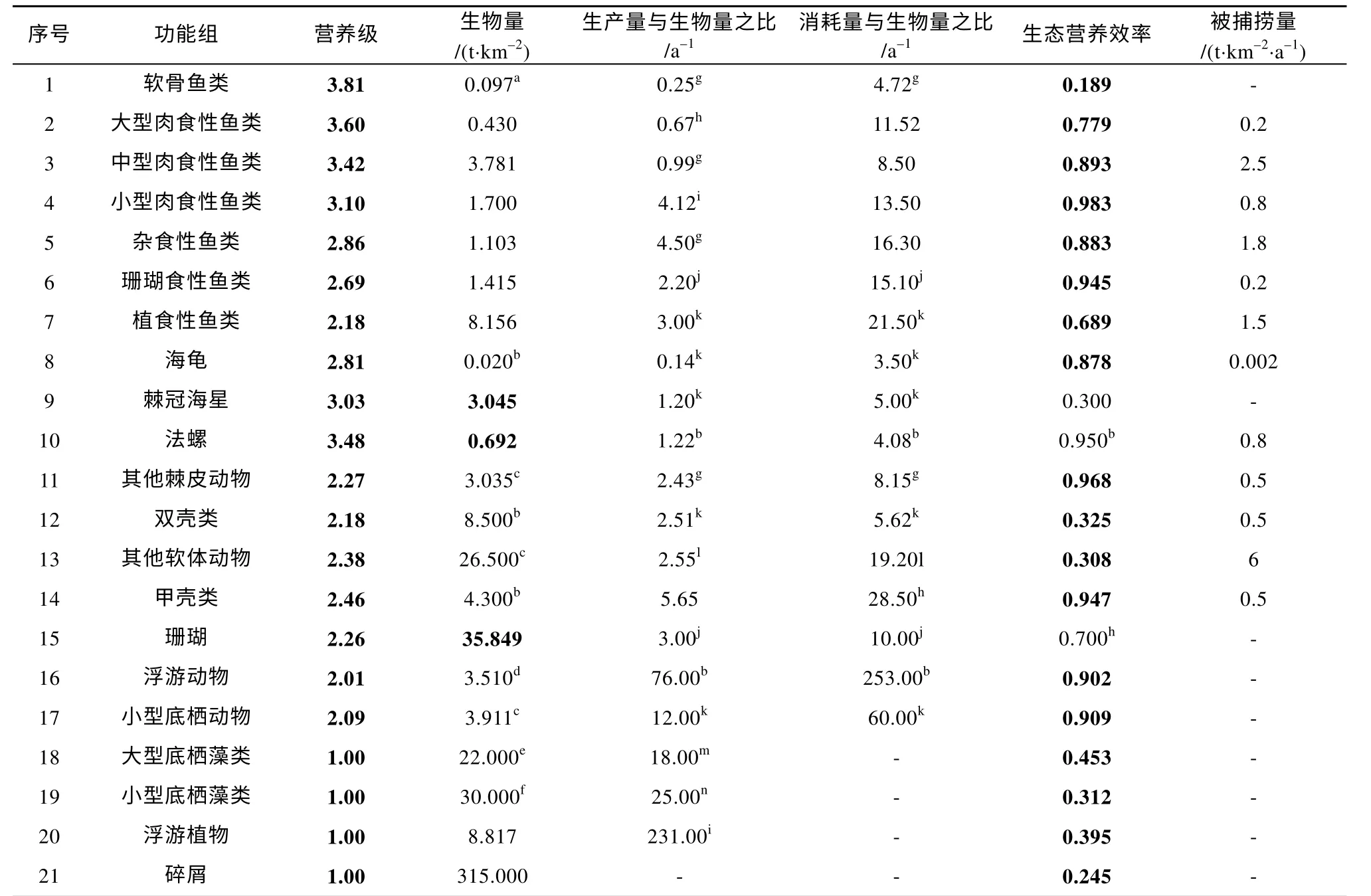
表3 七连屿珊瑚礁生态系统Ecopath 模型功能组估算参数Tab. 3 Basic input data and estimated parameters of the Ecopath model in Qilianyu Islands
通过营养级聚合将七连屿珊瑚礁海域生态系统21 个功能组的营养流合并为4 个整合营养级(表4),各营养级的总流量随营养级的升高而显著降低, 营养级Ⅰ和Ⅱ的总流量分别占各级总流量总和的70.67%和 24.00%, 营养级Ⅲ和Ⅳ则共占总和的5.34%, 基本符合生态学能量金字塔规律。作为生态系统能量和物质的主要来源, 营养级Ⅰ被捕食量占各级被捕食量总和的81.88%, 其流向碎屑量占各级流向碎屑量总量的71.71%(表4)。
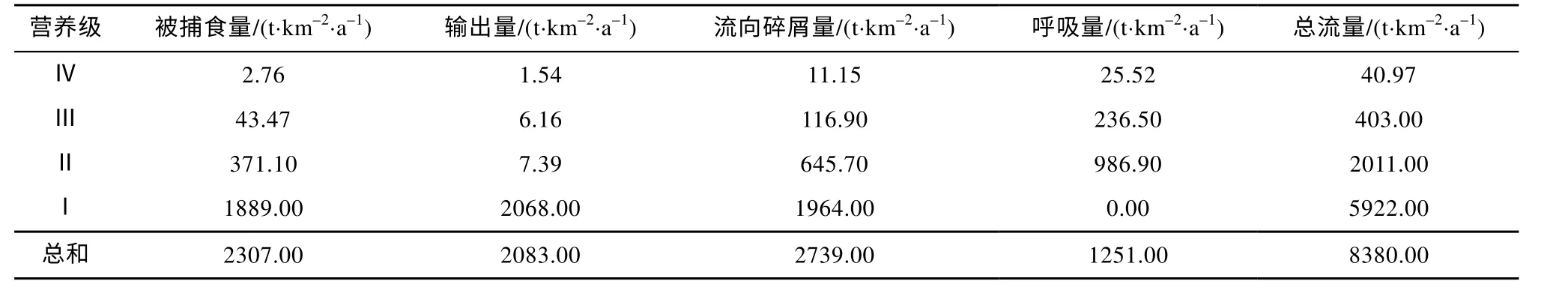
表4 七连屿珊瑚礁生态系统总能流的分布Tab. 4 Distribution of energy flows at aggregated trophic levels in Qilianyu Islands ecosystem
2.3 营养转化效率
根据生态系统中能量来源的差异, 本文的生态系统模型中食物网主要由牧食食物链和碎屑食物链所构成。由表5 可知, 在营养级Ⅱ~Ⅳ的范围中, 七连屿珊瑚礁生态系统总体的平均转化效率为13.45%;营养级Ⅱ具有最高的转化效率(18.82%), 而能流来源于初级生产者的营养级Ⅳ的转换效率最低, 为10.22%, 来自初级生产者和碎屑的平均转化效率分别为13.37%和13.58%。此外, 在七连屿珊瑚礁海域生态系统能量流动的来源中, 直接来源于碎屑的占总体能量流动的43%, 来源于初级生产者的占比则为56%, 显示当前七连屿珊瑚礁海域生态系统主要以牧食食物链为主导。
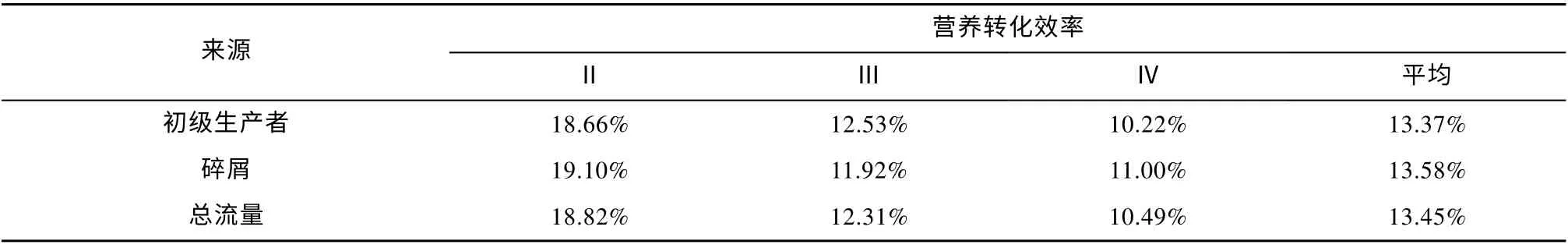
表5 七连屿珊瑚礁海域生态系统营养级的转化效率Tab. 5 Transfer efficiency of discrete trophic levels in Qilianyu Islands ecosystem
2.4 混合营养效应分析
在Ecopath 模型中, 通过混合营养分析以评估生态系统各功能组之间的相互关系, 某一功能组对另一功能组的消极效应代表两者具有相同的食性或后者受到前者的捕食作用; 某一功能组对另一功能组的积极效应则代表两者没有捕食竞争关系或前者为后者所捕食。本文模型分析显示初级生产者对大部分功能组均具有积极效应; 珊瑚对于珊瑚食性鱼类和棘冠海星具有显著的积极效应; 捕捞活动对大型肉食性鱼类、中型肉食性鱼类、法螺和海龟等功能组均具有明显的消极效应, 对于小型肉食性鱼类、珊瑚食性鱼类、植食性鱼类和棘冠海星等功能组则具有积极效应(图3)。

图3 七连屿珊瑚礁生态系统混合营养关系图Fig. 3 Mixed trophic impact in coral reef ecosystem of Qilianyu Islands
2.5 生态承载力评估
七连屿珊瑚礁生态系统中双壳类、珊瑚、法螺、大型肉食性鱼类和植食性鱼类等6 个功能组的生态承载力推算结果如表6 所示。其中, 双壳类功能组的生物量为8.5t·km–2, 当其生物量增加到原始生物量的4.7 倍或减少至原始生物量的0.3 倍时, 系统的平衡状态被打破, 双壳类功能组的EE 均超过1, 显示七连屿珊瑚礁生态系统对于双壳类功能组的生态承载力大约在2.55~39.95t·km–2之间。在现有捕捞条件下, 法螺和大型肉食性鱼类的原始生物量均接近其自身在生态系统中生态承载力的最小值,而植食性鱼类则具有较高的未利用的生态承载力空间(表6)。
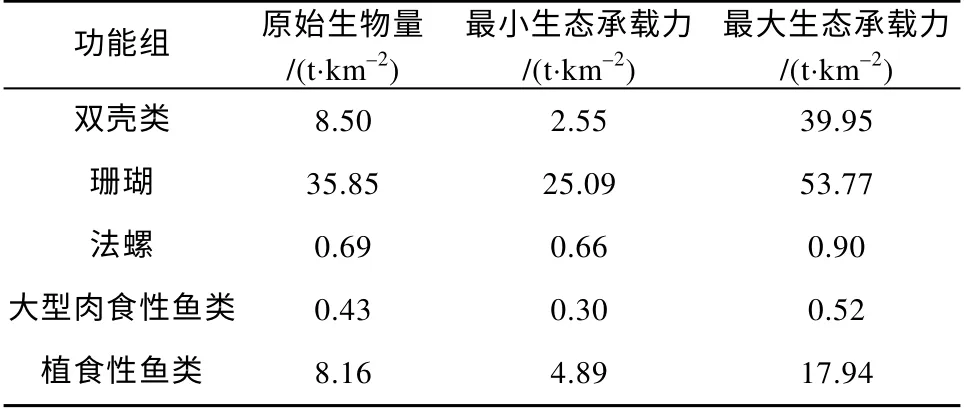
表6 七连屿珊瑚礁生态系统中部分功能组的生态承载力Tab. 6 Carrying capacity of some functional groups in Qilianyu Islands ecosystem
根据Ecopath 模型中进行群落生物量的增殖模拟发现, 在当前模型其他条件不变的情况下, 珊瑚礁鱼类群落的生物量整体上并无继续增殖的潜力;珊瑚礁无脊椎动物群落的生物量在其他条件不变的情况下则具有约1.9 倍的增殖空间(达到最大生态承载力); 在此基础上, 珊瑚礁鱼类群落在不破坏生态系统模型平衡的前提之下, 整体生物量可扩容2.1倍。此外, 当模型中珊瑚礁鱼类群落和珊瑚礁无脊椎动物群落的生物量均扩大到2.2 倍时, 两个群落的整体生物量均达到其最大生态承载力(表7)。
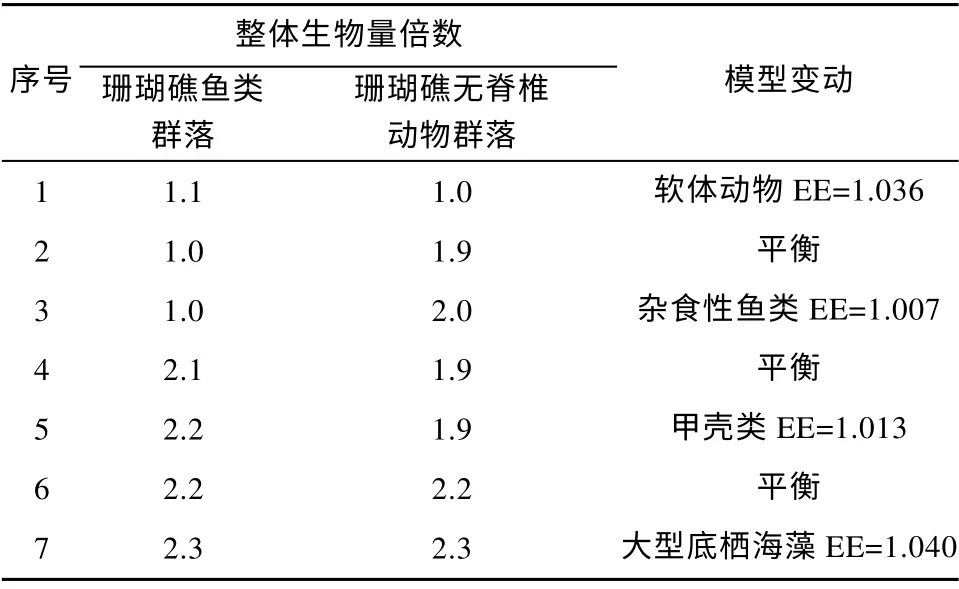
表7 七连屿珊瑚礁生态系统中各群落生物量增殖后的变动情况Tab. 7 Changes in the Qilianyu Islands Ecopath model after proliferating biomass of communities
2.6 生态系统的总体特征
七连屿珊瑚礁生态系统当前状态和两种群落增殖模式的总体特征如表8 所示。通过对比发现, 在两种群落增殖模式中, 系统总流量、总消耗量和总呼吸量均随模拟生物量的升高而升高。作为表征系统成熟度的关键指标, “数值3”模式下系统的总初级生产量与总呼吸量之比(TPP/TR)和总初级生产量与总生物量之比(TPP/TB)均为最低, 分别为 1.17和11.18; 3 个模型中系统联结指数(CI)和系统杂食指数(SOI)均无明显差异; Finn 循环指数(FCI)和Finn 平均路径长度(MPL)则均以当前系统为最低;且当前系统中系统权势占系统容量的 24.34%, 显著高于其他模拟结果。
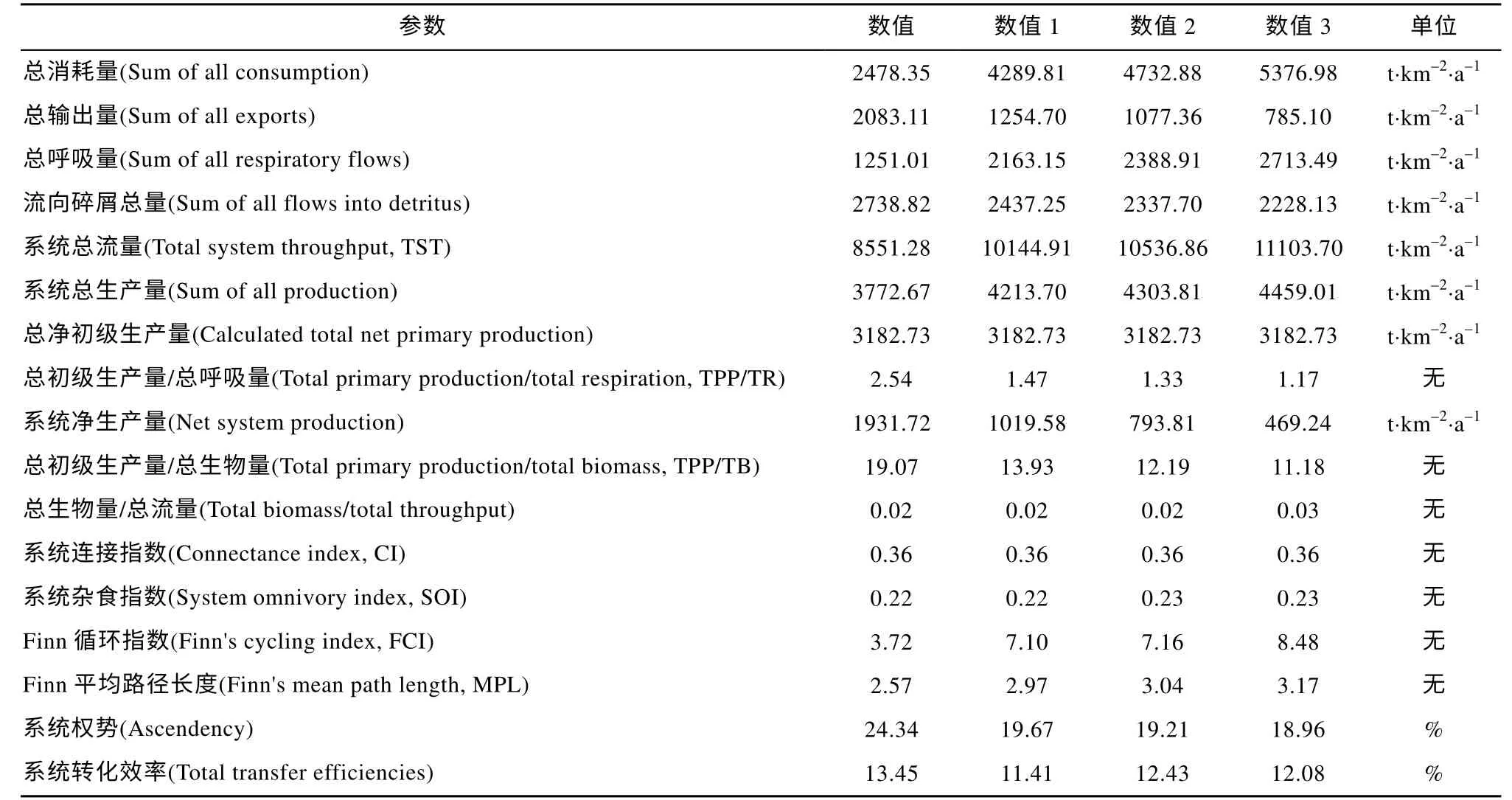
表8 七连屿珊瑚礁生态系统总特征参数表Tab. 8 Ecological system characteristic parameters of Qilianyu Islands ecosystem
3 讨论
本文通过构建七连屿珊瑚礁生态系统Ecopath模型, 定量分析了七连屿珊瑚礁生态系统的营养级结构和能流过程。Ecopath模型的Pedigree指数为0.33,对比Morissette 等(2006)对全球范围内150 个不同生态系统 Ecopath 模型的质量评价结果(Pedigree:0.16~0.68), 表明本文模型具有一定的可信度。在七连屿珊瑚礁生态系统的营养结构中, 软骨鱼类作为顶级捕食者, 其营养级仅为3.80, 与部分珊瑚礁生态模型的结果相似(Wabnitz et al, 2010; Fourriére et al,2019; Du et al, 2020), 这可能与软骨鱼类在海洋中具有较强的游泳能力且并未以珊瑚礁生境为其主要觅食区域等原因密切相关。此外, 作为目标渔获物的珊瑚礁鱼类的平均营养级(2.97)高于渔获物平均营养级(2.69), 显示七连屿珊瑚礁渔业中可利用的高营养级游泳动物种类较多。尽管不同生态系统类型具有不同的生态特征(Heymans et al, 2011), 但平均渔获物营养级仍可用以反映生态系统之间渔业保护的主要差异(Albouy et al, 2010)。本文模型的渔获物平均营养级不仅低于大多数珊瑚礁生态系统(Bozec et al, 2004;Cáceres et al, 2016; Ruiz et al, 2016; Fourriére et al,2019), 同时也低于南海北部大陆架海域的生态系统(2.93) (陈作志 等, 2010)。分析认为, 一方面, 珊瑚礁覆盖度下降所导致的环境变化会使得珊瑚礁生态系统内高营养级生物的丰度减少(Ainsworth et al, 2015);另一方面, 传统渔获对象的渔业资源在捕捞活动的作用下不断减少, 进而导致渔获物的平均营养级水平向下移动(Pauly et al, 1998)。
七连屿珊瑚礁生态系统的总能流转化效率为13.45%, 显著高于林德曼效率(10%)(Lindeman,1942); 其中营养级Ⅰ到营养级Ⅱ的转化效率最高,显示低营养级在能量转化过程中具有重要作用。尽管在总体能流途径中56%的总能流通过牧食食物链流动, 但是系统内部对于初级生产者(营养级Ⅰ)的能量利用仍较不充分, 来自初级生产者的能流占系统内各营养级流向碎屑的总量的71.71%。这表明七连屿珊瑚礁生态系统与典型珊瑚礁生态系统一样具有高初级生产力的特征, 但大部分能量都以碎屑的形式存储于生态系统低营养级中, 从而无法向上传递进入更高层次的营养流动。
在Ecopath 模型的输出结果中, 用以表征系统规模大小、稳定性、成熟性、发育状态等的参数, 是评价生态系统总体特征的主要指标。其中, TPP/TR和TPP/TB 作为表征生态系统成熟度的重要指标,TPP/TR 数值越接近1 则代表系统的稳定程度越高(Abdul et al, 2016), 当前模型所估算的TPP/TR 为2.54, 显示七连屿珊瑚礁生态系统中有机物质增加导致总初级生产量超过总呼吸量; 而本文模型所估算的TPP/TB 则为19.07, 显示生态系统向资源累积的方向发展, 这两个指标均反映当前七连屿珊瑚礁生态系统处于“稳定性较差”的未成熟状态。此外,FCI 和MPL 分别表示生态系统中再循环能流占系统总能流的比重以及各循环流经食物链的平均长度,两者的数值均随生态系统成熟度的增加而增大(Christensen et al, 2008)。当前模型中FCI 为3.72、MPL 为2.57, 与其他增殖模式的Ecopath 模型相比,这两个参数显示当前系统中被用以物质和能量再循环的生产力比例较低, 以及能量流动在系统中流经的营养级路径较短。结合当前模型中远小于1 的CI和SOI 值(用以表征系统内部联结复杂性的指标),表明七连屿珊瑚礁生态系统的食物网结构相对单一,以及系统成熟度和稳定性较低。
根据Ecopath 模型估算, 当前七连屿珊瑚礁中珊 瑚 和 法 螺 的 生 物 量 分 别 为 35.85t·km–2和0.69t·km–2, 二者的生态承载力区间分别为 25.09~53.77t·km–2和0.66~0.90t·km–2, 这显示珊瑚在现有环境条件下仍具有较大的增殖空间。法螺作为棘冠海星的少数天敌之一(Zhang et al, 2013), 其在本文模型中的生态承载力因受限于当前的生态条件而缺少大规模增殖的空间。这与法螺在商业渔业上的高价值特性有关, 其种群在过去遭受过度捕捞, 导致该物种存在灭绝的风险(Russo et al, 1990), 相关研究也显示法螺种群的衰退是推动珊瑚礁生境中海星不受控生长的主要原因(Brown, 1997; DeVantier et al,2007)。尽管由于底质环境差异, 珊瑚礁生境中双壳类动物的物种多样性相较于大陆沿海生境中要低(徐凤山 等, 2011), 但模型中双壳类功能组的当前生物量同最大生态承载力差距显著, 表明当前七连屿珊瑚礁生境中双壳类动物的增殖潜力巨大。砗磲作为在珊瑚礁中栖息的最著名的双壳类, 同时也是珊瑚礁生态系统中的重要组成部分, 其数量是评价珊瑚礁生态系统是否健康的重要指标之一(刘小霞,2017), 在一定区域范围内珊瑚礁生境的破坏会导致砗磲数量显著下降(Courtois de Vicose et al, 1999)。根据MTI 分析显示, 当前的捕捞活动对于法螺等具有高捕捞价值的高营养级动物具有显著的负面效应,由此直接或间接引发的棘冠海星暴发事件是造成七连屿珊瑚礁生态系统衰退的重要原因之一(李元超等, 2019), 同时也间接导致当前模型中双壳类功能组的生物量具有相对较高的增殖潜力。植食性鱼类为珊瑚礁生态系统提供重要的牧食功能, 以及提高珊瑚礁在气候变化相关干扰下的自我修复能力, 通常被认为是维持珊瑚礁生态系统健康的关键贡献者之一(Bellwood et al, 2004; Hughes et al, 2007; Bozec et al, 2016)。当前模型中植食性鱼类功能组具有较高的生物量和最大的生态承载力, 反映七连屿珊瑚礁生态系统可能出现由于大型肉食性鱼类遭受过度捕捞而导致其生态承载力区间收窄, 从而使植食性动物生物量的占比增加等现象(Arias-González et al,2004; Darling et al, 2017)。
依据前文有关七连屿珊瑚礁生态系统中珊瑚礁鱼类群落和珊瑚礁无脊椎动物群落增殖模式的模拟结果, 各模式下Ecopath 模型输出的总特征参数表明, 相较于当前七连屿珊瑚礁生态系统, 捕食者生物量一定程度的增加有助于维持生态系统的较高成熟度, 其中来自低营养级捕食者的作用更为明显。李元超等(2017)的研究显示, 当前模型中珊瑚礁鱼类受生境衰退和过度捕捞的影响而出现数量及密度下降, 结合本文中珊瑚礁无脊椎动物群落相较于珊瑚礁鱼类群落具有更高增殖潜力的模拟结果, 推测来自鱼类捕食所介导的下行控制对极端环境下底栖无脊椎动物的时空分布具有重要作用(Lin et al,2021)。此外, 不同于珊瑚礁鱼类群落, 珊瑚礁无脊椎动物群落由于受生境地形以及自身运动能力的限制, 食物来源相对单一, 但其构成的底栖食物网是碎屑进入系统能流的主要途径。群落增殖模拟显示,七连屿珊瑚礁生态系统作为典型的以上行控制为主导的珊瑚礁生态系统(Frank et al, 2007), 珊瑚礁无脊椎动物群落生物量的保障是珊瑚礁鱼类群落增殖的重要前提。因此, 在七连屿珊瑚礁渔业资源保护工作中, 除了要避免对珊瑚礁鱼类资源的过度捕捞, 更重要的是防止珊瑚礁栖息地被破坏, 以保护珊瑚礁无脊椎动物群落的规模, 维护珊瑚礁生态系统整体的稳定和平衡, 以及维持珊瑚礁渔业的可持续发展。
尽管本文在生态系统层次上对七连屿珊瑚礁的生态特征以及生态承载力进行了全面的评估和分析,但是仍然存在一些不足。Ecopath 模型作为一种静态模型, 并未考虑季节变化等时空因素所导致的环境因子变动而对模型产生的影响, 其预测结果存在可信度不高的风险, 因此模型预测的结果必须同实地调查结果相结合后谨慎使用。此外, 基于Ecopath所模拟的生态承载力以强调理论极限值为主, 缺乏关于各营养级生物生长发育对生态系统结构影响的考虑, 故难以科学地反映生态系统的真实状态。综合以上分析, 本文研究可为后续七连屿珊瑚礁生态系统的生态承载力动态变化研究提供背景基础, 未来还需开展长期的监测研究, 以便从生态系统水平为珊瑚礁渔业的开发和管理策略提供更为全面的科学理论基础。
猜你喜欢
中国交通信息化(2022年7期)2022-10-27
当代水产(2022年8期)2022-09-20
农业工程学报(2022年10期)2022-08-22
林业调查规划(2022年3期)2022-06-13
昆明医科大学学报(2022年2期)2022-03-29
家庭影院技术(2021年5期)2021-07-21
疯狂英语·新阅版(2020年3期)2020-09-22
百科探秘·海底世界(2020年8期)2020-07-29
安徽农学通报(2020年7期)2020-05-26
学校教育研究(2020年7期)2020-04-09
