
基于转录组的油茶DHHC型锌指蛋白基因家族的鉴定及分析
2022-02-10 12:35:16徐嘉娟朱亚艳
西北林学院学报 2022年1期
徐嘉娟,朱亚艳,李 芳,许 杰,王 港
(贵州省林业科学研究院,贵州 贵阳 550005)
蛋白质的翻译后修饰(posttranslational modification)与其稳定性和生理功能的正确行使息息相关[1-3]。棕榈酰化修饰是众多翻译后修饰形式中最常见且可逆的脂质修饰类型,影响着蛋白质的疏水性、稳定性与活性,从而在蛋白质定位、转运及蛋白互作等方面发挥着重要调节作用[4-5],其可逆性增加了对底物蛋白调控的动态性和精确性[6],最终影响蛋白质的各种生理功能。蛋白质棕榈酰基转移酶(protein s-acyltransferases,PATs)和棕榈酰硫酯酶2种相反类型的酶动态调节棕榈酰化修饰[7]。研究表明多数DHHC型锌指蛋白家族成员具有PAT活性,该家族是一类富含DHHC-CRD(半胱氨酸富集域)的蛋白,具有C2H2型锌指结构特征,在不同物种间具有遗传保守性,高度保守的DHHC-CRD(C-X2-C-X9-HC-X2-C-X2-C-X4-DHHC-X5-C-X4-N-X3-F,X为非保守氨基酸)残基是酶活性的关键决定位点[6,8-9]。
蛋白质棕榈酰基转移酶(PAT)最早发现于酿酒酵母中,A.F.Rothetal[10]发现酵母DHHC型锌指蛋白Akr1p具有PAT活性,对维持酵母细胞形态具有重要作用。研究表明棕榈酰化修饰在真核生物中涉及广泛的细胞功能[11-12]。对植物蛋白质组学研究发现植物中存在大量的棕榈酰化蛋白,涉及植物的生长发育、器官形成、胁迫响应等众多生命活动[13]。异三聚体G蛋白是一种非常重要的信号转导成分,参与细胞分裂、病原体防御、激素信号转导等过程,在拟南芥中,其靶向质膜需要依赖棕榈酰化修饰[14]。ROP是高等植物体内广泛存在的一类信号转导G蛋白,在植物的生长发育、逆境响应、激素应答等方面具有重要调控作用,其亚细胞定位情况与棕榈酰化修饰密切相关[15]。CDPK蛋白和CBL蛋白参与植物钙离子信号传导,棕榈酰化修饰在其生物功能的正确行使过程中具有重要作用[16-17]。
油茶(Camelliaoleifera)为山茶科(Theaceae)山茶属(Camellia)常绿灌木或小乔木,是我国重要的木本食用油料树种[18-19]。目前尚未发现有关油茶DHHC型锌指蛋白基因家族的研究报道。本研究基于不同结实量的油茶叶片转录组数据,在转录组水平上挖掘油茶DHHC型锌指蛋白基因,对其进行相关生物信息学分析,为后期深入、系统地研究DHHC型锌指蛋白基因家族在油茶生长发育、器官形成、逆境响应等过程中所发挥的调控功能提供一定的理论基础。
1 材料与方法
1.1 试验材料
试验材料为种植于贵州省玉屏县茶花泉油茶基地的盛产期长林23号普通油茶无性系(气候、种植条件一致),选取生长良好、长势一致而结实量差异较大的单株,在抽梢期、花芽分化期、果实成熟期分别采集植株中上部的叶片,取样后液氮速冻-80 ℃保存备用。
1.2 试验方法
1.2.1 DHHC型锌指蛋白基因家族鉴定 普通油茶转录组数据由本团队测序获得(相关数据未发表),共拼接组装获得275 805个unigenes。利用Phytozome12基因组数据库(https://phytozome.jgi.doe.gov/pz/portal.html)下载模式植物拟南芥和水稻的DHHC型锌指蛋白基因序列,将其作为探针序列,利用本地Blast软件在油茶转录组数据中进行blast搜索比对。同时,利用关键词“protein S-acyltransferase”和“DHHC”进行直接检索,去除重复序列,筛选出油茶DHHC型锌指蛋白家族基因,利用SMART和CCD对蛋白保守结构域进行鉴定,剔除不含DHHC型锌指蛋白基因家族特征结构域的基因。
1.2.2 DHHC型锌指蛋白基因家族的生物信息学分析 对筛选出的油茶DHHC型锌指蛋白基因,利用Expasy Prot Param等生物信息学软件对其蛋白基本理化性质、保守结构域、亚细胞定位、系统进化关系等进行分析(表1)。

表1 基因生物信息学分析软件Table 1 Gene bioinformatics analysis software
2 结果与分析
2.1 油茶DHHC型锌指蛋白基因家族的鉴定
通过对油茶转录组数据库的本地blast比对检索和关键词搜索,共获得63条DHHC型锌指蛋白基因序列,通过SMART和CCD对保守结构域的筛选,获得23个具有完整DHHC型锌指结构的序列。为方便研究,按照转录组unigene中的基因ID大小顺序统一编号(CoDHHC1~23)(表2)。分离得到的23个DHHC型锌指蛋白基因大小为731(CoDHHC1)~2 994 bp(CoDHHC17)。利用Expasy ProtParam对DHHC型锌指蛋白基因家族编码的氨基酸进行理化性质分析,23个基因编码蛋白的氨基酸数量为194~725 aa。预测的理论分子量大小为 21.89~78.44 ku。等电点大小为5.18~10.01,其中只有3个CoDHHC蛋白等电点小于7,显酸性,其余20个CoDHHC蛋白等电点均大于7,显碱性,表明不同的DHHC蛋白在不同的微环境中生物学功能存在差异。不稳定指数分析表明,有10个CoDHHC蛋白的不稳定指数大于40,为不稳定蛋白,其余13个CoDHHC蛋白为稳定蛋白。平均疏水指数为-0.541~0.503,其中10个CoDHHC蛋白的平均疏水指数大于0,为疏水性蛋白,其余13个蛋白为亲水性蛋白。
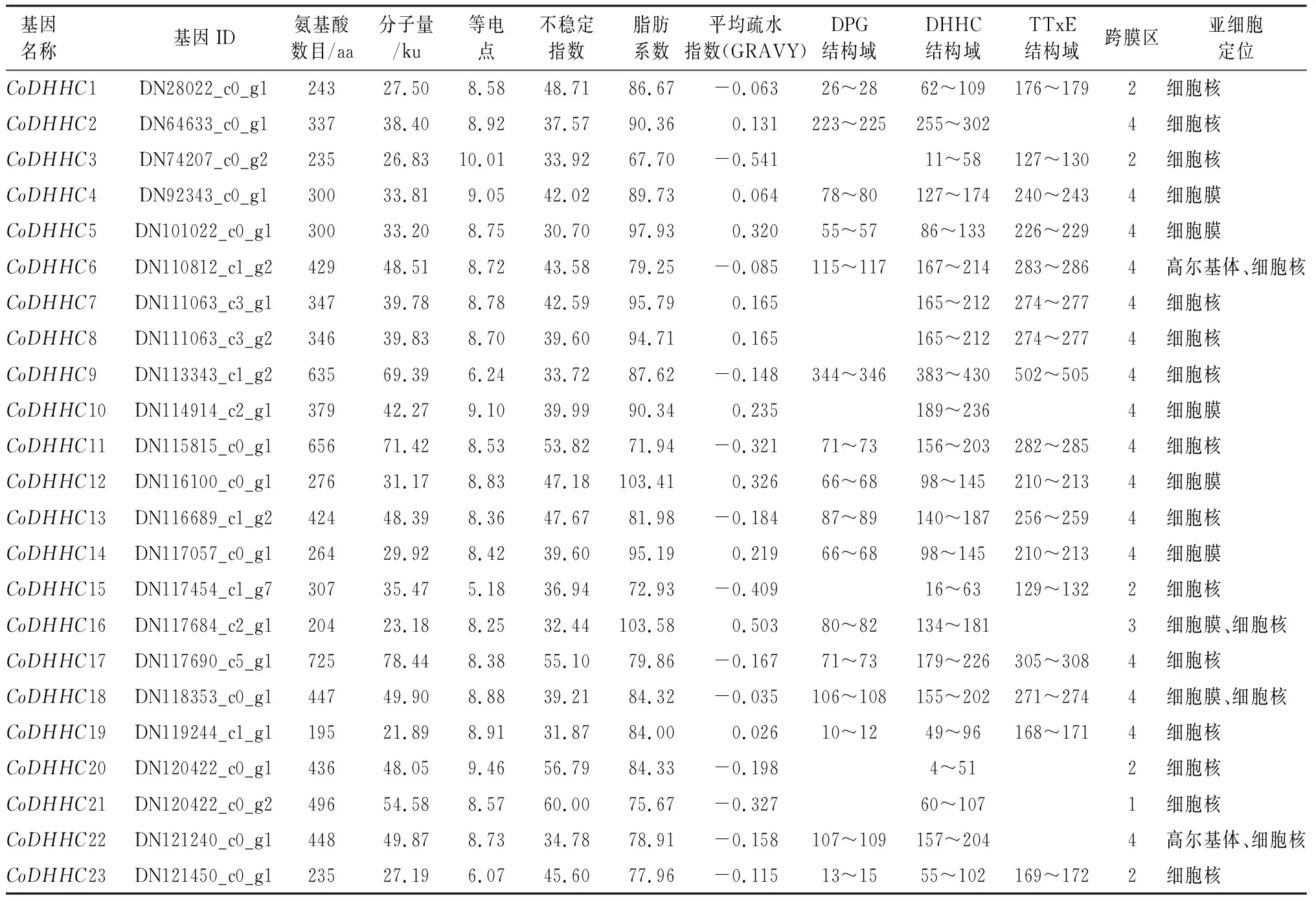
表2 油茶DHHC型锌指蛋白基因家族信息Table 2 DHHC gene family information of C.oleifera
亚细胞定位分析表明,CoDHHC1等14个蛋白在细胞核中有分布,CoDHHC4等5个蛋白在细胞膜中有分布,而CoDHHC6和CoDHHC22在高尔基体中有分布,在细胞核中也有分布,CoDHHC16和CoDHHC18在细胞膜中有分布,在细胞核中也有分布。总的来说,CoDHHC蛋白的氨基酸序列及蛋白特性变化均存在一定差异,表明油茶DHHC型锌指蛋白基因家族蛋白具有不同特性。
2.2 油茶DHHC型锌指蛋白保守结构域和基序分析
通过NCBI的CCD数据库对CoDHHC基因家族编码蛋白的保守结构域进行分析,23个CoDHHC均含有DHHC型锌指蛋白家族典型的DHHC-CRD(半胱氨酸残基聚集域)结构域,多数成员同时还含有DPG和TTxE(x为任意氨基酸)结构域,与具有蛋白质酰基转移酶活性的蛋白所特有的结构域相符。其中CoDHHC1/4/5/6/9/11/12/13/14/17/18/19/23同时含有3个结构域,占油茶CoDHHC基因家族总数的56.52%,CoDHHC2/16/22只含有DPG和DHHC结构域,CoDHHC3/7/8/15只含有DHHC和TTxE结构域,而CoDHHC10/20/21仅含DHHC结构域(表2,图1)。
利用在线软件MEME5.1.1分析CoDHHC蛋白中的保守基序(图2、图3),结果表明,23个油茶DHHC基因家族成员之间所包含的保守基序(motif)数目及种类存在一定的差异,其中CoDHHC10只含有motif 1和motif 3这2个保守基序。所有CoDHHC蛋白成员均含有motif 1,19个成员含有motif 2,19个成员含motif 3。4个成员含motif 4,5个成员含motif 5,5个成员含motif 6,20个成员含motif 7,4个成员含motif 8,4个成员含motif 9,4个成员含motif 10,2个成员含motif 11,2个成员含motif 12,2个成员含motif 13,4个成员含motif 14,5个成员含motif 15,2个成员含motif 16,2个成员含motif 17,4个成员含motif 18,3个成员含motif 19,3个成员含motif 20。出现频率越高的motif在油茶DHHC型锌指蛋白基因家族中越重要,如出现频率较高的motif 1和motif 3组成DHHC型锌指蛋白家族的特征结构域(DHHC-CRD)。
2.3 油茶DHHC型锌指蛋白基因家族进化关系分析
为更好地了解油茶DHHC型锌指蛋白基因家族各成员之间的进化关系以及生物学功能的相关性,利用MEGA7软件对筛选的23个油茶DHHC型锌指蛋白基因家族成员和拟南芥及水稻的DHHC型锌指蛋白基因家族成员构建系统进化树(图4)。结果表明,23个油茶DHHC型锌指蛋白可以划分为4个亚组,每个亚组均有油茶、拟南芥和水稻分布,GroupⅠ包含有2个DHHC型锌指蛋白基因家族成员,占家族总数的8.70%;GroupⅡ包含有8个成员,占家族总数34.78%;GroupⅢ包含6个成员,占家族总数总数的26.09%;GroupⅣ包含有7个成员,占家族总数30.43%。其中第Ⅱ亚族的CoDHHC7和CoDHHC8的同源性最高,第Ⅲ亚族的CoDHHC5、CoDHHC12、CoDHHC14的同源性较高,第Ⅳ亚族的CoDHHC13、CoDHHC18、CoDHHC22的同源性较高。总的来说进化关系较进的CoDHHC其氨基酸长度和蛋白结构较相似。
2.4 油茶DHHC型锌指蛋白基因家族的表达分析
利用课题组的油茶叶片转录组数据库(数据未发表),找到候选的23个DHHC型锌指蛋白基因对应转录本的RPKM值,采用Heml热图软件进行热图制作(图5)。结果显示,23个DHHC型锌指蛋白基因在结实量大和结实量小的单株的不同发育时期的叶片中均有不同程度的表达。CoDHHC9、CoDHHC13、CoDHHC22在油茶抽梢生长期的叶片中表达上调,CoDHHC6、CoDHHC12、CoDHHC18、CoDHHC23在花芽分化时期和果实成熟时期的叶片中表达量较高,而CoDHHC5、CoDHHC11、CoDHHC15在不同时期的叶片中表达量均较低。CoDHHC3、CoDHHC10在果实成熟时期即盛花期的叶片中表达上调,且在结实量小的单株叶片中的表达较结实量大的单株上调明显。
3 结论与讨论
基于不同结实量的普通油茶叶片转录组数据,筛选出23个DHHC型锌指蛋白基因,利用生物信息学分析方法对其理化性质、亚细胞定位、蛋白保守结构域、系统进化关系等进行分析。结果表明,23个CoDHHC蛋白均包含1个保守的DHHC-CRD结构域,主要定位于细胞核和细胞膜中,大部分蛋白具有4个跨膜区。系统进化分析发现23个DHHC型锌指蛋白与拟南芥和水稻均有不同程度的同源关系。它们在结实量大和结实量小的单株的不同发育时期叶片中均有不同程度的表达。本研究结果为进一步对油茶DHHC型锌指蛋白家族基因的克隆和功能分析奠定了基础。
近年来,随着越来越多物种全基因组测序的完成,多个物种已从基因组水平鉴定到DHHC型锌指蛋白基因,该家族成员在不同的物种中数量也不尽相同。其中小立碗藓(Physcomitrellapatens)中包含9个;裸子植物火炬松(Pinustaeda)中包含13个;拟南芥(Arabidopsisthaliana)中包含24个,水稻(Oryzasativa)有58个,杨树(Populustrichocarpa)有39个,玉米(Zeamays)有50个;双子叶植物苜蓿(Medicagotruncatula)中包含9个,马铃薯(Solanumlycopersicum)有24个[20]。本研究鉴定得到的23个DHHC型锌指蛋白基因序列,均含有DHHC蛋白家族典型的DHHC-CRD(C-X2-C-X9-HC-X2-C-X2-C-X4-DHHC-X5-C-X4-N-X3-F,X为非保守氨基酸)结构域,大部分序列还包含1个DPG和TTxE结构域及4个跨膜区,与具有蛋白质酰基转移酶(PAT)活性的蛋白质所特有的结构域相符,可能具有PAT活性,通过棕榈酰化修饰在油茶生长发育、逆境响应等过程中起重要调控作用。而部分序列如CoDHHC3缺少DPG结构域且只包含2个跨膜区,因其序列长度可能导致结构域和基序的缺失,这可能是转录组测序或序列拼接过程中的误差所导致[21-22]。
系统进化分析结果表明,23个油茶DHHC型锌指蛋白基因和拟南芥、水稻的DHHC蛋白基因被分为4个亚类,其与拟南芥和水稻的DHHC型锌指蛋白基因均具有不同程度的同源性,可能与DHHC-CRD结构域在不同物种间具有较高的遗传保守性有关。CoDHHC6、CoDHHC12、CoDHHC18、CoDHHC23在结实量较小的单株的花芽分化时期和果实成熟时期的叶片中表达量较结实量较大的单株中的表达量明显上调,可能与结实量较小的单株开花数量较多而结实量大的单株花朵数量较少有关,说明这些基因可能参与花芽分化及花朵发育过程。而CoDHHC3、CoDHHC10只在果实成熟时期的叶片中表达上调,而这一时期油茶正处于盛花期,因此它们可能参与花朵发育。Q.Wangetal[23]对拟南芥中20个DHHC型锌指蛋白基因的组织表达分析发现,AtPAT7(At3g26935)在花、成熟果荚、茎中有表达,在花中的表达量较其他组织高。徐嘉娟等[8]研究表明,LcPAT7在鹅掌楸花芽和盛花期的叶片、叶芽、花瓣、雄蕊、雌蕊中均有表达,在花瓣和雌蕊中表达丰度最高。而与AtPAT7遗传距离较近的CoDHHC6、CoDHHC18在花芽分化时期和盛花期叶片中表达上调,表明它们可能发挥相似的功能。
研究表明,PATs在真核生物中具有广泛的细胞功能,对植物的生长发育、器官形成、生殖发育及胁迫响应等生命活动具有重要的调控作用。P.A.Hemsleyetal[13]从拟南芥突变体中分离得到的DHHC型锌指蛋白基因TIPl与酵母的AKR1p高度相似性,且存在PAT活性,参与拟南芥根毛的发育及细胞的极性生长;研究发现,AtPAT10(At3g51390)能够调节CBLs蛋白在液泡膜的定位,它的功能缺失会导致拟南芥叶片变小、变矮并且不育,同时还参与植株对外界盐胁迫的响应过程[6]。水稻OsDHHC1具有PAT活性,可促进水稻分蘖、构建合理株型,从而增加水稻的产量[24];而OsDHHC13能促进清除H2O2的相关酶基因表达,调节内源性H2O2的动态平衡,对水稻的抗氧化胁迫能力起正调控作用[25]。PATs功能的发掘为植物株型构建、增产增收以及品质改良等分子设计育种工作提供了新的研究思路和基因资源。本研究利用生物信息学手段从序列同源性和结果相似性方面探讨了油茶DHHC型锌指蛋白基因家族的生物学功能,而它们在油茶生长发育、逆境信号响应等过程中的具体功能还有待系统深入的研究。
猜你喜欢
中学生天地(A版)(2023年1期)2023-02-17 00:33:04
昆明医科大学学报(2022年1期)2022-02-28 07:46:10
实用临床医药杂志(2021年13期)2021-01-10 19:52:18
阅读(快乐英语高年级)(2019年8期)2019-09-10 07:22:44
生命科学研究(2018年1期)2018-05-29 01:12:47
小火炬·智漫悦读(2018年12期)2018-03-13 09:24:52
莫愁(2017年36期)2017-12-25 05:52:36
上海农业学报(2017年3期)2017-04-10 12:39:14
山东农业工程学院学报(2016年6期)2016-12-01 05:38:19
中华胰腺病杂志(2015年5期)2015-12-08 12:18:10
