
林木新种质创制研究进展
2022-02-16 06:29王军辉
中国农业科技导报 2022年12期
王军辉
(中国林业科学研究院林业研究所,林木遗传育种国家重点实验室,林木培育国家林业和草原局重点实验室,北京 100091)
林木是森林生态系统的主要组成,是全球碳循环的重要载体[1],既可利用叶片光合作用固定空气中的二氧化碳释放氧气,又能通过根系生长、细根周转将固定的碳以有机物形式释放到土壤中,在维持全球碳收支平衡和生态平衡中发挥重要作用[2-3]。我国木材需求量较大,是全球第一大木材进口国,而全球性天然林禁伐使得木材供用量锐减,木材供需矛盾不断加剧。林以种为本,必须自主培育产量、品质、抗性等目标性状具有重要突破的林木新品种,为国家生态和木材安全提供种质保障。
历经70多年的发展,我国在种质资源收集和精准评价、优良新品种选育、新种质创制核心关键技术及规模化繁育等方面取得了较大进展,由传统的“经验育种”向现代、定向、高效的“精准育种”转变,由宏观的群体、数量遗传研究深入到细胞、分子和基因水平[4-5]。目前,杨树(Populusspp.)、桉树(Eucalyptusspp.)、火炬松(Pinus taeda)等主要树种的基因组测序已经完成,全基因组选择、基因组编辑等精准育种技术取得重要突破,培育出一批抗旱、耐盐、抗虫的基因工程新品种[6-9]。在占全国乔木人工林面积和蓄积50%以上的杨树、杉木、落叶松、桉树、马尾松等主要造林树种中,建立了生态育种理论和高效育种技术体系[10]。这些为现代林木新种质创制和育种创新奠定了基础,使我国在同领域研究达到国际先进水平。但我国林木育种起步晚,加之林木树体高大、生长年限长等影响,仍然与林业发达的国家存在一定差距,需充分挖掘和利用林木种质资源,探究前沿育种技术及方法[11-12]。从林木新种质创制现状、核心关键技术、繁育和未来发展方向4个方面总结了我国林木新种质创制进展,并提出传统杂交育种与现代生物技术相结合,加快开发种质创制及繁育新技术的建议,以支撑优质种质创制和突破性新品种的培育。
1 林木新种质创制现状
1.1 林木种质资源收集和精准评价
植物种质是国家重要战略资源,种质资源数量、精准评价和开发利用能力已成为国家与科学实力的重要标志之一[13]。美国建成世界上第一座国家级种质库,保存59.3万份资源[14-15]。我国相继建立了各行业的种质资源保存库,其中国家作物种质资源库和种质圃收集保存种质资源52万份[14],西南野生生物种质资源库收集保存野生植物种质资源9万余份[15]。
我国林木种质资源收集保存和评价工作开始于20世纪七、八十年代,林木育种研究机构和林木种苗管理部门保存了一批主要造林树种的种源、家系、无性系、优良单株等种质资源与育种材料。国家林业和草原局(http://www.forestry.gov.cn/)共认定了294个国家林木良种基地、161个国家级林木种质资源库。国家林业和草原种质资源库包含各类保存库200余个,资源13.5万份,基本形成了覆盖全国的资源保存与共享服务体系。
种质资源鉴定评价逐步向精准化和规模化发展。各国主要在作物育种中建立了表型鉴定设施、高通量采集和分析数据,实现种质资源自动化、智能化鉴定。而林木种质资源精准评价工作相对落后,我国仅个别用材和经济树种开展了表型性状、品质性状以及分子性状的精准评价。随着测序技术发展,结合基因型数据,极大推动了林木功能基因组和重要性状精准评价研究[15]。研发林木表型精准评价技术,实现自动化、高通量、高精度的表型评价将为林木育种、生长监测、病虫害防治等提供基于大数据的决策支持。
1.2 林木优良新品种选育
林木优良新品种是林业生产和林业生态建设的重要基础。近10年来,我国相继完成了60余个树种的优良品种选育工作,国审林木良种289个,其中经济林172个,用材林75个,园林绿化用36个,珍贵树种和竹藤各3个(图1)。杨树良种数量最多,随后依次是枣、桃、杜仲、沙棘、苹果、核桃、葡萄和落叶松等(图2)。而热门树种(油茶)和生态观赏树种的良种数量和质量难以满足生产和生态需求,需进一步加强良种选育工作。
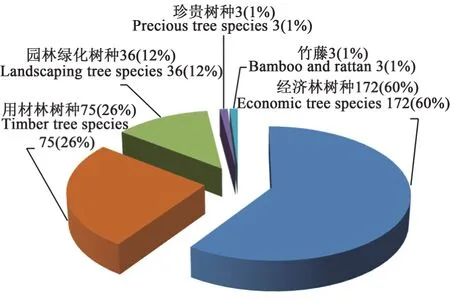
图1 2012—2021年各类树种国审良种审定数量及比例Fig. 1 Number and proportion of approved superior national varieties of various tree species from 2012 to 2021
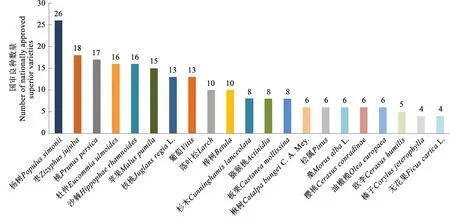
图2 2012—2021年主要树种国审良种数量统计Fig. 2 Statistics on the number of nationally approved superior varieties of major tree species from 2012 to 2021
1.2.1 速生用材树种良种选育 我国速生用材树种良种选育始于1946年,历经70余年,借鉴现代遗传学、生物技术、生物统计学成果,按照引种、选择、杂交、诱变、现代生物技术等育种理论与技术,系统开展了近60个主要造林树种的种源、家系和无性系等不同水平的遗传分析和综合选择,育成了一批速生性好、材质优良和抗逆性强的新品种[10]。
基于轮回选择不断推进以选择、交配、遗传测定为核心的育种循环,主要造林树种大多进入第一、二代遗传改良阶段,少数进入第三代甚至更高遗传改良阶段。一些阔叶树种在此基础上开展远缘杂交和多倍体育种,遗传增益提高30%以上;我国参与世界首例树木毛果杨的全基因组测序,率先完成鹅掌楸、白桦、银杏、杉木等树种的全基因组测序,绘制了杨树、杉木、马尾松、鹅掌楸等树种的高密度遗传图谱,定位了重要性状QTLs(quantitative trait loci),挖掘了一批重要功能基因[10,16]。建立了主要林木抗逆分子育种关键技术体系,创制出世界上首例转多基因抗逆林木新品种“多抗杨”。划分出杨树、杉木、落叶松、桉树、马尾松五大树种国家育种区,建立了现代育种技术体系,完善了林木生态育种理论,使我国林木育种理论和技术达到国际同类研究先进水平[10,17]。
1.2.2 珍贵用材树种良种选育 我国珍贵树种良种选育起步于1990年,逐渐形成了柚木、西南桦、降香黄檀、楸树、白桦、红松等南北方珍贵用材树种统筹兼顾,常规育种和分子育种相并行的格局。在种质资源保存与挖掘、新种质创制、优异性状机制解析、优良品系选育等方面积累了较为深厚的研究基础,育成了一大批良种,支撑了我国珍贵用材林的高质量建设与发展。
珍贵树种良种选育以常规育种为主。柚木中构建了生长与耐寒性多目标无性系选择技术体系,选育出国家级和省级良种6个。基于西南桦生长、形质和抗虫多个性状,选育国家级和省级良种8个,从表型和DNA水平上揭示了西南桦群体结构和遗传多样性,确定种质资源收集、保存和应用策略[18]。降香黄檀中,利用亚系育种策略建立育种群体,初选优良家系1个[19]。楸树中创立了四级优良无性系选育程序,缩短育种周期6~8年,为不同生态区选育出速生型、抗寒和抗旱型、耐水涝型国家级和省级良种29个[20]。根据白桦地理变异模式与规律,划分5个种源区,选育出国家级和省级良种12个[21]。选育光皮桦省级良种1个。开展白桦和光皮桦基因组测序,鉴定了与抗旱、耐盐、材性形成相关的转录因子。建立了白桦遗传转化系统、基因编辑技术等,获得了抗逆、花期调控、生长、材性等转基因株系156个。建立了红松单性状及多性状综合选择技术,选育国家级和省级良种 74个[22]。
1.2.3 经济林树种良种选育 我国是经济林大国,种植面积和产量均居世界首位。目前我国经济林种植面积约6.2亿亩(0.41亿hm2),林产品年产量逾2亿t,年产值超2.2万亿元,占林业第一产业产值的约60%[23-24]。
10年来,良种对产业的贡献率逐年提高,支撑了产业的高质量发展。我国六大木本粮油经济林树种包括油茶、核桃、仁用杏、板栗、枣和柿,先后选育出国审、省审新品种400余个,取得了显著的提质增效[25]。其中,核桃作为我国种植面积最大的经济林树种,选育出国审、省审新品种140余个,核桃干果单产由16.5 kg提高至20.5 kg;全国油茶种植面积约0.68亿亩(0.045亿hm2),选育出国审、省审和新品种120余个,年产茶油逾80万t[5],茶油单产由不足 10 kg提高至 12.2 kg[23,26];仁用杏全国种植面积约0.21亿亩(0.014亿hm2),选育出国审、省审新品种20余个,年产杏仁约25 万~30 万 t[27-28],杏仁单产由不足 9.2 kg提高至14.2 kg[23,27]。
我国已成为世界经济林种植大国,但不是产业强国,主要原因是科技支撑能力不足导致的产业低产、低质、低效的问题依然较为突出。迫切需要优质、高产、高抗、宜机化等聚合优良性状,适合不同主产区并能充分发挥潜力的“接穗+砧木+授粉”主栽品种配置组合的新一代良种,实现产业高质量发展。
1.2.4 竹藤良种选育 竹子隶属于禾本科竹亚科,全球有竹类植物80余属1 600多种;棕榈藤隶属于棕榈科省藤亚科省藤族,全世界有棕榈藤13属600多种[29-33]。竹藤植物良种选育的研究起步较晚,取得的成就集中于种质资源库建设和品种选育、杂交育种和生物技术育种等方面。
我国建有多处竹子资源库,如浙江安吉竹博园、安徽太平竹类种质资源库、福建华安竹种园、四川长宁世纪竹园、广东华南植物园竹类园、云南西双版纳植物园等。中国、印度尼西亚、马来西亚、菲律宾和印度等国建有棕榈藤资源库[34-36],这些资源库收集并保存了世界上绝大多数的竹藤资源。通过资源库建设选育适应性强、利用价值高的品种是竹藤良种选育的主要途径。杂交是实现竹藤种质创新和品种选育的重要途径,竹子杂交育种已有成功的范例,藤类则较为罕见。张光楚等[37]通过杂交选育出撑麻、麻版等杂交品种;宁材强等[38]通过杂交选育出撑绿系列品种袁金玲等[39]和Yuan等[40]选用耐寒亲本开展杂交,旨在选育抗逆性强的品种。竹藤植物的生物技术育种集中于基础研究领域,探明了部分竹藤品种的遗传多样性[41-42],建立了部分种类的快繁体系[43-44],破译了毛竹、麻竹、单叶省藤和黄藤等重要品种的全基因组序列[45-48],突破了麻竹、毛竹等竹种的遗传转化和基因编辑技术[49-51]。
1.2.5 木本花卉良种选育 从近10年数据看,木本观赏植物新品种授权逐渐增多,传统名花授权比率高达55%,前4甲分别是月季(691种)、牡丹(173种)、杜鹃花(163种)和山茶(115种),桂花和梅花分别列在新品种授权数量的第7和第14位,这些都属于我国的十大传统名花。月季主要采用人工杂交进行选育,少量通过芽变选育,极个别是自然杂交选育[52]。牡丹育种以常规杂交育种为主,主要通过自然杂交、突变选择等方法选育,但较少获取明显优于亲本的品种[53];与芍药以及野生滇牡丹进行远缘杂交,逐渐成为牡丹新品种选育的主要手段[54]。
传统名花在我国栽培历史悠久、群众基础好、市场广阔,为新品种的培育和推广奠定了良好的基础。授权的新品种以观花植物为主,占比达90%,观叶和观果植物仅占少数,如观叶植物槭树植物仅有53种,观叶又观果的卫矛属植物仅38种;以灌木为主,占65%,乔木为辅,占35%;绝大多数新品种的产权属于国内,占80%,仅少部分产权属于国外[55]。在授权的木本观赏植物中,一些传统不在观赏植物范畴的种类,由于出现新的变异(一般为叶色)成为新的观赏植物,如杨树、核桃、构树等。目前,木本观赏植物育种仍以传统人工杂交育种为主,少量通过芽变选育而来,极个别品种是自然杂交选育而来。
2 林木新种质创制的核心关键技术
2.1 种质资源评价与挖掘
林木种质资源是全球性重要的战略资源,是培育新品种的基本物质。种质资源的系统性、科学性评价和挖掘是培育适应社会需求和全球气候变化品种的核心基础,主要包括表型评价和遗传基础评价[56]。表型是林木在生长发育过程中与立地环境互作后而呈现出来的性状[57],主要包括树高、胸径、果实大小等形态性状,材积、重量等产量性状,木质素、纤维素、木材弹性、成分含量、果实颜色等品质性状及抗旱、抗寒、耐盐碱、抗病虫害等抗性性状[58-61]。
环境对林木种质资源各类性状的评价和挖掘具有重大影响,连年、多区域的持续评价可以准确获得相关性状的变异水平,为培育新品种提供准确信息[56-57]。林木种质资源性状评价主要采用田间鉴定、室内测量、仪器检测和数据分析等方式。近年来,随着设备的更新换代和多组学、智能与信息等技术的发展和交叉利用以及相关理论水平的提升,林木种质资源评价已从小样本的简单表型粗放型鉴定、分子标记等发展为大样本、大尺度、多区域、连续多年、多性状、高通量等方面[58-61],为林木种质资源精准、快速、高效评价和优异种质挖掘提供重要支撑。
2.2 育种群体构建
多群体育种策略是林木多世代改良、轮回选择的重要途径。新西兰辐射松、美国火炬松、加拿大云杉、欧洲落叶松等高世代遗传改良的成功,得益于大规模育种群体的建设、大批量控制授粉子代的创制、分组分层育种群体的构建和长期经营。加强育种群体组成、规模、结构与管理等设计和研究,能获得长期遗传增益,拓宽遗传基础,减少近交衰退。
通过对育种群体成员进行分组、亚系化及亚系内新种质的创制,为下一世代育种培育选择群体,具体环节为:① 对主群体作全同胞控制授粉,对精选群体则实行多系交配或自由授粉;② 主群体中各成员间可做较多的交配组合,并采用不同的遗传测验设计进行广泛测定,以实现亲本更精确的遗传评价和排序,使近交只能发生在主群体中;③ 通过无性繁殖无性系化核心组合的子代,改善家系内选择效果,快速实现世代转换,提高单位时间的增益[62]。
20世纪80年代开始,我国对40多个主要造林树种营建40多个种子园,基本完成了第一代改良。但第一代改良未能有计划、有目标地为下个世代改良选择创造足够大的选择群体;虽然建立了杉木、马尾松第二、三代种子园,仍未组建完善有效的育种群体,育种群体与生产群体混淆,子代测验群体当作育种群体。我国主要针叶树育种群体构建相比国际水平严重滞后、理论和技术薄弱,影响了遗传增益最大化、遗传多样性维持和良种生产的世代升级。目前,杉木营建第三代种子园[63];落叶松建立了核心育种区和高世代种子园,实现育种由一代向二代转型[64];系统收集和引入云杉种质资源3 869份,包含33个云杉种的270个种源;马尾松育种重点开展育种群体构建,针对材用和脂用性状构建了大规模的一代育种群体和较小规模的二代育种群体[65];建立了湿地松、火炬松种质资源库和生产性种子园,已建或正在建设第二代种子园,开展高世代杂交制种和测定工作[66]。
2.3 育种值估算方法
育种值是指有性繁殖的育种群体中,特定个体作为亲本在随机交配状况下传递给子代的性状加性遗传效应值,是影响该性状所有等位基因平均效应的总和,反映其育种贡献大小。数量遗传学中引入统计分析方法,通过构建表型模型,分析遗传效应和环境效应对表型变异的贡献,获得育种值估计值,个体的育种估计值排序可直接作为家系或单株选择依据。
育种值早期被视为固定效应,以普通最小二乘法(ordinary least squares,OLS)线性模型进行估算。由于交配设计及多世代改良累积、亲缘关系复杂性增加、区组和试验点数据不平衡等情况,混合线性模型分析法逐渐取代OLS,育种值被视为随机变量,产生了选择指数法、最佳线性预测法和 最 佳 线 性 无 偏 预 测 法(best linear unbiased prediction,BLUP)。BLUP法针对不平衡数据预测单性状育种值,采用包含固定效应和随机效应的混合线性模型,既能估算固定的环境效应和遗传效应,也能估测随机的遗传效应[67]。BLUP法估算和预测的各种效应遵循线性、无偏和最佳原则,公式如式(1)所示。BLUP法估算育种值的关键是确定线性模型结构,估计固定效应β值。在单株育种值预测时,可通过混合模型组计算大数据观测值的方差协方差矩阵的逆矩阵。相关遗传参数和矩阵计算可通过ASREML、DFREML、GAREML、TREEPLAN、SAS和 Matlab等 软 件完成[68]。

式中,y为观测值向量,β为固定效应,u为随机遗传效应,e为随机误差效应,X和Z分别为β和u的关联矩阵。
当前林木改良的实际应用中,BLUP法及其衍生的标记辅助/基因组BLUP法、BLUP-GGE(BLUP-genotype main effects and genotype ×environment interaction)联合分析等方法在火炬松、日本落叶松、辐射松、杉木、湿地松、桉树等世界主要人工造林树种的单性状育种值预测中广泛应用[69-71]。多性状综合育种值预测时,对每个性状单独BLUP分析后可将所有性状的数据综合起来产生选择指数,在亲缘关系及具体育种目标约束下,按指数排序进行选择[72]。
2.4 基因组选择育种
常规育种难以高效、快速培育林木良种,分子标记辅助选择(marker-assisted selection,MAS)方法无法捕获微效位点、遗传作图分辨率低,不能有效推动林木遗传改良进程。鉴于性状表型选择和MAS的不足,Meuwissen等[73]提出基因组选择(genomic selection,GS)。GS是利用覆盖全基因组的高密度遗传标记信息辅助选择的早期选择技术,具体程序为:① 建立训练群体,测定所有个体的表型和基因型,利用统计模型估计每个SNP(single nucleotide polymorphism)的效应值,建立表型和基因型间的GS模型;② 对候选群体的个体进行基因分型,利用GS模型计算候选群体个体基因 组 育 种 值(genomic estimated breeding value,GEBV);③ 根据GEBV排名在种苗期对林木个体进行选留,选留个体完成表型测定后再放入训练群体,更新和优化GS预测模型。林木GS真正实现利用基因组信息指导育种实践,有效加速遗传改良进程,为林木智能设计育种提供核心技术。
GS是目前动物和作物遗传育种的关键技术,也成为林木育种技术创新的热点,主要由美国、巴西、瑞典等针对重要树种研发GS育种技术,但仍处在理论研究阶段。2008年至今,林木GS领域有约80项成果发表,集中于桉树、云杉、火炬松等用材树种,针对生长、材性和抗病虫性状,探究影响预测准确性的因素,包括训练群体规模与群体结构、性状遗传力、标记数量、模型算法对比和优化等[74-77],但GS实践和跨越多世代的GS研究鲜有报道。
为促进林木GS育种发展,应建立基因型和表型获取平台,高通量、低成本、标准化获取基因型和表型数据。根据林木特点优化和开发林木GS模型算法,结合多组学、多环境和多年份数据,开发基因型×环境/年龄互作模型,提高GS模型的普适性和长期功效性。整合分析多群体大样本数据,获得稳定可靠分子标记,开发特定性状SNP芯片产品;促进数据收集和共享,注重研究机构和育种公司合作,保证长期稳定投入和研究[78-80];促进林木GS与早花、体细胞胚发生、种子基因型鉴定、基因编辑等技术耦合,进一步缩短育种周期[81]。
2.5 基因编辑等分子改良
基因编辑是基于序列特异核酸酶在基因组水平上对靶标基因进行定向、准确修饰的基因工程方法,包括锌指核酸酶、类转录激活因子效应物核酸酶、巨核酶和CRISPR/Cas9系统等。CRISPR/Cas9是基于细菌自身适应免疫防御系统改造的方法,操作更加简便、敲除效率更高、编辑更精准,具体环节为:① 经人工设计的单一指导的核糖核酸sgRNA包含能被Cas9蛋白特异性识别的发卡结构,二者结合形成切割复合体;② sgRNA上的序列通过碱基互补配对,特异性识别外源基因组原型间隔序列毗邻基序PAM NGG位点;③sgRNA引导Cas9蛋白对PAM位点附近的DNA双链进行切割,造成断裂;④ 断链的DNA通过非同源末端连接修复,形成碱基突变和插入缺失,造成基因敲除或敲入,完成对基因组DNA的修饰。
自2013年来,CRISPR/Cas9系统已应用于24个属45个科植物的基因功能研究和重要性状遗传改良中[82]。2014年,CRISPR/Cas9系统首次应用于林木树种甜橙,虽未获得完整的基因编辑植株,但证实了其可行性[83]。接着在模式树种杨树和经济林木橙子、苹果、梨、咖啡等多个树种,逐步建立了CRISPR/Cas9技术体系,实现抗旱、抗病等林木新品种的培育,在调控木质素合成、缩短林木育种周期等方面也发挥重要作用[84]。杨树发展最为迅速,如成功敲除八氢番茄红素脱氢酶基因导致杨树白化表型[85]。Estefania等[86]成功编辑4-香豆酸辅酶A连接酶基因,获得茎秆呈红褐色的银灰杨突变体材料。Bruegmann等[87]利用SOC1等9个杨树内源基因,分析比较了不同sgRNA产生基因编辑的效率。
多数树种尚处于CRISPR/Cas9体系建立的初步阶段,为促进发展需完善和加强靶位点设计分析平台基因组数据的收录,建立更多树种的sgRNA设计和脱靶效应检测公共网络平台[88];研发更多树种的原生质体制备体系、组培再生体系和稳定的遗传转化体系[84],提高单细胞转化效率,直接向细胞导入Cas9/sgRNA复合物解决外源DNA污染问题。
3 新种质规模化繁育
3.1 无性快繁
无性快繁是林木繁育技术的重要组成部分。针对选育出的优良单株,通过扦插、嫁接、组织培养等技术,可在短时间内培育大量苗木,既推广了良种,又提高了遗传增益。扦插是杨树良种利用和推广的主要方式,扦插及其配套的采穗园幼化技术在多种松科植物育种实践中也应用广泛[16,89]。嫁接可繁育优质苗木、缩短结实时间,在红松的生产实践中使用较多[90],桉树中应用组织培养技术相对较多[91]。
再生是无性快繁技术中的重要生物学过程,影响再生的因素也限制着快繁技术的应用,比如母株的基因型和年龄、繁殖材料在母株上的位置、环境因素和外源激素[16,92-95]。揭示再生过程潜在的调控机理有助于更高效、稳定地开发和利用无性快繁技术。此外,完善的组培再生体系能为基于基因编辑的林木遗传改良提供关键技术保障。因此,深入挖掘林木无性快繁技术及其调控机制,既是林学的基础理论研究热点,又对指导林业生产有重要意义。
3.2 种子园矮化结实
种子园在林木种子生产、良种培育过程中发挥着不可替代的作用,大多数树种造林仍依靠种子来源的苗木。然而,随着树龄增加和树体增大,结实部位逐渐上移,给人工授粉、采种、采条等操作带来不便,增加了种子的生产成本,是种子园生产经营过程中面临的难题。因此,种子园“矮化”是良种基地的经营方向和迫切需要解决的问题[96]。
重新建立矮化种子园或对现有种子园母树进行矮化处理,可实现种子园“矮化”。重新建立矮化种子园所用苗木是在品种培育过程中选择出来的,或常规苗木经过嫁接、修剪和整型培育出来的,未来不会生长太大。现有种子园中的高大母树经过截干去顶和疏枝后也可以得到矮化、果实增产的目的,这也是国内目前比较成熟的做法[97-99]。由于树种本身的遗传和生长特性,同样的矮化处理措施在不同树种中取得的效果不同。树木生长发育机制的研究有利于有效地管理树体,科学地建立矮化种子园。培育矮化新品种,从遗传上改变树木生长特性,创制矮化新种质是未来重新建立矮化种子园的有效途径。
3.3 体细胞胚胎发生
体细胞胚胎发生技术是单个体细胞或组织在离体条件下经过脱分化、再分化形成胚胎的技术。林木体细胞胚胎发生研究始于20世纪70年代,该技术是推动林业无性系化的关键技术,由于繁殖效率高,被认为是针叶树最重要的无性繁殖技术[100-102],有助于实现针叶树优良新品种的快速繁育。目前,已有冷杉属、落叶松属、云杉属、松属、黄杉属和北美红杉属的50多个针叶树树种开发了体胚技术[102-107]。欧洲和北美等林业发达国家体胚技术较为成熟,已应用于苗木生产,年产量可达千万株[101],产生了巨大的经济和生态效益。
成本控制是大规模推广体胚技术必需面临的挑战。随着生产规模的扩大,实验室规模下微乎其微的原料和人工成本都会急剧增加,如何在不影响体胚质量的前提下降低成本是现在很多公司和机构十分关注的问题。推广体胚繁育技术需要进一步提高规模化和自动化程度,建立监测冻存和生产材料状态的数据库、研究光照对体胚苗发育的影响,以达到高效、高质、经济地生产数千万体胚苗的培育目标。
4 林木育种未来发展方向
4.1 建立林木高通量精准评价技术体系,充分挖掘利用种质资源
林木高通量精准评价技术体系的建立有助于开发利用种质资源,为林木育种、生长监测等提供基于大数据的决策支持。生物技术的发展使得基因组、转录组、蛋白组、代谢组、表型组等多组学方法逐步成为林木种质资源评价的重要工具。通过无人机高通量表型观测,研发适合林木特点的可移动、折叠式的低成本表型工厂,建立林木高通量精准评价技术体系,对收集保存种质资源进行精准评价分析,筛选关键性状特异种质;联合全基因组功能性标记和基因型精准鉴定,充分挖掘种质资源特殊价值,创制具有突出性状的育种新材料,将是林木种质创新的重要方向[108-109]。
4.2 建立林木全基因组选择育种技术,高效设计林木新种质
全基因组选择是结合功能基因组研究成果和SNP芯片技术的早期选择育种技术[73,110],具有巨大应用潜力。目前GS分析模型多基于家畜和农作物,无法满足林木目标性状纵向分析需求。需基于大量模拟和真实数据评估和验证,建立符合林木特性的GS育种模型。未来20~30年内,应率先在常规育种程序完整的树种中建立GS育种平台,逐步构建以林木“基因型-GS模型-表型”为核心的良种选育智能决策系统,实现定向、高效、早期选育符合市场和生态需求的林木新种质,将是林木遗传育种的发展方向。
4.3 建立符合林木特点的基因组操作技术,精准创制突破性新种质
林木体积大、生长年限长、育种效率不高,通过基因组编辑技术可实现林木多基因控制性状的直接遗传改良,提高育种效率。目前,毛果杨、挪威云杉、巨桉、鹅掌楸等主要树种的基因组测序已经完成[7,85-86,111-112],将基因组、转录组、代谢组、蛋白质组等相结合,定位克隆调控目标性状的关键基因和生物元件,建立符合林木特点的基因组操作技术,对目标性状进行精准改良,可实现林木多性状整合,定向创制突破性林木新种质,对于加快林木育种进程和培育高产、优质、抗性强的战略新品种具有重要意义。
4.4 利用合成生物学等手段创制林木新种质
自然环境下林木变异频率低、幅度小,且有益变异难,难以满足实际需求。合成生物学、航天诱变等手段可以在较短时间创造优良新种质,加快林木新品种选育[113-114]。林木合成生物学研究具有独特的优势,如合成生物过程需要消耗的大量糖类可通过光合作用固定大气中CO2提供[115]。航天诱变对植物的生长、形态特征、物候期等产生积极影响,为林木种质创新提供新途径[116-117]。随着全球气候变化,极端天气频繁出现,合成生物学、航天诱变等技术将成为创制新种质,进而提高林木产量、质量及适应性的重要手段。
4.5 建立细胞工厂,智能化工程化繁育种苗
细胞工厂是将细胞设计创建成为一个以可再生细胞为原料的生产工厂,具有绿色、清洁、可再生的特征[118]。细胞工厂在林木领域的主要应用方向是苗木育种与生产,利用细胞全能性通过组织培养和体胚繁育技术大量繁殖具有相同基因型的良种苗木。相比天然种子,通过细胞工厂生产的人工种子既能保留杂种优势,也能通过包裹材料添加不同物质影响植物的生长,对于种苗培育具有重要意义[119]。细胞工厂需要多学科的汇聚和技术整合,是经济社会可持续发展的重大方向,在未来林业发展中将掀起新一轮的革命。
5 结语
我国林木资源丰富、生态建设和产业发展需求多样,未来在进一步加强种质资源收集、保存基础上,需结合传统杂交育种与现代生物技术,加快开发林木高通量精准评价、全基因组选择、基因组操作、合成生物学、细胞工厂等育种新技术,创新林木育种理论,突破性新种质精准创制和规模化繁育技术,提升林木种业科技自主创新能力,实现林木高效育种,服务国家行业发展需求。
猜你喜欢
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
林业科技(2020年3期)2021-01-21
意林·少年版(2020年17期)2020-10-12
发明与创新·中学生(2020年4期)2020-04-17
四川蚕业(2020年4期)2020-02-10
农民致富之友(2018年12期)2018-06-29
中国麻业科学(2018年6期)2018-04-09
现代园艺(2018年2期)2018-03-15
现代园艺(2017年23期)2018-01-18
