
冷季不同饲养方式对阿什旦牦牛空肠差异蛋白的影响
2022-02-08 12:09:22张莹莹张怀霞张春梅贾建磊
西南农业学报 2022年11期
张莹莹,张怀霞,谢 雯,任 昊,张春梅,陈 倩,贾建磊
(青海大学农牧学院,西宁 810016)
【研究意义】青藏高原地区海拔高,气候寒冷,牧草生长期短,造成冷季(10月至次年6月)牧草匮乏,牦牛得不到足够的供给会造成牦牛的生产效能低下甚至死亡,给牧民造成损失。为防止冬季亏损严重,有效的提高牦牛的生产效益和经济效益,牦牛的舍饲和半舍饲就成了当下的一种趋势[1-3]。反刍动物日粮需要摄入40%~70%的粗饲料,以维持正常的胃肠功能和消化道菌群环境,现代集约化饲养模式下,牦牛日粮精料超过75%[4]。长期的高精料喂养会导致亚急性瘤胃酸中毒等代谢疾病的发生,从而导致动物的机体和消化道受损,从而使牦牛的生产性能下降[5]。【前人研究进展】动物的肠道是动物消化系统的重要组成,对动物摄入的营养物质有吸收、代谢和维持机体免疫的功能。空肠是动物消化、吸收、代谢的重要场所,也是抵御有害物质进入机体对机体的损伤的重要屏障。空肠由单层上皮上细胞构成,易受到肠道内环境中pH和微生物等有毒有害物质的损伤,从而导致动物机体的损伤[6]。Jahan等[7]研究表明,高精料饲养公牛会导致瘤胃中NH3-N浓度上升,而NH3-N的含量可以直接反应瘤胃内环境的状态;Li等[8]研究表明,精粗比能够影响反刍动物瘤胃的pH,从而影响挥发性脂肪酸(VFA)和菌体蛋白的含量,菌体蛋白的浓度可以反映出瘤胃微生物利用氨态氮的能力。高精料的碳水化合物饲粮减少了其在瘤胃中的停留时间,使得大量的过瘤胃淀粉会进入到空肠,改变空肠内的发酵参数,使肠道的pH降低,挥发性脂肪酸(VFA)和脂多糖(LPS)浓度上升,引起肠道的酸中毒,从而破坏肠道上皮形态结构的完整性,使肠道屏障受损[9-10]。周力等[11]发现饲粮的精粗比可以显著影响青海黑藏羊小肠的营养物质转运载体基因的表达水平,进而影响黑藏羊小肠的代谢功能,精粗比为70∶30时效果较好;李岩等[12]发现饲粮的精粗比增加会抑制育肥后期牦牛对蛋白质的合成和对营养物质的消化,使牦牛的平均日增重和总增重降低。同时,Hristov等[13]发现长期的高精料舍饲喂养会导致动物机体代谢的紊乱,降低反刍动物的免疫能力,从而影响反刍动物的健康。此外,谢昕廷等[14]研究表明日粮精粗比会对牦牛肉的氨基酸和脂肪酸含量造成显著的影响。【本研究切入点】通过蛋白质组学研究不同饲养模式下(自然放牧和高精料舍饲)牦牛空肠的蛋白质表达差异。【拟解决的关键问题】从分子生物学的角度去探索冷季长期高精料饲养对牦牛空肠的损伤,为合理科学的进行牦牛饲养,提高生产效益和经济效益提供理论依据。
1 材料与方法
1.1 试验设计
选取海北藏族自治州天然放牧的条件下12只(4±0.5)岁左右的阿什旦公牦牛,随机分为两组,每组6只,试验期为2018年11月16日至2019年5月4日,历时180 d,前10 d为试验组牦牛预试期,后170 d为正试期。
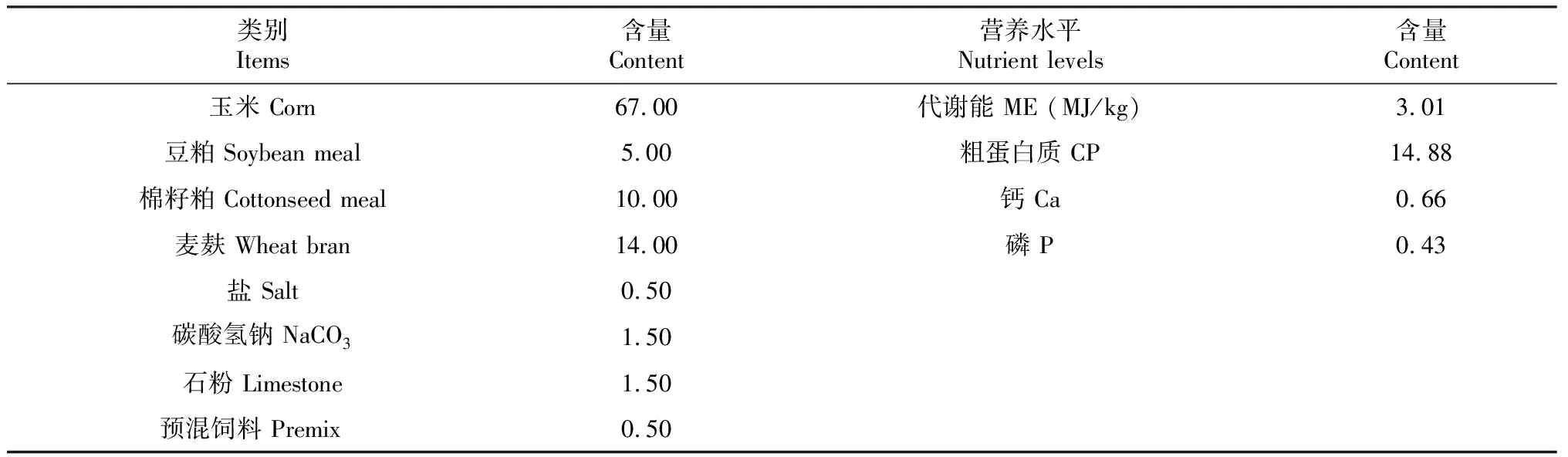
表1 试验精饲料组成及营养水平(干物质基础)
试验组通过高精料舍饲的方式饲养,对照组为放牧饲养。对照组牦牛每天6:30放牧,20:00归牧(归牧后不补饲)。试验组牦牛根据中国肉牛饲养标准(NY/T 815—2004)配制饲料,精饲料组成及营养水平见表1。粗饲料为玉米青贮和燕麦干草按1∶1比例混合,试验期间每只牦牛每日饲喂4 kg精饲料和2 kg粗饲料(干物质),每天按时饲喂2次(9:00和17:00),两组均自由饮水。
1.2 试验样品采集
饲养期结束后每组随机选取3只牦牛,在夏华畜牧产业集团空腹电击击晕后颈动脉放血屠宰,试验牦牛均已经经过国家动物屠宰检疫总局(中国,青海)批准屠宰。在屠宰后迅速采集牦牛空肠组织样本(前段、中段和后段各1 cm混合),PBS冲洗干净后装入无菌冻存管中,将样本存入液氮中带回实验室,-80 ℃保存备用。
1.3 试验方法
1.3.1 蛋白质提取 蛋白质提取方法根据贾建磊等[15]组织蛋白提取方法进行空肠组织样品总蛋白提取,每组3个重复。
1.3.2 蛋白质酶解和质谱分析 蛋白质酶解和质谱分析在北京诺禾致源生物信息科技有限公司进行。根据超滤辅助样品制备法对蛋白质进行酶解(每个样品250 μg),肽段采用流动相A相溶解后,使用EASY-nLCTM 1200 纳升级UHPLC系统进行肽段分离,肽段分离后用Q Exactive TM HF质谱仪和Nanospray FlexTM(ESI)离子源对样本进行分析,生成质谱原始数据。利用MaxQuant软件分析质谱分析的数据,并与UniProt Bostaurus数据库进行比较。
1.4 生物信息学分析
利用Blast2GO程序根据uniport数据库对所有已鉴定蛋白进行细胞组分(CC)、生物学过程(BP)和分子功能(MF)分析。信号通路分析利用KEGG mapper平台(http://www.genome.jp/kegg/mapper.html)Search pathway在线工具进行信号通路分析。利用String程序(http://string-db.org/)检索相互作用基因/蛋白质,利用Cytoscape软件对交互网络进行图形可视化和分析。
1.5 统计分析
数据测试完成后先采用Excel 2010表格进行初步统计处理,然后用SAS 9.2数据分析软件进行独立样本t检验,结果均以平均值±标准差表示,显著水平为P=0.05。
2 结果与分析
2.1 差异蛋白聚类分析
在不同饲养方式下(放牧饲养和全舍饲饲养),检测出不同饲养模式下牦牛空肠的差异表达蛋白共96种,采用FC(fold change)≥1.2且P≤0.05筛选上调表达蛋白,当FC≤1/1.2且P≤0.05筛选下调表达蛋白。样本各差异蛋白相对含量聚类热图结果(图1)表明,放牧饲养组(A)和全舍饲饲养组(B)呈现出2个明显区别的亚类,较全舍饲组相比,放牧组有17种上调差异蛋白和79种下调差异蛋白。
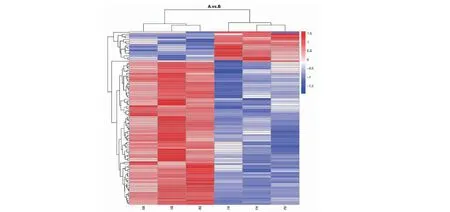
不同颜色代表蛋白质的相对丰度,深红色的代表较高的强度,蓝色代表较低的强度,聚类枝越短相似度越高Different colors indicate the relative abundance of proteins, dark red represents higher intensity and blue represents lower intensity, and the shorter the clustering branch the higher the similarity图1 放牧组(A)和舍饲组(B)差异蛋白聚类热图Fig.1 The heat map of differential protein clustering for grazing group (A) and confinedness group (B)
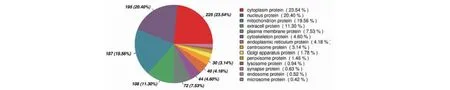
图2 差异蛋白亚细胞定位分析Fig.2 Differential protein subcellular localization prediction
2.2 亚细胞定位分析
亚细胞定位分析(图2)表明,差异表达蛋白中细胞质蛋白占23.54%,核蛋白占20.40%,线粒体蛋白占19.56%,细胞外蛋白占11.30%,质膜蛋白占7.53%,其他的所占比例较少。
2.3 生物信息学分析
2.3.1 差异蛋白GO富集分析 GO分析(图3)显示,基于细胞组分分析,差异蛋白主要在细胞膜(Membrane)、细胞质(Cytoplasm)和线粒体(Mitochondrion)中富集。 基于分子功能分析,差异蛋白主要在氧化还原酶活性(Oxidoreductase activity)、细胞间粘着连接(Cell-cell adherens junction)、腺苷酸激酶活性(Adenylate kinase activity)、结合蛋白质(Protein binding)和结构分子活性(Structural molecute activity)中富集。基于生物过程分析,差异蛋白主要在细胞凋亡(Apoptosis)、炎症反应(Inflammatory response)、细胞对脂肪酸的反应(Cellular response to fatty acids)、肌动蛋白细胞骨架重组(Actin cytoskeleton reorganization)和腺苷酸环化酶抑制G蛋白偶联受体(Adenylate cyclase- inhibiting G protein-coupled receptor)中富集。
2.3.2 差异蛋白KEGG富集分析 96种差异蛋白主要在以下20种信号通路中的富集。其中去除疾病通路显著富集的通路为:紧密连接信号通路(Tight junction signaling pathway)、NF-kB信号通路(F-kappa B signaling pathway)、MAPK信号通路(MAPK signaling pathway)、细胞因子受体相互作用(Cytokine-cytokine receptor interaction)、钙信号通路(Calcium signaling pathway)、调控干细胞的多能性的信号通路(Signaling pathways regulating pluripotency of stem cells)。可知,KEGG通路分析与GO富集分析结果相匹配。
2.3.3 蛋白质互作分析 利用String程序和Cytoscape软件对放牧饲养和全舍饲饲养的牦牛空肠的96种差异表达蛋白的互作作用进行分析,得出相应的互作信息(图5),确定了两个主要主题:蛋白质结构(Occlusion,Claudin,Zo-1,ATP1B3和HKDC1)和炎症反应(GPR41,IL-6,TNF-α和CCL5),构成了一个复杂而强大的PPI网络。
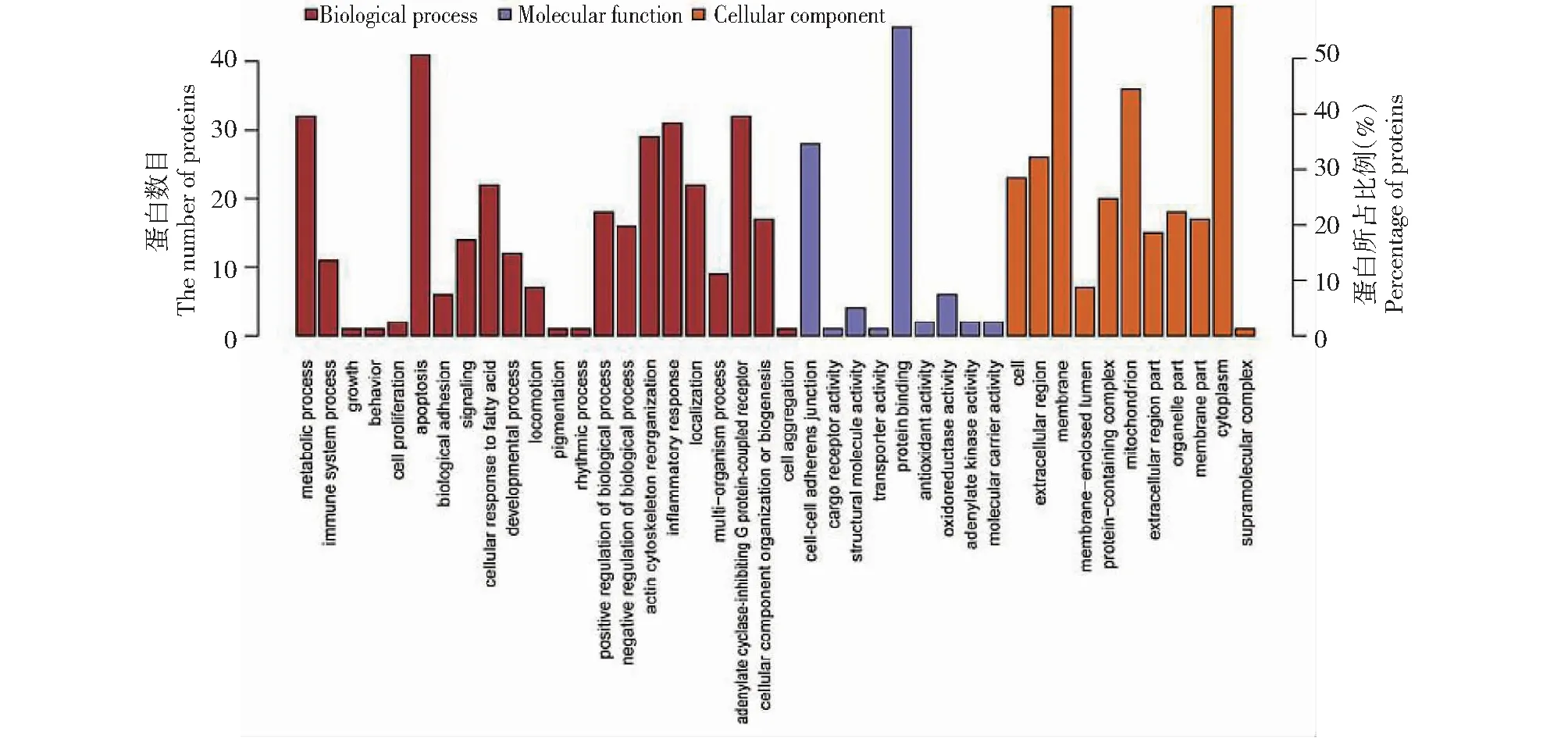
图3 GO富集分析Fig.3 GO enrichment analysis
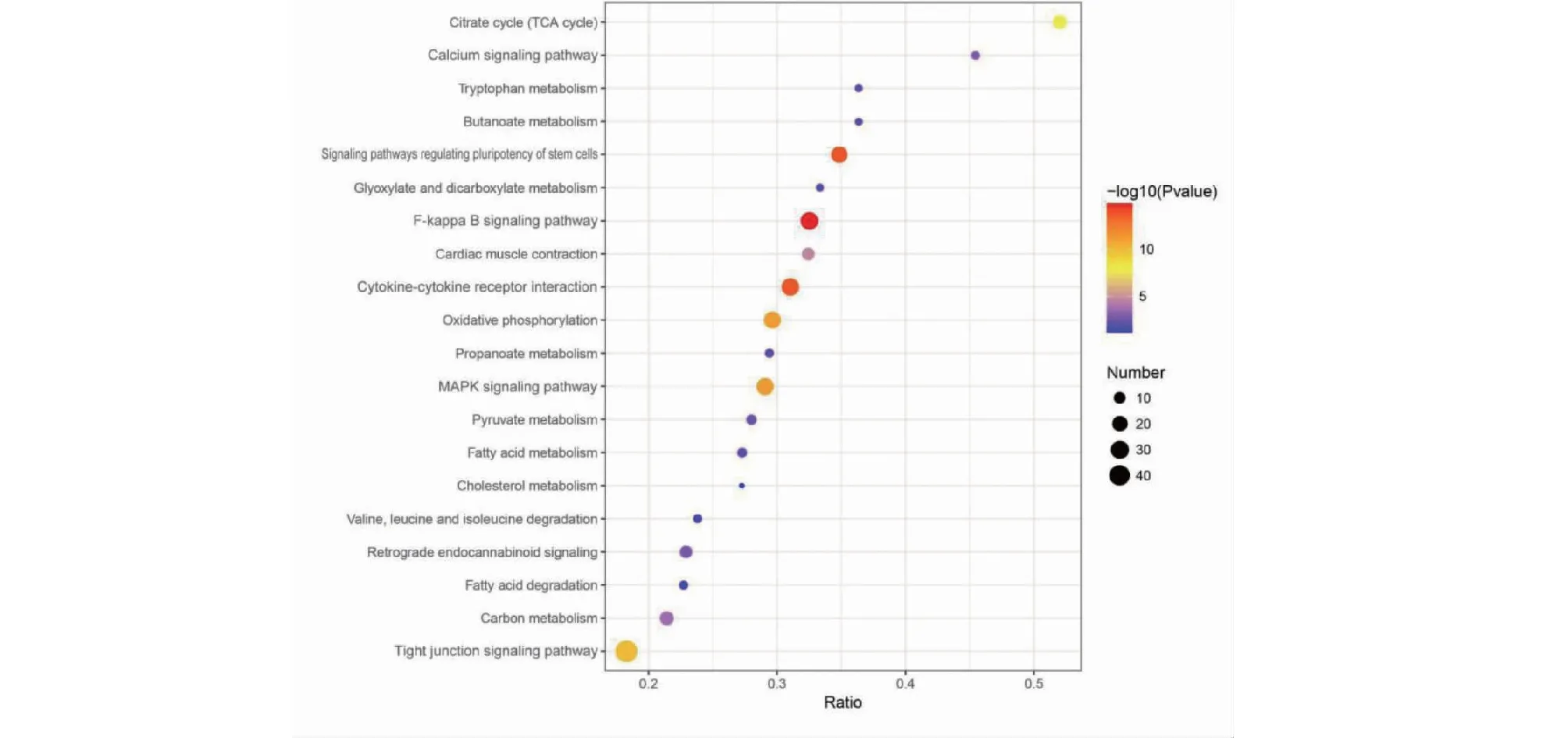
图4 KEGG富集分析Fig.4 KEGG pathways enrichment analysis
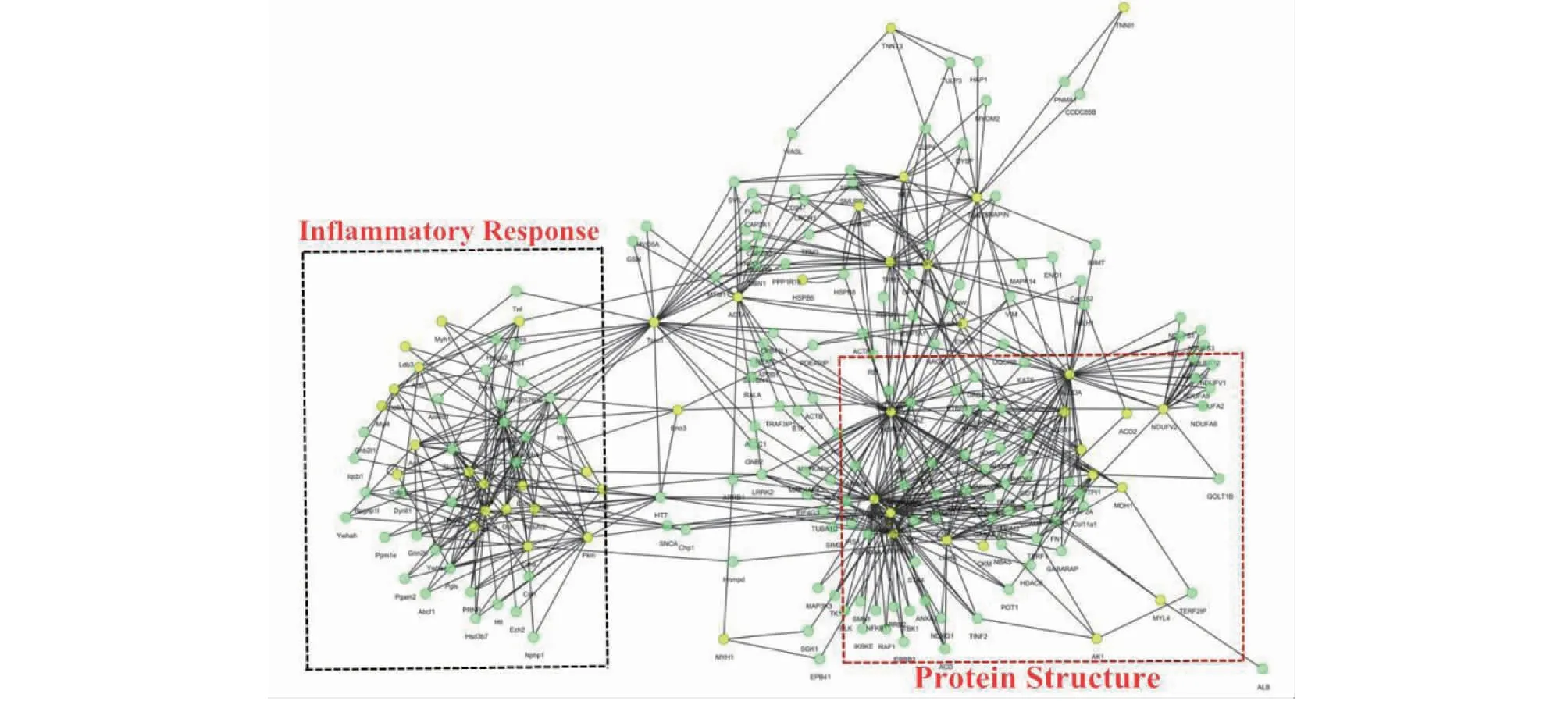
图5 基于Cytoscape软件的不同饲养模式下牦牛空肠提取物差异表达蛋白的蛋白质互作作用网络Fig.5 Protein-protein interaction networks of the differential abundance proteins of different feeding groups in Yak jejunum based on Cytoscape software
3 讨 论
空肠是动物消化吸收的主要场所,在消化系统中扮演者极其重要的角色,对牦牛来说极其的重要。近年来由于青藏高原冬季牧草不足的原因,更多的人选择用舍饲和半舍饲的饲养方式来饲喂牦牛,以提高牦牛的经济效益为自己减少损失[16]。同时舍饲饲养也可以提高牦牛的生产性能,使牦牛的日增重增加,但长期的高精料饲养模式很容易对动物的健康造成影响,也容易对动物肠道造成伤害[17]。空肠的机械屏障通过维持空肠粘膜的正常形态结构对空肠进行机械保护,一旦被破坏就会引起空肠功能障碍,导致肠上皮细胞被感染,致使紧密连接结构损伤,通透性增加和炎症反应[18],会促使肠道损伤,增加肠原性感染的发生[19]。
根据以上研究可以得出,靶蛋白可以分为两个主题,分别为蛋白质结构通路(紧密连接信号通路,钙信号通路和调控干细胞的多能性的信号通路)和炎症反应通路(MAPK信号通路,趋化因子信号通路和细胞因子受体相互作用)。GPR41会在被瘤胃所消化的短链脂肪酸所激活,可以调控脂类的代谢,在肠道对于营养物质的消化吸收中起了重要的作用,作为营养能量调节受体影响了紧密连接的信号传导通路[20-21]。GPR41也参与炎症反应的调节,大剂量摄入可启动肠上皮细胞的糖异生过程,激活紧密连接信号通路,然后调节下游相关细胞增殖和凋亡通路,降低紧密连接蛋白的表达,同时,GPR41介导调节性T细胞增殖并分泌大量的抗炎因子(IL-6,TNF-α,CCL5),促进炎症反应通路(MAPK)相关基因的表达,从而调节炎症因子的表达[22-23]。在本研究中检测到相对于放牧饲养组,在高精料的全舍饲饲养中GPR41明显上调,可得出在高精料全舍饲饲养的牦牛空肠中会促进炎症等一系列反应[24]。
本研究还测定出几种差异蛋白:Occludin,Claudin,Zo-1,ATP1B3,HKDC1,IL-6,TNF-α和CCL5。在与放牧组相比,在高精料的全舍饲饲养条件下Occlusion,Claudin,Zo-1,ATP1B3和HKDC1的表达明显下调,而IL6,TNF-α和CCL5则呈相反的趋势。紧密连接蛋白是空肠机械屏障的重要组成部分,由转运蛋白家族组成,包括Occludin,Claudin和Zo-1,主要参与肠细胞的增殖分化,凋亡和细胞旁路通透性调节,抵御大分子的入侵[25-28]。所以当Zo-1,Occludin和Claudin下调时会影响肠道机械屏障的完整性和通透性,增大了粘膜的通透性,会使得进入血液的病菌增多,促进炎症反应从而对牦牛个体产生影响[29-30]。NF-κB信号通路是调控炎症的主要通路之一[31],发生炎症反应之后NF-κB信号通路会被显著激活诱导IL-6,TNF-α炎症因子的表达,同时L6和TNF-α会促使T细胞产生淋巴因子,从而促进炎症反应的进行[32]。CCL5对于白细胞的趋化作用,多聚的CCL5还会促进细胞的分化和生长,能够激活炎性因子是炎症反应过程中很重要的一个步骤[33]。所以IL6,TNF-α和CCL5的上调同样也是在促进炎症反应,容易对牦牛个体造成损害。
4 结 论
不同的饲养模式下筛选了牦牛空肠差异蛋白质96种,其中9种蛋白质(Occludin,Claudin,Zo-1,ATP1B3,HKDC1,GPR41,IL-6,TNF-α和CCL5)相对较活跃,并且与牦牛空肠屏障和功能损害有关。由此可得,长期高精料的全舍饲饲养模式会对牦牛的空肠造成一定的损害,高精料的全舍饲饲养只适合短期内的牦牛育肥,而不适合长期的牦牛养殖。
猜你喜欢
今日农业(2022年2期)2022-11-16 12:29:47
散文诗(2021年22期)2022-01-12 06:13:54
世界科学技术-中医药现代化(2021年9期)2021-12-31 03:31:02
小哥白尼(野生动物)(2020年9期)2021-01-18 07:23:50
河南畜牧兽医(2020年1期)2020-01-11 05:00:25
河南畜牧兽医(2020年3期)2020-01-10 14:45:01
兽医导刊(2016年6期)2016-05-17 03:50:26
中国当代医药(2015年33期)2015-03-01 02:09:29
河南医学研究(2014年5期)2014-02-27 14:52:44
河南医学研究(2014年3期)2014-02-27 14:51:44
