
酰化黄酮苷类化合物研究进展
2022-02-13 14:52钟朝敏徐念智
湖北农业科学 2022年23期
钟朝敏,徐念智,李 兵
(广西中医药大学药学院,南宁 530200)
黄酮类化合物有多种生物活性,在食品、保健品和化妆品等行业均有广泛的应用前景。有研究表明黄酮糖苷类化合物经酰化修饰后,其脂溶性和水溶性有明显提高,通过改变理化性质还能提高其生物利用率[1]。该类化合物具有低毒、高活性等优点。目前,利用特异性酰化来修饰黄酮苷类化合物以提高其稳定性和生物活性越来越引起研究者的兴趣。结合现有文献,本研究总结了酰基化黄酮苷类化合物的国内外研究进展,从酰化结构种类和生物活性方面展开讨论,以期为该类化合物的深入研究提供理论基础和研究思路。
1 酰化黄酮苷类化合物类型及结构特点
酰基化是修饰药物结构及功能的有力手段,大量研究报道了黄酮类化合物主要是采用酶催化的方法实现酰基化修饰[2]。该类化合物的表面装饰主要以乙酰基、对香豆基、没食子酰基等修饰呈现,其中对香豆基的酰基化修饰是最重要的形式[3]。有研究表明[4],花青素酰基化后结构更稳定,与多酚聚集一起可呈现出更蓝的花色,促进脂肪及类脂代谢的同时,还增强了抗菌和抗氧化等生物活性。从Afendi等[5]总结的黄酮类物质信息库发现,酰基化黄酮类化合物主要存在于黄酮苷、黄酮醇类和花青素中,母核结构类型如图1 所示。本研究主要对乙酰基、咖啡酰基、香豆酰基、没食子酰基等几类酰化单元进行归纳。
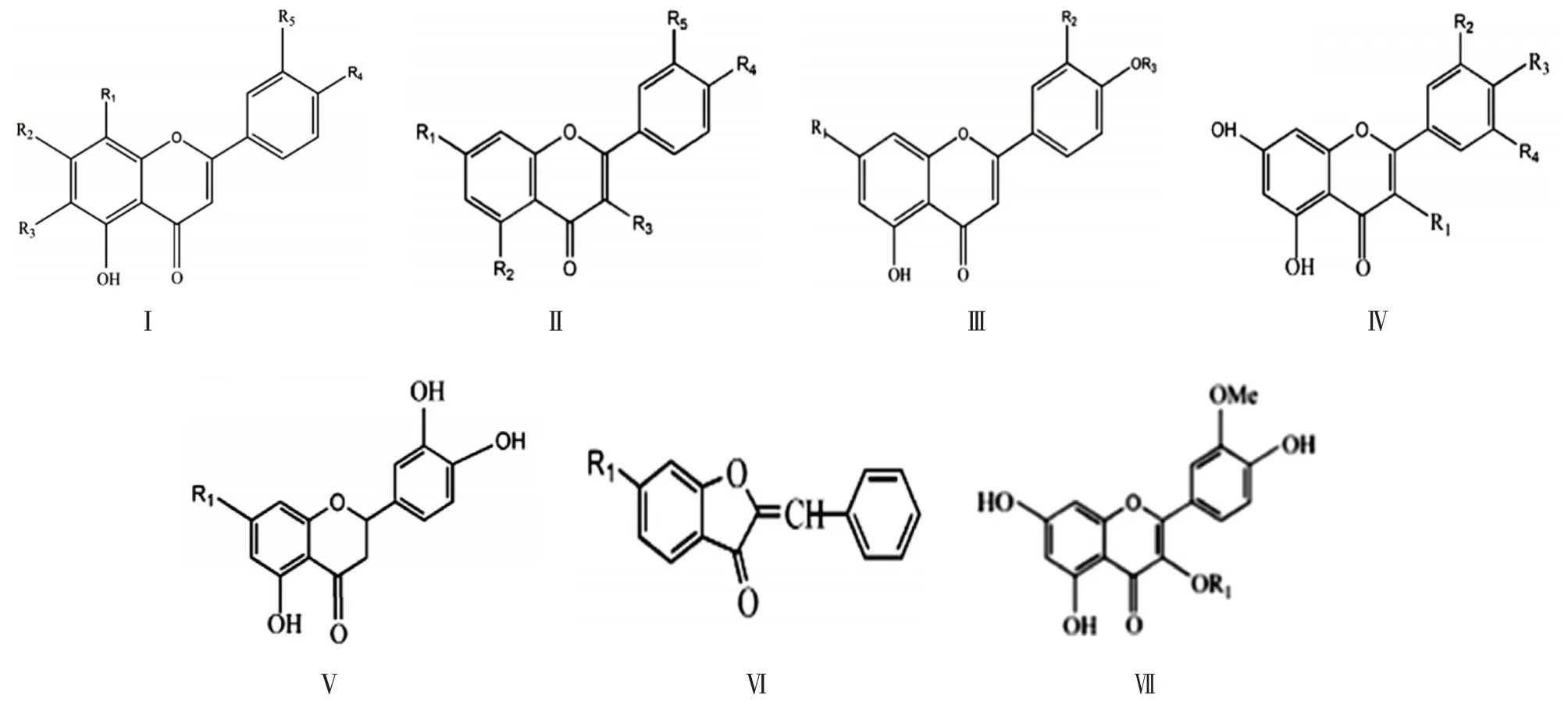
图1 酰基化黄酮苷母核类型
1.1 乙酰基黄酮苷
Lee 等[6]首次从春黄菊地上部分的甲醇提取物中分离出2 种新的乙酰化黄酮醇二糖苷。雷军等[7]从点地梅95%乙醇提取物的正丁醇萃取部分分离出4 种乙酰基黄酮醇苷,糖基结构均为吡喃鼠李糖苷。还有大量研究报道了从不同植物中分离得到该类成分,如地稔、狗脊、桑叶等,乙酰基黄酮苷的来源及化学式[6-19]见表1。
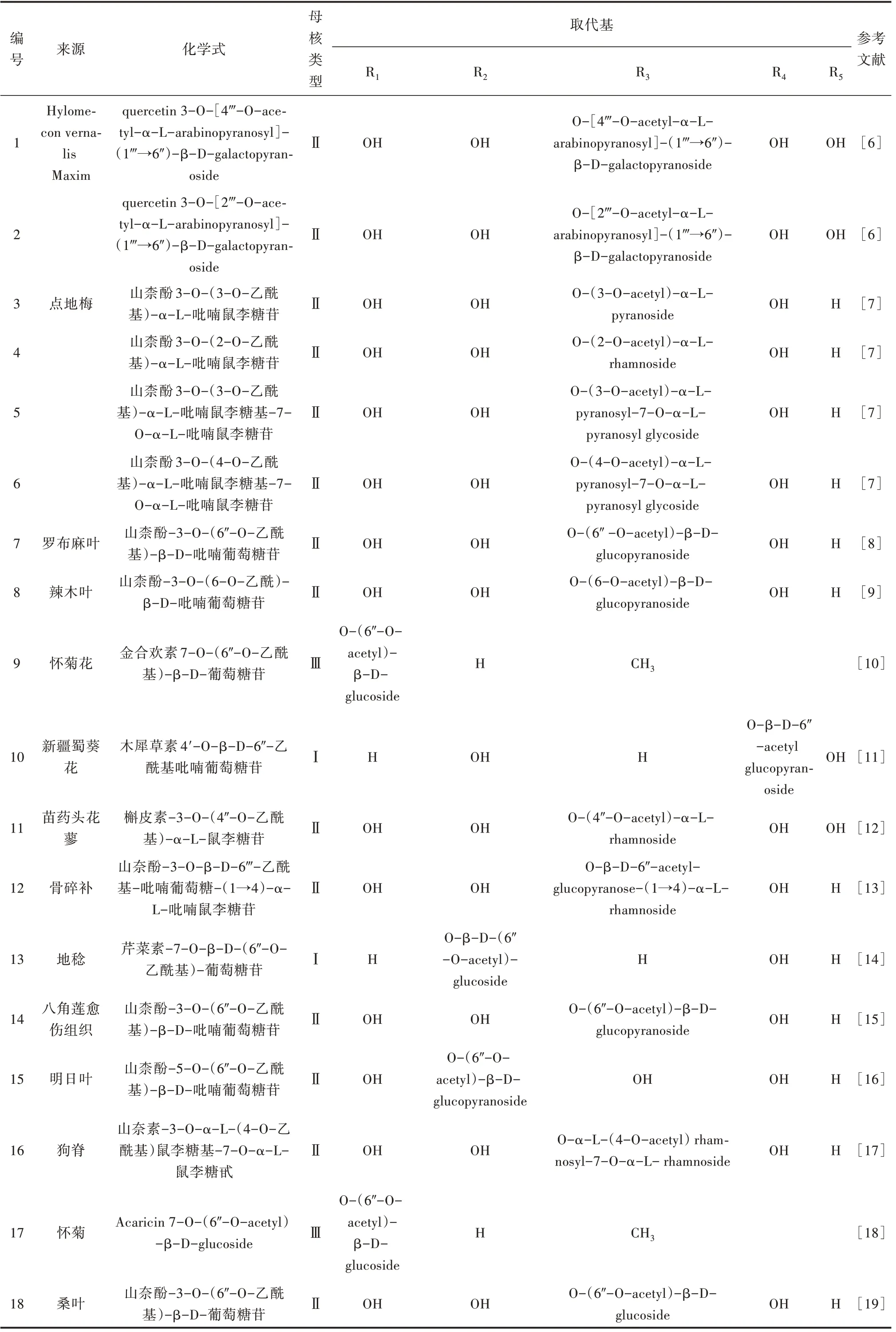
表1 乙酰基类黄酮苷来源及化学式
1.2 咖啡酰基黄酮苷
张洁等[20]、潘争红等[21]从裸花紫珠中发现了3种以木犀草素为母核的酰基化黄酮苷。Xi 等[22]从鼠曲草乙酸乙酯部分分离出3 种新的咖啡酰基黄酮醇苷,除母核不一样外,其糖基、酰基的基团和连接均相同。部分学者还从绿萝叶、牡荆叶、粘毛蓼、盒果藤等植物中发现此类黄酮苷,咖啡酰基类黄酮苷来源及化学式[20-29]见表2。
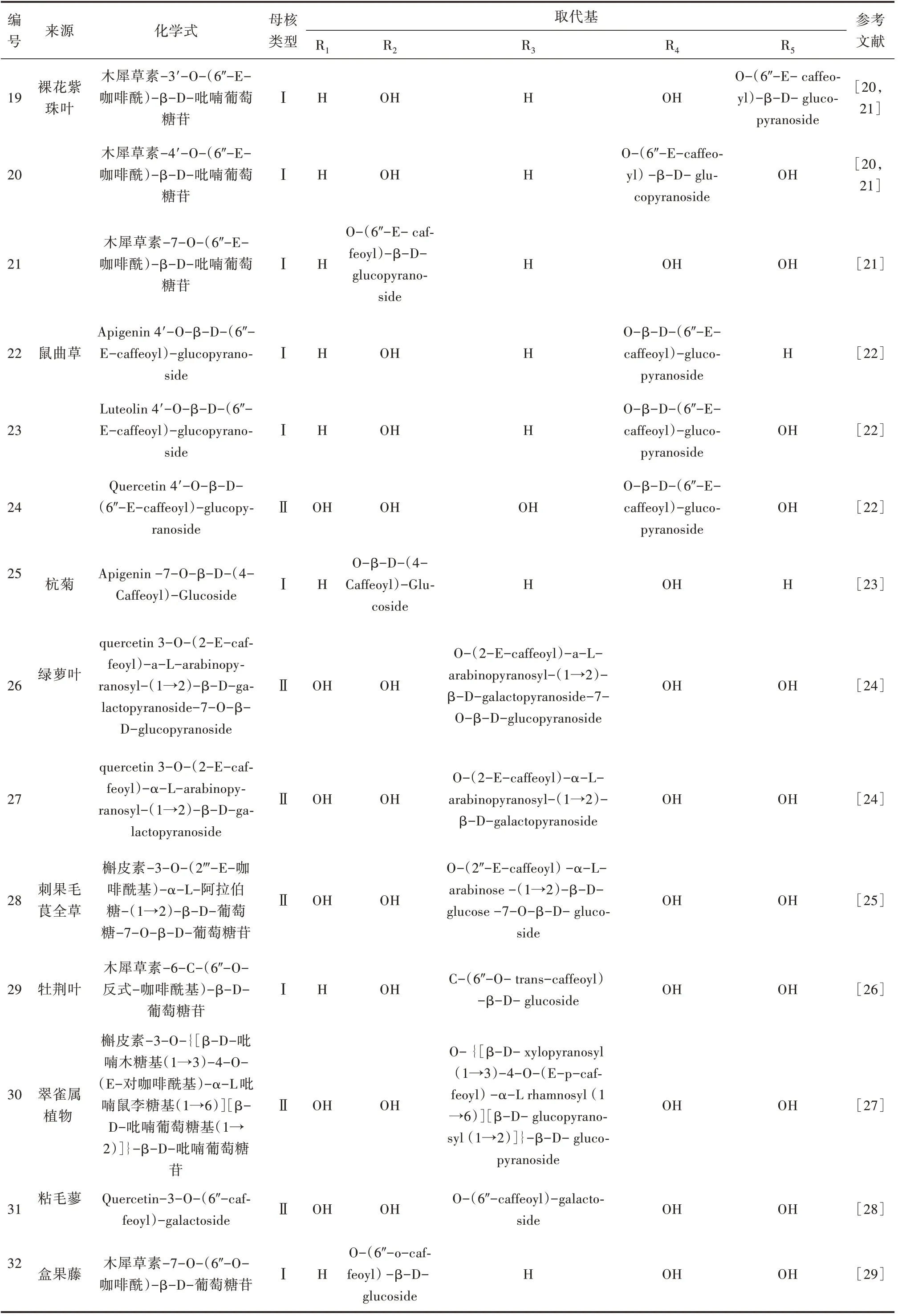
表2 咖啡酰基类黄酮苷来源及化学式
1.3 香豆酰基黄酮苷
李向日等[30]从零陵香的70%乙醇提取物中发现一种新的酰化黄酮四糖苷成分,命名零陵香黄酮A。Tian 等[31]、Lu 等[32]从FBT 中分离出3 种新的酰基糖苷黄酮化合物,包括2 种槲皮素酰基糖苷和1 种山奈酚酰基糖苷,分别命名为茶黄素苷E、茶黄素苷F、茶氨酸氨基糖苷A。大量研究发现,该类成分还存在于蜀葵花、地锦草、月季花等植物中,香豆酰基类黄酮苷来源及化学式[30-53]见表3。

表3 香豆酰基类黄酮苷来源及化学式
1.4 阿魏酰基黄酮苷
潘争红等[21]从裸花紫珠叶的95%乙醇提取物中分离出一种木犀草素-7-0-(6″-E-阿魏酰)-β-D-吡喃葡萄糖苷。吴碧灵等[25]从刺果毛茛全草的乙醇提取物分离得到槲皮素-3-O-(2‴-E-阿魏酰基)-β-D-槐糖-7-O-β-D-葡萄糖苷。师帅等[54]从金莲花体积分数为60%的乙醇溶液提取物中分离得到一个apigenin-8-C-(2″-O-feruloyl)-β-D-glucoside。阿魏酰基黄酮苷相应的植物来源及化学式[21,25,36,38,43,51,54-56]见表4。

表4 阿魏酰基类黄酮苷来源及化学式
1.5 没食子酰基黄酮苷
李梦云[57]首次从水蓼全草中分离出一种酰基化黄酮苷,结构鉴定为槲皮素-3-O-(2″-O-没食子酰基)葡萄糖苷。石娟等[58]从鹿衔草的全草中发现一种没食子酰黄酮苷(2″-O-没食子酰基金丝桃苷)。没食子酰基黄酮苷相应的植物来源及化学式[12,38,40,46,57-64]见表5。
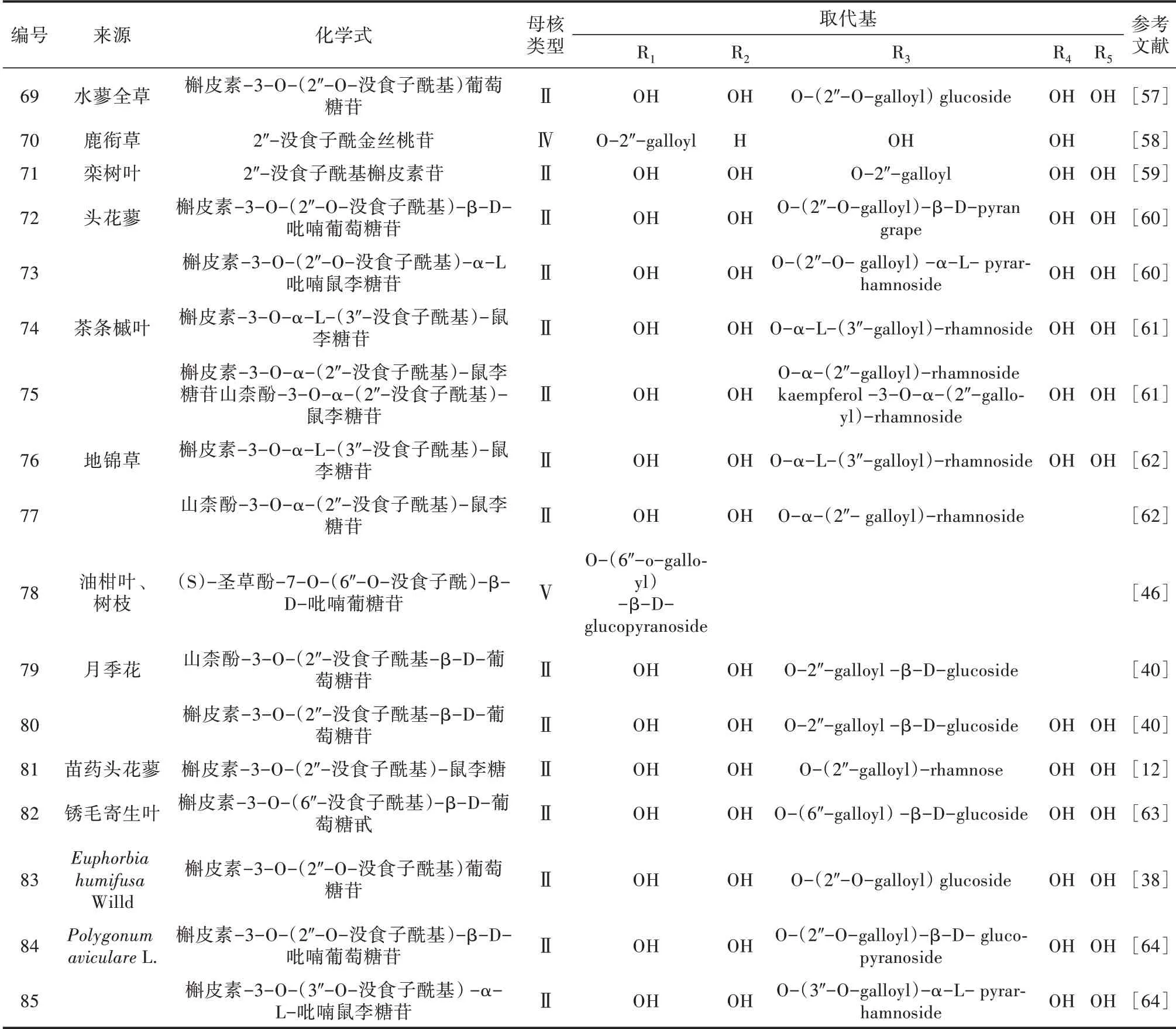
表5 没食子酰基类黄酮苷来源及化学式
1.6 其他类型的酰基黄酮苷
陈月红[65]从金盏银盘中鉴定出一种丙酰基橙酮苷,其结构为Z-6-O-(6″-丙酰基-β-D-吡喃葡萄糖基)-6,7,3′,4′-四羟基橙酮。杨小凤等[66]从栾树种子中分出2 种新的酰基化黄酮五糖苷,分别命名为栾树酮A、栾树酮B。还有许多科研者研究报道了该类化合物,相应产物来源及化学结构[9,55,65-74]见表6。

表6 其他酰基类黄酮苷来源及化学式
2 酰化黄酮苷的生物活性
2.1 抗病毒活性
黄酮类化合物是天然产物中抗病毒的典型代表,大量的研究表明该类化合物具有广泛的抗病毒活性,主要涉及抗HIV、抗流感病毒、抗HBV、抗HCMV 等。酰化黄酮苷类化合物在抗突变、抗增殖、抑制癌细胞入侵等方面具有较好的效果,近年来受到人 们 的 普 遍 关 注。Tian 等[75]在CamellikaempferosideA 存在或不存在的情况下,以MTT 测定MCF-7和丙二醛-甲基溴-231 细胞的活性,结果表明,CamellikaempferosideA 对恶性细胞MCF-7 和MDAMB-231 细胞的生长有相似的抑制作用。田瑛[76]首次发现从地锦草抗HBV 活性部位得到的2 种酰基化黄酮单糖苷具抗HBV 活性,通过研究活性部位中酰基化黄酮苷与体外抗HBV 活性的影响,发现黄酮类化合物结构中的糖基数量及母核结构与其抗HBV活性有密切关系,表现为黄酮单糖苷>黄酮二糖苷,芹菜素>木犀草素>槲皮素;而经酰基化黄酮糖苷有较好的体外抗HBV 活性,表现为没食子酰基>阿魏酰基>香豆酰基。

续表3
2.2 抗氧化活性
植物中的黄酮类化合物,可有效清除机体内氧自由基,防止细胞退化和衰老,是一种广泛存在的抗氧化剂,也是黄酮类化合物重要的生物活性。有学者研究了黄酮类化合物的体外抗氧化功能,发现其对自由基、超氧阴离子、过氧化氢等有害物质均有较好的清除效果[77]。陈月红[65]对金盏银盘中得到的丙酰基橙酮苷进行抗氧化活性方面的研究,发现其在浓度为0.025~0.100 mg/mL 时对OH-的清除率较高,且清除率与浓度呈正相关。酚羟基是黄酮类化合物清除自由基的主要活性基团,研究发现,在一定程度上增加酚羟基的数目,抗氧化活性也会相应增强,分子中的羟基数目、位置与活性有直接关系。Manir 等[78]通过1,1-二苯基-2-苦基肼(DPPH)自由基测量,比较4 种槲皮素酰基糖苷Camelliaquecetiside A-D的抗氧化性能,发现Camelliaquecetiside A清除DPPH 自由基的能力最强,IC50=(19.8±0.1)μmol/L。李梦云[57]对水蓼中分离出的槲皮素-3-O-(2″-O-没食子酰基)葡萄糖苷进行清除DPPH 自由基测试,发现其抗氧化能力强于阳性对照的维生素C,说明水蓼可以作为一种较好的天然抗氧化剂。
2.3 抗炎活性
有研究报道,黄酮类化合物发挥抗炎作用的主要场所在巨噬细胞,该类化合物通过抑制核转录因子κB(nuclearfactorκB,NF-κB)信号通路和下调促炎标记物的表达来抑制巨噬细胞的炎性反应[79]。张静[72]利用LPS 诱导的BV-2 小胶质细胞活化模型,对从夏至草中分离得到的山奈酚和槲皮素酰化黄酮苷进行体外抗炎活性筛选,使用NO 试剂盒检测一氧化氮(NO)的含量,发现山奈酚戊二酰基糖苷能不同程度地抑制脂多糖诱导的BV-2 细胞释放NO。
2.4 抗菌活性
李梦云[57]通过对水蓼中分离出的槲皮素-3-O-(2″-O-没食子酰基)葡萄糖苷进行抑菌活性测试,发现其对金黄色葡萄球菌最小抑菌浓度最小,为31.2 μg/mL,表明抑菌效果最好,当槲皮素上3-OH基团被糖苷取代后抑菌活性有所下降且对不同的细菌影响效果不同,进一步证明了水蓼具有抑菌活性的物质基础。Gatto 等[80]的研究发现,3-O-acylquercetines 在100 mg/mL 时具有较广的抗菌谱,对选定的革兰氏阳性菌株(金黄色葡萄球菌、伊凡诺氏利斯特氏菌等),革兰氏阴性菌株(大肠杆菌、福氏志贺氏菌、鼠伤寒沙门氏菌等)和酵母菌(白色念珠菌、光滑念珠菌)均有显著抑制作用,且其抑菌效果和亲脂性都比槲皮素好,因此可更好地应用在皮肤或黏膜上的抑菌治疗。2005 年,赵海誉[81]发现北葶苈子中的苄基芥子油有广谱抗菌作用,且对酵母菌等20 种真菌及数10 种其他菌株都表现出抗菌效应。
2.5 抗补体活性
补体系统是先天免疫的重要组成部分,在调节获得性免疫中也起着重要作用,二酰基化黄酮苷对补体系统有显著的抑制效果。2012 年,Xi 等[22]基于迈耶斯的改进方法,从补体的经典途径出发,研究鼠曲草中的咖啡酰基黄酮化合物对补体的抑制作用,发现黄酮类化合物主要以剂量依赖的方式表现出补体调节特性。黄酮醇-糖-咖啡酰构型的4 种酰化黄酮醇苷均显示出很强的活性,其中木犀草素4′-Oβ-D-(6″-E-咖啡酰)-吡喃葡萄糖苷具有最高的抗补体活性,其IC50为(0.045±0.005)mg/mL。这是首次对木犀草素抗补体活性进行报道,同时也初步证明了该植物是一种有效的补体抑制剂。孙连娜等[82]公布了酰化黄酮苷化合物及其在制备补体抑制剂药物上的发明,该发明经体外试验证实了鼠曲草中4 种酰化黄酮苷在对补体系统的经典途径激活所引发的细胞溶血均有抑制作用,且各单体化合物的活性比鼠曲草总提取物的活性强,表明酰化黄酮苷是一类很好的补体抑制剂,且其有效浓度低,可作为活性成分用来制备新型的抗补体药物。
2.6 其他生物活性
随着分离提取及纯化技术的提升,酰基化黄酮的生物活性研究得以更加深入,除以上归纳的作用,该化合物还有很多种生物活性,如抑制乙醇脱氢酶活性、预防骨骼肌萎缩、抑菌、降血糖血脂和减肥等。HMG-CoA 还原酶是肝细胞合成胆固醇过程中的限速酶,催化生成甲羟戊酸(MVA),抑制HMG-CoA 还原酶从而阻碍胆固醇的合成。Lu 等[83]通过分子对接模拟分析了AGFs 及其代谢产物对α-葡萄糖苷酶和HMG-CoA 还原酶的抑制能力,结果表明,AGFs的抑制活性显著受到酰基结构、糖基数目和构象的影响,并发现从茯砖茶中分离的酰基化合物对α-葡萄糖苷酶和HMG-CoA 还原酶具有很强的抑制活性,其中Camelliquercetiside F 抑制HMG-CoA 还原酶活性最高,与含酰基的代谢物相比,不含酰基的代谢物一般抑制能力较低,分析原因可能是酰基结构参与稳定疏水力的形成,提高了生物活性化合物的生物活性。2020 年,Hsieh 等[84]研究发现Teaghrelins与ghrelin 一样,都是通过下调Atrogin-1andMu RF-1泛素E3 连接酶的表达量以减缓地塞米松引起的肌管直径减小,表明其对骨骼肌萎缩有保护作用。
3 小结与展望
近年来,黄酮类化合物广泛地应用于医药、食品等行业中,然而,由于该类化合物分子结构导致其溶解性差,生物利用率受到限制。因此,在不同植物来源的黄酮类化合物的基础上,对该类化合物进行酰基化修饰,不仅可以改变其溶解性,促进脂肪及类脂代谢,提高其生物利用度,而且还能增加该类化合物的抗菌及抗氧化等生物活性。随着酰基化黄酮苷的进一步研究,未来将会有更多的成分及功效被发现。通过查阅并分析国内外的相关文献,发现酰化黄酮苷类化合物在天然植物中广泛分布,目前该类产物的植物来源已有大量报道,但在分离提取以及生物活性方面的研究报道仍较少,期待今后有更多相关领域的研究。
猜你喜欢
保健与生活(2021年13期)2021-07-14
中国果树(2020年2期)2020-07-25
天然产物研究与开发(2018年11期)2018-11-30
中国洗涤用品工业(2017年2期)2017-04-16
当代化工研究(2016年2期)2016-03-20
中国洗涤用品工业(2016年2期)2016-02-28
华东理工大学学报(自然科学版)(2015年4期)2015-12-01
医学研究杂志(2015年9期)2015-07-01
云南中医学院学报(2014年2期)2014-11-07
应用化工(2014年5期)2014-08-08
