
9 个抗稻瘟病基因在291 份水稻种质资源中的分布及组合特征
2022-02-13 09:11桑世飞王亚男王君怡曹梦雨孙晓涵姬生栋
河南农业科学 2022年12期
桑世飞,王亚男,王君怡,曹梦雨,孙晓涵,姬生栋
(1. 河南师范大学 生命科学学院,河南 新乡 453007;2. 河南省作物遗传改良与种质创新工程研究中心,河南 新乡 453007)
我国是世界上最大的水稻生产与消费国,平均每年水稻生产与消费量超过全世界总量的30%[1-2]。因此,确保水稻高产稳产与国家粮食安全紧密相关。水稻稻瘟病是水稻生产中一种重要的病害,严重影响水稻产量。在稻瘟病较流行的年份,其可致水稻减产10%~40%;在稻瘟病暴发的年份,其可致水稻减产50%~60%[3-4]。而生产上应用的水稻品种由于稻瘟病菌生理小种的快速变异、栽培环境和生态环境的改变致使生态遗传抗性减弱,品种的抗病性下降,极大地制约了水稻的农业生产[5]。因此,为降低因稻瘟病暴发带来的产量及经济损失,选育抗稻瘟病水稻品种显得尤为重要。
近年来,关于广谱稻瘟病抗性基因的克隆、鉴定与分析的研究已取得一系列突破,为水稻稻瘟病抗性改良奠定了基础。目前,已鉴定出100 多个抗稻瘟病主效基因与500个稻瘟病抗性数量性状位点(QTL)[6-7],其中有37 个基因被成功克隆和鉴定,NBS-LRR(Nucleotide binding site-leucine-rich repeats)类基因是水稻中应用较广泛的稻瘟病抗性基因,在水稻抗稻瘟病方面具有重要作用。例如,已经克隆的广谱稻瘟病抗性基因有Pi2、Pi9、Pi-kh、Pi50、Pi54、Pi54rh、Pi56(T)、Pi64、Pigm、Pizh和Pita-2等[8-11]。其中,位于6号染色体上的Pi50、Pigm、Pizh、Pi2、Pi9同为Piz 位点上的复等位基因[12]。一部分NBS-LRR 类基因已被应用于水稻抗稻瘟病品种的选育,例如,吴婷婷等[13]应用Pi9基因改良了水稻不育系丰源A的稻瘟病抗性。
随着水稻功能基因组与分子标记技术的快速发展,分子标记辅助选择与传统育种方法相结合成为现代育种的趋势,其克服了传统育种效率低、周期长等问题。目前,已经利用分子标记辅助选择选育出一些稻瘟病抗性种质。例如:何弯弯等[14]将含有Pib抗稻瘟病基因的品种津稻179 与徐稻3 号进行杂交,通过水稻系谱法选育及田间接种和自然诱发鉴定,对受体的抗病性进行改良,最终获得具有优良农艺性状且水稻稻瘟病抗性增强的X13、X40、X41 和X42 四个品系;吕军等[15]利用分子标记辅助选择聚合水稻抗稻瘟病基因Pita和香味基因fgr,最终筛选出含有抗稻瘟病基因Pita和香味基因fgr的4 个辽粳香系列品系;何光明等[16]通过分子标记辅助选择结合回交转育,首次成功地将抗衰老基因IPT(Isopentenyl transferase)、抗白叶枯病基因Xa23和抗稻瘟病基因Pi26聚合到一起;宋泽等[17]以携带Pi1和Pi9抗病基因的BC4F2群体为基础材料,利用分子标记辅助选择完成BC5F5株系的构建,改良了水稻特异种质大粒溪香的稻瘟病抗性;曾晓强等[18]通过连续回交结合分子标记辅助选择,将抗稻瘟病基因Pi2导入到高产优质晚粳品种长农粳1 号中,改良了长农粳1 号的稻瘟病抗性;姚姝等[19]利用分子标记辅助选择选育出产量高、抗病性好的品系南粳0051;潘存红等[20]用携带Pizt和Pib基因的粳稻品种武运粳21 号与优质、高产、感稻瘟病的中间材料L0 杂交,通过分子标记辅助选择获得6 个不同类型的稳定株系,显著提高了L0的稻瘟病抗性。然而对于品种或品系内的稻瘟病抗性基因组合特征缺乏系统性研究。为进一步深入研究当前水稻品种(品系)中稻瘟病抗性基因的组合特征,对收集的291份水稻种质资源进行抗稻瘟病基因组合分析,系统分析当前生产上水稻品种(品系)中的稻瘟病抗性基因组合特征,为今后抗稻瘟病分子标记辅助选择育种提供理论依据。
1 材料和方法
1.1 试验材料
供试材料为来自河南(177份)、江苏(39份)、天津(23 份)、浙江(9 份)、国际水稻所(8 份)、宁夏(4份)、山东(5 份)、安徽(5 份)、湖南(3 份)、河北(2份)、福建(2份)、上海(1份)及东北三省(黑龙江、吉林、辽宁)(13 份)等省(市、区)的共计291 份水稻种质资源,以黄淮海区域种质为主,主要由新乡市农业科学院、河南省农业科学院、盐城市农业科学院、浙江省农业科学院、河南师范大学水稻新种质研究所等单位提供。
1.2 试验方法
1.2.1 抗稻瘟病基因分子标记收集及Pib基因分子标记开发 通过查阅文献收集相关稻瘟病抗性基因的分子标记(表1)。其中,有5 个共显性标记,分别为Pi9、Pi5、Pi2、Pita、Pi-kh;3个显性分子标记,分别为Pikm、Pit、Pb1。所有分子标记的引物均由生工生物工程(上海)股份有限公司合成。
从 GenBank(https://www. ncbi. nlm. nih. gov/genbank/)数据库检索下载Pib抗病基因型序列(登录号:AB013448.1)以及感病种质资源9311的Pib等位基因序列,设计引物(F:5'-TCCTGTCCCAGAAGACATAT-3';R:5'-GTGACGGCGGATGCCTGACG-3')扩增收集的种质资源津稻10 号(Jindao 10)、新科稻29(Xinkedao 29,简 称XKD 29)、新 科 稻89(Xinkedao 89,简称XKD 89)的基因组序列,扩增产物送生工生物工程(上海)股份有限公司进行Sanger测序,采用MEGA 5.1 进行序列比对分析,以Pib感病、抗病基因型差异位点开发特异引物。
1.2.2 PCR 扩增及电泳检测 采用CTAB 法[25]提取水稻幼嫩叶片基因组DNA,以诺唯赞GreenTaqMix推荐浓度,设置10 μL PCR体系,根据各分子标记引物的Tm 值设定退火温度进行PCR 扩增。150 V 电压、1%琼脂糖凝胶电泳15 min 检测扩增的目的条带。
2 结果与分析
2.1 Pib分子标记开发
将Pib基因抗病基因型序列与水稻9311、津稻10 号、新科稻29、新科稻89 中的Pib等位基因序列进行比对(图1),发现序列比对区间存在多处变异,基于变异序列设计2 个分别针对性扩增Pib感病与抗病基因型的分子标记,且互相不干扰,能够混合在一起实现共显性扩增。在Pib抗病基因型特异区域 开 发 出1 个 显 性 抗 病 分 子 标 记(PibFk:5'-AGCGAGGAAGCGGAGGGTGG-3',PibR:5'-CAATGCATTTAA(G/A)CGTGCTCAC-3');同时在Pib感病基因型特异区域设计1 个显性感病分子标记(PibF:5'-CTGCTCGTTTGCGACCAATG-3',PibRg:5'-GTTGAAGCCCTAGGATCCGCT-3')。理论上抗病材料可扩增出500 bp 的目的片段,感病材料可扩增出350 bp的目的片段。
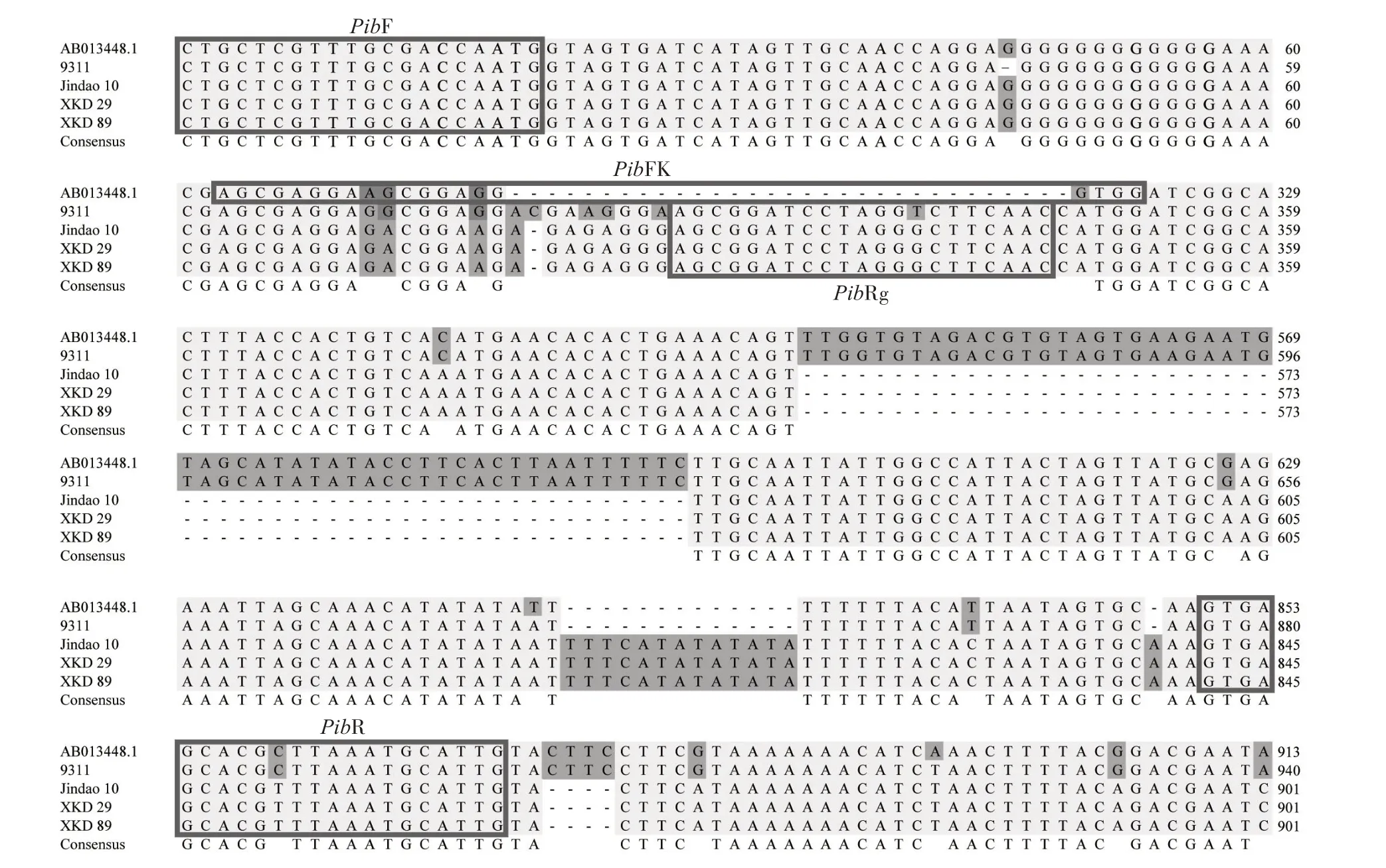
图1 不同水稻种质资源Pib基因序列比对分析Fig.1 Sequence alignment analysis of Pib gene in different rice germplasm resources
将上述2 对引物混合在一起,选取12 份种质资源进行Pib基因型检测,结果筛选出6 份含有Pib抗病基因型的水稻种质(玉粳3、联丰88、华粳5 号、新稻59、新粮310、武运粳21),扩增出500 bp 的目的条带,6 份含有Pib感病基因型的水稻种质(圣稻806、镇稻99、新稻89、菡香1 号、新丰5 号、华粳5号),扩增出350 bp 的目的条带(图2),表明Pib分子标记可应用于水稻抗稻瘟病基因型的检测。
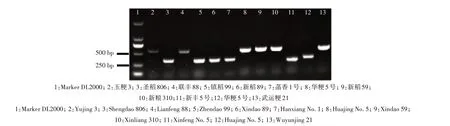
图2 基于Pib基因分子标记的抗(感)稻瘟病水稻种质资源中Pib基因型鉴定Fig.2 Identification of Pib genotype in rice germplasm resources with blast resistance(susceptibility)based on molecular markers of Pib gene
2.2 9个抗稻瘟病基因在水稻种质资源中的分布
本研究利用前人报道以及本研究开发的Pib基因分子标记(2 对引物混合)共计9 个抗稻瘟病基因分子标记对291份水稻种质资源中的抗稻瘟病基因进行检测(部分结果见图3),发现Pi9、Pi2、Pib、Pikh、Pita、Pi5这6 个基因的分子标记具有共显性标记特征。
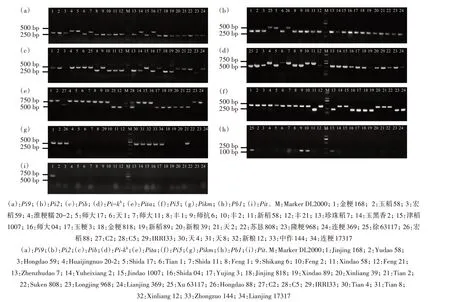
图3 部分水稻种质资源中9个抗稻瘟病基因的分子检测Fig.3 Molecular detection of nine blast resistance genes in some rice germplasm resources
对水稻种质资源携带的抗稻瘟病基因数目进行分析(图4),发现单份水稻种质资源携带的抗稻瘟病基因数目为0~6 个,其中携带6 个抗稻瘟病基因的种质资源仅有1 份,即垦香48,占比最低,为0.4%;携带5 个抗稻瘟病基因的种质资源有26 份,占比8.9%,包括武运粳80、玉粳3、新稻59、艺稻628、新稻58 等;携带4 个抗稻瘟病基因的种质资源有56份,占比19.2%;携带3个抗稻瘟病基因的种质资源有119份,占比最高,为40.9%;携带2个抗稻瘟病基因的种质资源有59份,占比20.3%;携带1个和0 个抗稻瘟病基因的种质资源分别有24 份和6 份,分别占比8.2%和2.1%。
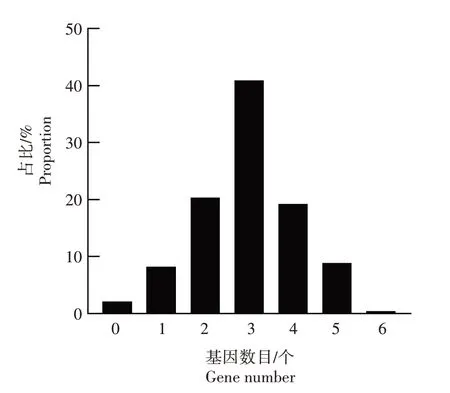
图4 水稻种质资源携带抗稻瘟病基因数目分析Fig.4 Analysis on the number of rice blast resistance genes carried by rice germplasm resources
对291份水稻种质资源中各个抗稻瘟病基因的出现频率进行分析(图5),发现9 个抗稻瘟病基因在水稻种质资源中出现的频率存在较大差异,介于7.2%~59.1%。其中,Pi9基因出现频率最高,为59.1%;Pi-kh次之,为50.8%;Pib基因出现频率为41.2%;Pi5基因出现频率为36.4%;Pita基因出现频率为29.2%;Pb1基因出现频率为28.8%;Pi2基因出现频率为19.5%;Pikm基因出现频率为17.1%;Pit基因出现频率最低,为7.2%。
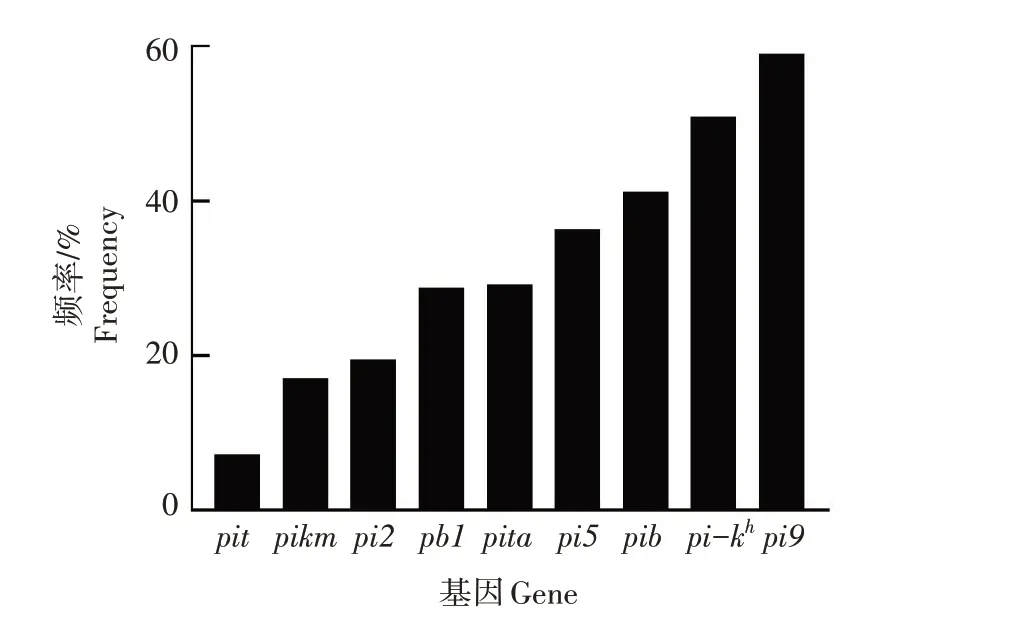
图5 9个抗稻瘟病基因在水稻种质资源中的出现频率Fig.5 Occurrence frequency of nine blast resistance genes in rice germplasm resources
2.3 291份水稻种质资源中9个抗稻瘟病基因的组合类型
对291份水稻种质资源的抗稻瘟病基因组合进行分析(表2),发现291 份水稻种质资源中共有抗稻瘟病基因组合93 个。其中,Pi9+Pi-kh+Pita组合水稻种质资源最多,有18 份;其次为Pi9+Pi-kh+Pib组合,有17 份;组合为Pi9+Pb1的水稻种质资源有11份;组合为Pi9+Pi5+Pb1、Pi9+Pi-kh和Pi-kh+Pita的种质资源各有9 份;含有Pi9+Pi5+Pi-kh+Pib+Pita、Pi9+Pi5+Pi-kh和Pi-kh组合的种质资源各有8份;含有Pi9+Pi5+Pib和Pi9组合的种质资源各有7份;含有Pi2+Pib+Pit、Pi2+Pi5+Pi-kh和Pi5+Pi-kh组合的种质资源各有6份。而其余组合数目则相对较少,如含有Pi9+Pi-kh+Pib+Pb1和Pi5+Pi-kh+Pib组合的种质资源各有5份;含有Pi9+Pib+Pita+Pikm+Pb1、Pi9+Pi-kh+Pib+Pita、Pi9+Pi5+Pi-kh+Pib、Pi2+Pi-kh+Pib+Pb1、Pi2+Pib+Pikm+Pb1、Pi9+Pi5+Pikm、Pi2+Pita+Pikm、Pi5+Pb1、Pi9+Pi5和Pi5组合的种质资源各有4份;含有Pi9+Pi5+Pib+Pita+Pikm、Pi2+Pib+Pita+Pikm+Pit、Pi9+Pi5+Pi-kh+Pb1、Pi9+Pi5+Pikm+Pb1、Pi9+Pi-kh+Pita+Pb1、Pi2+Pi5+Pi-kh+Pib、Pi9+Pib+Pb1、Pi9+Pita+Pb1、Pi9+Pi-kh+Pb1、Pi2+Pib+Pikm、Pi2+Pi-kh+Pib、Pi5+Pib和Pi9+Pib组合的种质资源各有3 份;含有Pi9+Pi5+Pi-kh+Pib+Pb1、Pi9+Pi5+Pita+Pikm+Pb1、Pi9+Pi5+Pi-kh+Pita、Pi9+Pib+Pikm+Pit、Pi9+Pi5+Pib+Pb1、Pi2+Pib+Pikm+Pit、Pi2+Pi-kh+Pib+Pit、Pi9+Pib+Pita、Pi2+Pi5+Pib、Pi2+Pi-kh、Pib+Pita、Pb1和Pita组合的种质资源各有2份;其余均仅有1份,如表2所示。
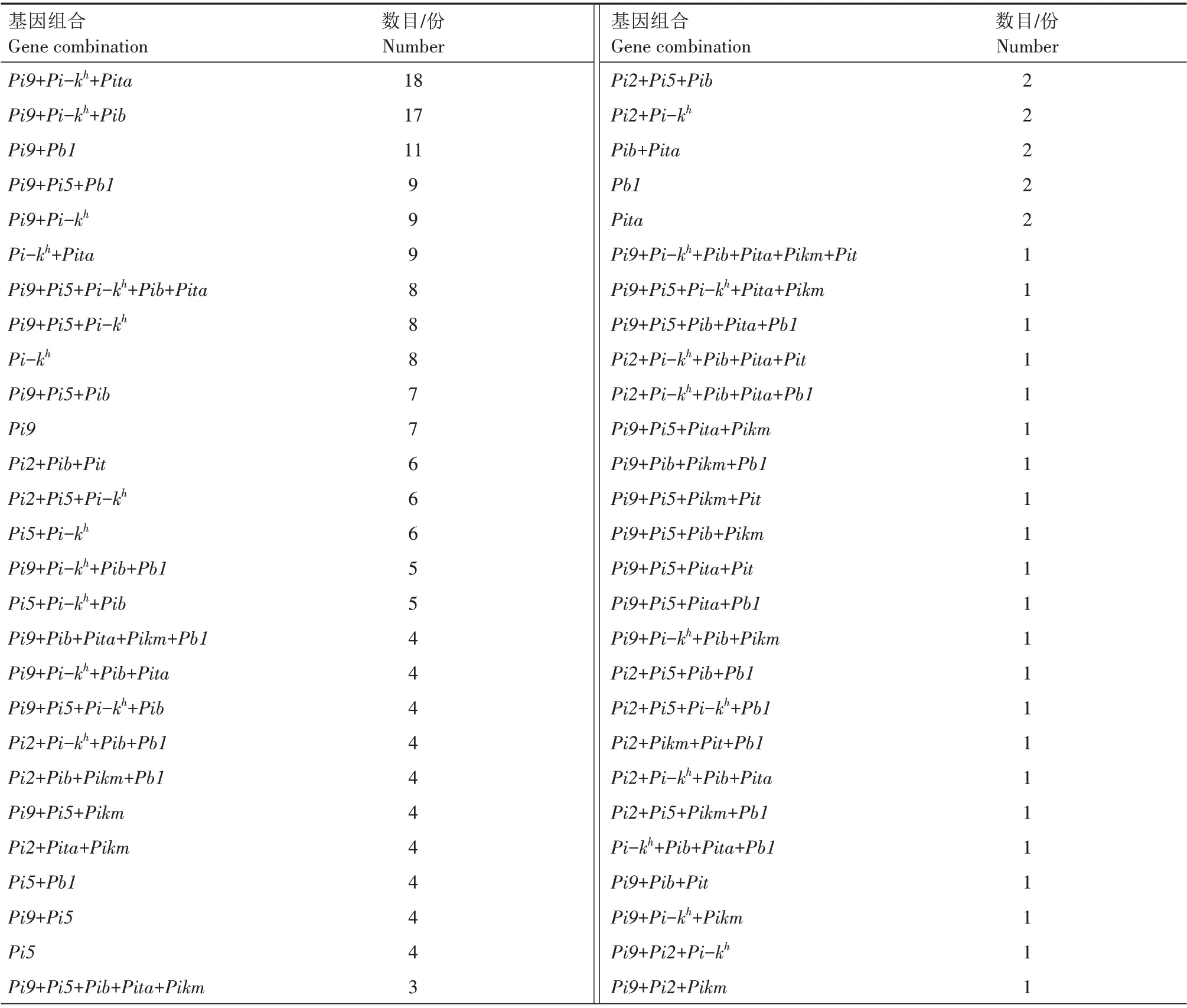
表2 291份水稻种质资源9个抗稻瘟病基因组合分布Tab.2 Distribution of nine blast resistant gene combinations in 291 rice germplasm resources
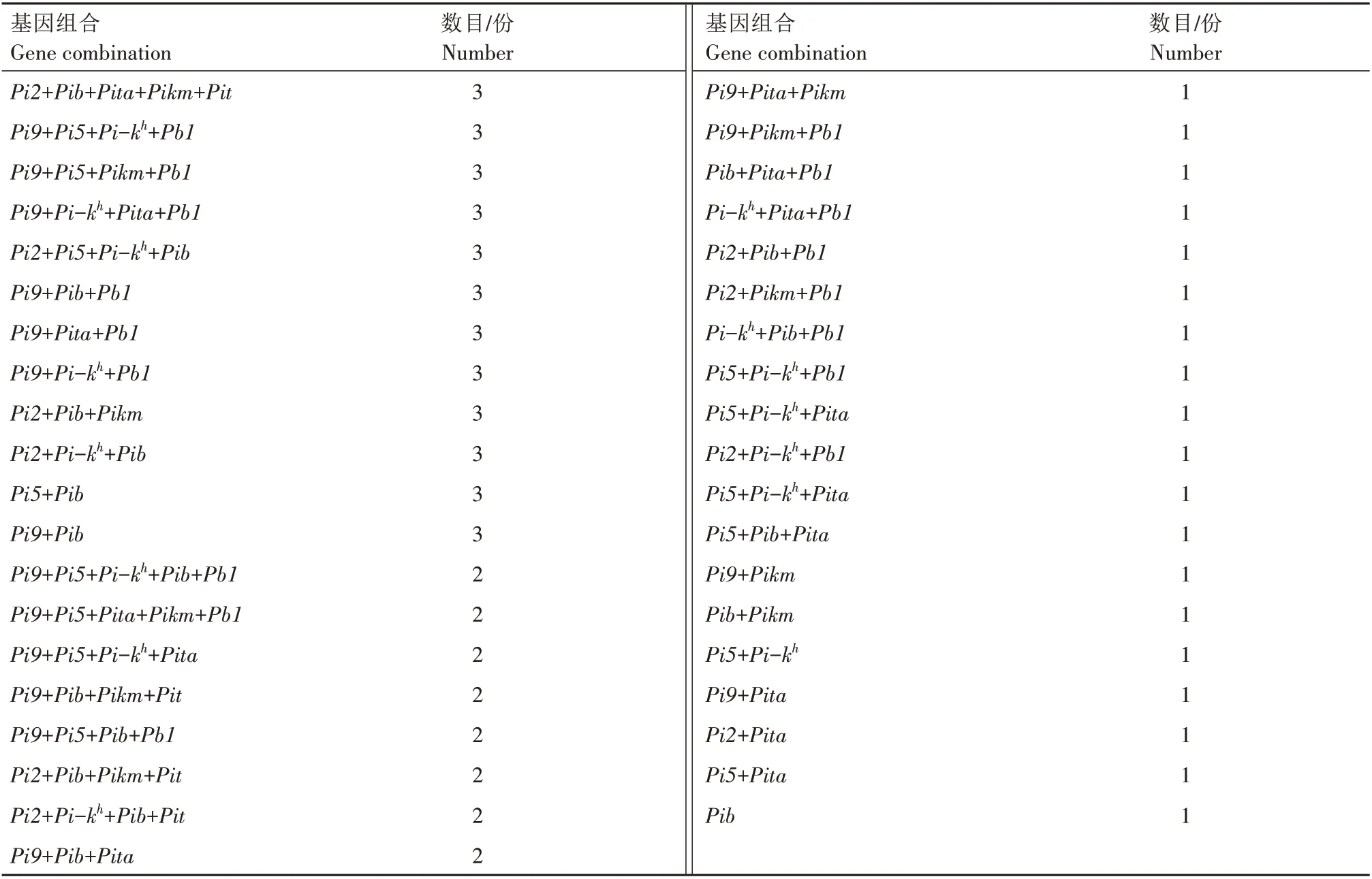
续表2 291份水稻种质资源9个抗稻瘟病基因组合分布Tab.2(Continued) Distribution of nine blast resistant gene combinations in 291 rice germplasm resources
3 结论与讨论
稻瘟病是当前水稻生产中威胁最大的一个真菌性病害,且稻瘟病菌变异快,育成品种稻瘟病抗性容易丧失[11]。因此,解析水稻稻瘟病抗性、选育抗稻瘟病水稻新品种已经成为当前解决稻瘟病的一个重要途径。
利用广谱抗性基因分子标记检测水稻品种中含有的抗稻瘟病基因类型、组合,分析各地区抗稻瘟病基因的分布比例,对特定地区种质资源的稻瘟病抗性改良具有重要的指导意义。研究发现,在不同地区抗稻瘟病基因的利用频率存在一定的差异。王亚等[22]对河南省139 份水稻种质资源中的Pi-kh和Pi5基因进行检测,发现Pi-kh基因出现频率为48.4%,而浙江省水稻种质资源中Pi-kh基因出现频率则为41.3%[24]。说明不同地区可能由于稻瘟病生理小种的不同选育出来的水稻品种所携带的抗稻瘟病基因存在一定的差异。本研究发现,Pi9和Pi-kh基因在291 份水稻种质资源中的出现频率均高于50%,Pib基因出现频率超过40%,表明这些基因在收集的种质资源中的应用频率较高,另外这些种质分布较广,可选择部分携带这类抗稻瘟病基因的种质作为以后育种的主要抗性材料。本研究还发现,Pi5、Pit、Pi2、Pb1、Pita、Pikm基因在291 份水稻种质资源中的出现频率相对较低,在未来抗稻瘟病基因改良中有必要加大对Pi5、Pit、Pi2、Pb1、Pita、Pikm基因的利用率,以便提高稻瘟病抗性及持久性。
在育种实践中,聚合抗稻瘟病基因可以提高水稻种质资源稻瘟病抗性。例如:陈红旗等[26]利用分子标记技术聚合Pi-1、Pi-2和Pi-33三个抗稻瘟病基因,提高了金23B 的稻瘟病抗性。然而水稻种质资源中聚合过多的抗稻瘟病基因可能会出现连锁负效应[27];聚合数目过少,则难以维持水稻种质资源的稻瘟病长期抗性。本研究发现,含有3~4 个抗稻瘟病基因的水稻种质资源共占60.1%,意味着在水稻抗稻瘟病基因聚合育种时,聚合3~4 个稻瘟病抗性主效基因基本可以满足生产或品种审定的需求,然而聚合稻瘟病抗性基因数目大于4的种质资源依旧相对稀少,若要继续维持老旧品种的生命力,则有必要对当前推广的品种做进一步的抗稻瘟病基因聚合,以提升对多种稻瘟病生理小种的抗性,延长水稻品种的生命周期。吴云雨等[28]研究发现,不同生态区选择合适的抗性基因组合,才能够保证水稻稻瘟病的持久抗性。本研究发现,携带Pi9+Pi-kh+Pib和Pi9+Pi-kh+Pita2 种组合的水稻种质资源较多。由于本研究选取的黄淮海区域的水稻种质资源较多,或许意味着这2 种抗稻瘟病基因组合较适宜黄淮海区域的生态环境,这为今后黄淮海区域的抗稻瘟病育种提供了较有价值的参考,有助于加速黄淮海区域的抗稻瘟病育种进程。
猜你喜欢
智慧农业导刊(2022年21期)2022-11-03
今日农业(2022年14期)2022-09-15
今日农业(2022年13期)2022-09-15
河北科技师范学院学报(2022年2期)2022-08-26
作物学报(2022年6期)2022-04-08
空间科学学报(2021年1期)2021-05-22
四川蚕业(2020年4期)2020-02-10
农民文摘(2019年7期)2019-01-14
中国麻业科学(2018年6期)2018-04-09
中国果菜(2016年9期)2016-03-01
