
乌鲁木齐达坂城山区地衣物种多样性研究
2022-02-10 12:35:26李作森买吾拉江衣沙克艾尼瓦尔吐米尔
西北林学院学报 2022年1期
李作森,买吾拉江·衣沙克,艾尼瓦尔·吐米尔
(新疆大学 生命科学与技术学院,新疆 乌鲁木齐 830046)
地衣作为一种特殊的共生生物,广泛地分布在陆地生态系统的各种生境,因其结构特殊、种类多样,对环境的污染及变化非常敏感,且富含多种次生代谢物质,被广泛地用于环境质量监测,医药、香料和染料生产等多个领域,引起了各国学者的广泛重视[1-3]。近年来随着全球变化,生态环境变差,地衣栖息地受损,地衣物种多样性不断丧失,地衣物种多样性研究已成为该领域的重点和热点[4]。国外有关地衣资源调查和地衣物种多样性编目研究较早,17世纪末和18世纪初欧洲、北美等地区地衣区系及系统分类方面的研究已开始。O.Galløe[5-6]在“丹麦地衣生态学”中系统描述了丹麦的地衣种类及其分布特征。近年来众多学者在不同地区开展了地衣种类组成研究,取得了一系列成果[7-9]。据估计,全世界已发现的地衣种类有2.8万种,其中子囊菌门、担子菌门与不完全地衣分别占98%、0.4%与1.6%,我国地衣型真菌理论上应有3.6万种,但已知的地衣型真菌种类仅有3 000多种,占理论估计数的8.5%[10-17]。因此,研究我国地衣物种多样性,查明地衣资源研究潜力巨大且紧迫。
有关新疆地衣的研究主要集中在天山山脉的博格达峰的北坡、阿尔泰山脉和昆仑山脉等地区的地衣区系地理、地衣群落生态学特征、岩面生地衣、壳状地衣和大型地衣物种多样性及群落结构,地衣物种分布与环境因素之间的关系等方面[9,18-24],但有关博格达峰南坡的达坂城山区地衣多样性的研究尚未见报道。本研究根据实地调查和室内鉴定数据,分析了达坂城山区地衣多样性及其分布特征,以期为该地区地衣资源保护提供参考。
1 材料与方法
1.1 研究区概况
达坂城区位于天山东段最高峰博格达峰南部(44°33′N、88°23′E),距乌鲁木齐市区86 km,西临大湾乡和托里乡,东南与吐鲁番市、托克逊县交界,北接芦草沟乡和阜康市、吉木萨尔县,总面积5 881.8 km2。达坂城属中温带大陆性干旱气候,冬夏冷热悬殊,昼夜温差大,干燥少雨是主要的气候特征,光热资源丰富,无霜期135~150 d。年平均气温为6 ℃,而最热月7月的平均最高气温有27.8 ℃,极端最高气温可达37.5 ℃;最冷月1月的平均最低气温为-14.6 ℃,极端最低气温达-30.9 ℃。年平均降水量为70 mm,年平均风速6.3 m·s-1。达坂城境内有著名的博格达峰、天池、湿地等生态景观,生境独特,自然生态景观丰富多样[25-26]。
1.2 研究方法
野外调查和采样在达坂城区西沟乡、东沟乡、阿克苏乡(黑沟村、黑沟冰湖、高崖子牧场)等6个地点进行。根据研究区的自然环境特征,在海拔1 700~3 400 m采集地衣标本,记录采集点的海拔高度、经纬度、基物类型等信息。在实验室用MOTIC光学体视显微镜和日本OLYMPUS显微镜观察地衣体的外部形态和子囊、子囊孢子和侧丝等内部解剖结构;并结合化学显色反应(chemical spot test)鉴定地衣种类。对有些分类较难的地衣标本进行鉴定时,采用了显微化学结晶法(MCT)和薄层色谱法(TLC),对具有分类意义的地衣化学成分进行了测定[19]。
2 结果与分析
2.1 地衣种类组成
初步鉴定达坂城山区的地衣共68种,隶属11目23科38属(表1)。

表1 达坂城山区地衣名录
达坂城山区茶渍目地衣在种、属和科的数量上占优势地位,它含有7科14属25种、分别占该地区地衣科、属和种总数的30.43%、36.84%、36.76%。其次为黄枝衣目和微孢衣目。11个目中排名前3的优势目含的种数占地衣总种数的66.18%,其余8个目占地衣总种数的33.82%(表2)。
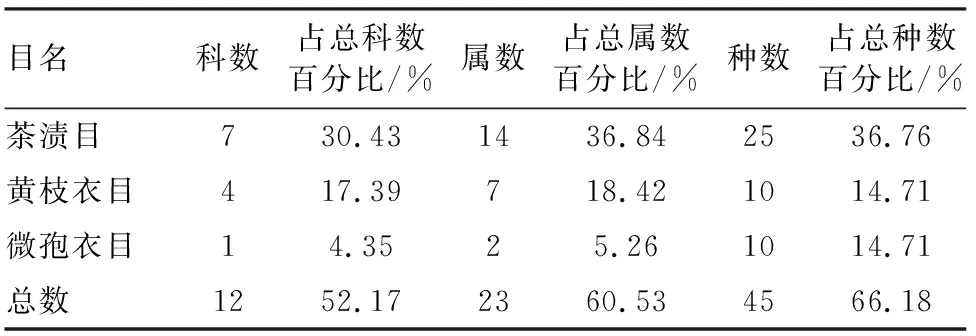
表2 达坂城山区地衣优势目组成
壳状(壳状,鳞片状)地衣在本研究区占绝对优势,共有41种,占地衣总种数的60.29%,其中34种壳状地衣分布在岩面上,只有7种分布在地面(地面生,苔藓上生,苔藓伴生)。其次是叶状地衣,共有21种,占地衣总种数的30.88%,其中14种分布在岩面,7种分布在地面(地面生,苔藓上生,苔藓伴生)。枝状地衣共有6种,占地衣总种数的8.82%。
达坂城山区地衣55种有子囊盘,可进行有性繁殖,占该地区种数总数的80.8%,而通过粉芽、裂芽、小裂片、分生孢子壳等繁殖体进行无性繁殖的地衣有21种,有8种地衣既能有性繁殖又能无性繁殖,不同的繁殖方式保证了该地区地衣广泛的分布。就共生藻类型而言,达坂城山区地衣所含共生藻大多为共球藻和球形绿藻,蓝藻地衣仅有7种,绿藻对干旱环境比较耐受,与该地区干旱的气候条件相适应。

续表1
2.2 地衣科属的构成
达坂城山区地衣种数在5种以上的科有4科,共32种,其种数占地衣总种数的47.06%。其中梅衣科地衣有4属10种,分别占该地区地衣属和种总数的10.53%和14.71%;微孢衣科地衣有2属10种,分别占地衣属和种总数的5.26%和14.71%;茶渍科地衣有4属6种,分别占地衣属和种总数的10.53%和8.82%;大孢衣科地衣有3属6种,分别占地衣属和种总数的7.89%和8.82%(表3)。
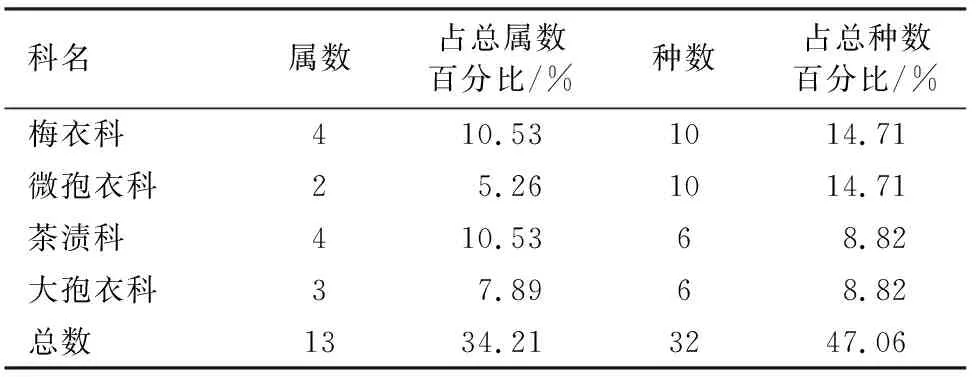
表3 达坂城山区地衣优势科组成
另外还发现,研究区石蕊科、粉衣科、腊肠衣科、胶衣科、地卷科、盾衣科、文字衣科7科地衣仅有单属、单种,珊瑚枝科、蜈蚣衣科、鳞网衣科、树花衣科、柄盘衣科、地图衣科、瓶口衣科、石耳科8科仅有2种。含5种及以上地衣种的优势属共2属,包含16个地衣种,占该地区地衣总种数的23.53%。其中微孢衣属、黄梅属的种分别占该地区地衣总种数的13.24%、10.29%(表4),5种以下且3种及以上的地衣属有3属,分别为平茶渍属(4种)、网衣属(3种)和黄茶渍属(3种),其种数分别占该地区地衣总种数的5.88%、4.41%和4.41%。

表4 达坂城山区地衣优势属组成
2.3 地衣物种分布特征
达坂城山区的地衣主要为岩面生和地面生地衣。其中分布在岩石上的岩面生地衣共有49个种,隶属于23属、16科,分别占该地区地衣种、属和科总数的72.06%、60.53%和69.57%。岩面生地衣种类中,微孢衣属(9个种)、黄梅属(5个种)和平茶渍属(4个种)占优势,它们包含的种数分别占该地区地衣总种数的13.24%、7.35%、5.88%。分布在地面生境中的石浮土生、苔藓生、地面生地衣种类有19个,占种总种数的27.94%,隶属于17属、13科,分别占该地区地衣属、科总数的44.74%和56.52%。
达坂城山区地衣的分布具有垂直地带性差异(表5)。在海拔1 100~1 600 m地带,生境主要由裸露岩石和贫瘠土壤构成,气候干旱、光照强烈。分布在该地带的地衣物种最多,共有47种,隶属27属、19科,分别占该地区地衣种、属和科总数的69.12%、71.05%、82.61%。其中,微孢衣属、黄梅属的种类最多。从基物类型得知,岩面生地衣有37种、占该地区地衣总种数的54.41%,地面生地衣10种。在海拔2 400~2 900 m地带,生境同样以裸露的岩石和稀疏的草丛组成,光照强、但该地带的气候相对湿润,人为干扰较少、降雨量较多。分布在该地带的地衣共有27种,隶属18属、15科,分别占该地区地衣种、属和科总数的39.71%、47.37%、65.22%。其中岩面生地衣有21种,地面生地衣有6种。在海拔2 900~3 400 m的阴坡,生境主要以裸露的岩石和草甸为主,草甸覆盖度高,气候潮湿、光照弱,几乎没有人为干扰。分布在该地带的地衣种共有25种,隶属21属、18科,分别占该地区地衣种、属和科总数的36.76%、55.26%和78.26%。其中,岩面生地衣17种,地面生地衣8种。分布在达坂城山区不同海拔区域的地衣中,43种地衣仅分布在低,中或高海拔区域,25种地衣在不同海拔区域有分布。
达坂城山区海拔和岩石不同坡向是导致岩面生地衣种类差异的主要因素。但嗜氮的丽石黄衣(X.elegans),嗜氮和嗜酸的地图衣等种类广泛的分布在不同海拔的岩石表面,体现了其对不同生境的高度适应能力。包含球形绿藻的包氏微孢衣、被膜微孢衣、莲座微孢衣等种类分布在光照较强、干旱的岩石表面,而含有蓝藻的地衣主要分布在较湿润土壤(地面,苔藓生)基质。具有粉芽的种类分布在岩石的裂缝、岩石侧面来保护粉芽,防止受到雨水和风的破坏,具有裂芽和子囊盘的种类分布在岩石表面的区域,说明干旱地区岩面生地衣在适应干旱环境的繁殖方式基本保持一致[22]。
3 结论与讨论
研究区地衣共有68种,隶属11目23科38属。茶渍目、黄枝衣目和微孢衣目为该地区地衣优势目,种数占该地区地衣总种数的66.18%,优势目显著,单属科、单种属情况明显,且寡种科(小于等于2种)有15科,占该地区地衣科数的65.22%,表明该地区环境异质性较高,物种适应环境导致物种分布差异,形成了较高物种多样性[27],同时,大量的单属种、单属科、寡种科(小于等于2种)反映出博格达峰南坡生态系统又较脆弱。
达坂城山区岩面生地衣具有绝对优势,共49种,占总种数的72.06%,且岩面生地衣大部分为耐旱型的壳状地衣,地面生(地面生,苔藓生,苔藓伴生)地衣19种,仅占总种数的27.94%,这主要与本研究区的生境有关,大量的裸露岩石为岩面生地衣提供了绝佳的生长环境,尤其为干旱区壳状地衣提供了稳定的基物和栖息环境。地衣能广泛生长在多种基物上并形成稳定的群落,但很多物种对基物具有高度的专一性[28]。本研究发现,除了黄梅属、泡鳞衣属和黄茶渍属地衣外,其余属的地衣只分布在一种基物上,充分体现了干旱区地衣对基物的高度专一性,与艾尼瓦尔·吐米尔等[20]在石人沟进行的研究报道一致。此外,研究区地衣共生藻类型主要为球形绿藻,能够在干旱缺水的条件下利用极少量水分进行光合作用,且不同的繁殖和传播方式能够使地衣在不同生境和不同范围内传播定殖,确保该地区地衣的广泛分布格局。地面生地衣的分布受海拔、湿度和植物盖度等多种因素共同影响[29]。在达坂城山区地面生地衣主要分布在低海拔的山地草原带,在亚高山-高山草甸带随着高等植被盖度的增加,地面生地衣无法获得充足的光照,其盖度逐渐减少。
群落物种多样性的分布变化受海拔的影响,海拔高度决定着研究区的水、热,以及温度、湿度和光照等多种生态因子,是生境影响植物多样性因子的重要因子[30]。本研究发现,达坂城山区地衣的分布具有垂直地带性质差异,分布在海拔1 100~1 600 m的地衣种类最多,共有47个种(岩面生地衣37种),海拔2 400~2 900 m的地衣共有27种(岩面生地衣21种),海拔2 900~3 400 m的地衣共有25种(岩面生地衣17种)。低海拔区域裸露的岩石种类较多,为岩面生壳状和叶状地衣提供了适宜的栖息地,中海拔地区人类的活动较多,岩石种类较单一,因此地衣的种类数量相对低海拔区域较少,随着海拔的升高,生境条件恶劣,气候变化剧烈,风速较高,造成地衣分布相对较少。由此可见,地衣物种多样性不仅受自然环境因素影响,还受到人为因素的重要影响。达坂城山区自然生态系统比较脆弱[31],随着全球气候变化和旅游业的快速发展,该地区植被栖息地受到了一定程度的破坏,因此,保护区可以通过保护和恢复生态系统的多样性,从而保护生物多样性。此外,合理规划土地,科学管理研究区旅游业,尽可能减少由于人为因素带来的环境污染和栖息地破坏,是保护该区生物多样性的有效途径。
猜你喜欢
落叶果树(2021年6期)2021-02-12 01:29:10
数理化解题研究(2018年34期)2018-12-27 08:30:04
数理化解题研究(2018年16期)2018-07-12 02:46:56
北京文学(2017年12期)2017-12-12 20:23:37
体育科技(2016年2期)2016-02-28 17:06:07
吐鲁番(2015年1期)2015-11-17 19:53:54
西华师范大学学报(自然科学版)(2015年3期)2015-02-27 15:31:17
新晨(2013年1期)2014-04-03 03:40:48
中国期刊年鉴(2014年0期)2014-02-15 03:03:56
中国期刊年鉴(2014年0期)2014-02-15 03:03:54
