
H9N2亚型禽流感病毒NS1蛋白原核表达载体的构建及表达
2022-01-28 07:09刘宇卓黄欣梅赵冬敏韩凯凯章丽娇刘青涛
江苏农业科学 2022年1期
马 平, 杨 婧, 刘宇卓, 李 银, 黄欣梅, 赵冬敏, 韩凯凯, 章丽娇, 刘青涛
(1. 江苏省农业科学院兽医研究所,江苏南京 210014; 2.南京农业大学动物医学院,江苏南京 210095)
禽流感病毒属于正黏病毒科中的甲型流感病毒属,它由8个分节段的有包膜的单股负链RNA构成了全基因组,且每个节段至少编码一种蛋白质,AIVs共编码11 种蛋白。其中,非结构蛋白NS(nonstructural)基因是内部基因组中最小的基因节段,它共编码NS1和NS2(NEP)2种蛋白,非结构蛋白NS1存在于除病毒颗粒以外的病毒上清或是被感染的细胞体内。NS1蛋白在AIVs感染早期主要存在于细胞核中,随着感染时间的推移,NS1蛋白分布扩散至细胞浆中,并且能刺激机体产生特异性抗NS1蛋白的抗体。NS1的C-末端效应结构域是与宿主相互作用最频繁的功能结合区,其中,第134~161位氨基酸为其核心序列,该部分序列发挥了阻断宿主mRNA剪接、多聚腺苷酸化和核输出等一系列作用。NS1在病毒-宿主相互关系中发挥了极其重要的作用,该病毒成分不仅在AIVs感染宿主过程中能拮抗机体的天然免疫反应,而且可以影响流感病毒的致病性和宿主的适应性,因此对NS1蛋白的功能、致病性等进行进一步研究至关重要。
NS1是由流感病毒最后一个节段的RNA编码合成,该蛋白含202~237个氨基酸且其大小约为 26 ku,它的结构域包括:N末端1~73个氨基酸构成的RNA结合结构域和C末端88~202个残基的效应结构域,两者之间连接区域为10~15个氨基酸形成的链接区域。NS1蛋白是一种从病毒和宿主两方面进行调节的多功能调控蛋白,从病毒方面来讲,NS1蛋白通过提高流感病毒的翻译能力,提高病毒对宿主的抗病毒反应从而直接性地增强流感病毒的致病作用;从宿主方面来讲,NS1蛋白可通过拮抗干扰素、调控宿主细胞凋亡、抑制宿主细胞蛋白的合成等方式来减弱宿主机体的抗病毒反应从而间接性地增强病毒的致病作用。因此,对蛋白功能的拮抗能够降低甲型流感病毒的复制效率和限制病毒传播空间,具有良好的应用价值。
本研究于2019—2021年在江苏省农业科学院兽医研究所对的基因特性进行了分析,成功构建重组质粒pET32a-,并在BL21(DE3)感受态细胞中进行优化表达,对表达的重组蛋白进行分析和纯化后通过Western-blot和免疫荧光来鉴定,为后续疫苗研发及诊断方法的探究奠定试验基础。
1 材料与方法
1.1 病毒、载体和菌株
H9N2亚型禽流感病毒和pET32a原核表达载体由江苏省农业科学院兽医研究所禽病实验室保存;DH5α和BL21感受态细胞购自南京擎科生物科技有限公司;pMD18-T载体购自TaKaRa公司。
1.2 主要试剂
QuickCutⅠ和QuickCutRⅠ限制性核酸内切酶,购自TaKaRa公司;5 000 bp DNA Marker,购自诺唯赞生物科技有限公司;核酸提取试剂盒、琼脂糖凝胶回收试剂盒和质粒DNA小提试剂盒,均购自美国Axygen公司;镍离子亲和层析蛋白纯化试剂盒和His-tag抗体,均购自碧云天生物公司;辣根过氧化氢酶(HRP)标记的羊抗鼠IgG,购自中衫金桥生物技术有限公司。
1.3 引物设计与合成
根据GenBank库里的全基因序列,针对编码区设计1对特异性引物,引物两端分别引入Ⅰ(碱基序列为CTCGAG)、RⅠ(碱基序列为GAATTC)酶切位点,此引物送至金斯瑞生物科技有限公司合成。引物序列:F:5′-C A A A G A A T T C A T G G A T T C C A A C A C T-3′(含RⅠ酶切位点),R:5′-T T T C C T C G A G C T A C T T T G G A G A A A-3′(含Ⅰ 酶切位点)。
1.4 目的基因的扩增与克隆
利用RNA提取试剂盒提取H9N2 AIVs的基因组,以提取的基因组为模板,用引物R进行反转录,所得产物为cDNA,作为PCR扩增的模板。PCR扩增体系为50 μL,2×Mix 25 μL,上下游引物(10 μmol/L)各1 μL,cDNA 3 μL,超纯水20 μL。扩增程序为:94 ℃ 5 min;94 ℃ 30 s,52 ℃ 45 s,72 ℃ 3 min,10个循环;94 ℃ 30 s,58 ℃ 45 s,72 ℃ 1 min,30个循环;最后72 ℃延伸7 min。将扩增产物进行琼脂糖凝胶电泳,用胶回收试剂盒回收纯化阳性片段,与pMD18-T载体连接后,转化至大肠杆菌DH5α,挑取阳性克隆进行序列测定,筛选出含有保真序列的基因的克隆。
1.5 重组质粒的构建与鉴定
将筛选到的保真质粒pMD18-T-NS1与表达载体PET32a进行双酶切,分别电泳鉴定并回收PET32a酶切质粒和基因片段,将两者在16 ℃连接30 min后转化至大肠杆菌DH5α,挑取单克隆菌落扩大培养后提取质粒,对质粒进行PCR和双酶切鉴定。
1.6 NS1蛋白的诱导条件优化
为摸索蛋白诱导表达的最佳条件,将构建成功的PET32a质粒转化到表达感受态细胞BL21中,鉴定为阳性的菌株进行扩大培养,37 ℃振荡培养至0.6~1.0,分别以不同浓度的TPTG和不同诱导时间来进行优化,根据SDS-PAGE的结果,确定最佳优化条件。
1.6.1 IPTG最佳浓度的探索 在5管为0.6~1.0的4 mL菌液中加入不同量的IPTG使其终浓度分别为0.0、0.1、0.5、1.0、2.0 mmol/L,37 ℃,120 r/min 诱导6 h,诱导结束后,8 000 r/s离心 10 min,弃上清,1×PBS洗涤3遍后用480 μL PBS重悬,取样本进行SDS-PAGE分析,根据结果确定IPTG 最佳诱导浓度。
1.6.2 最佳诱导时间的探索 在培养基中加入终浓度为2.0 mmol/L IPTG,分别诱导2、4、6、8 h,对样品进行SDS-PAGE检测,根据电泳结果确定出最佳诱导时间。
1.7 重组蛋白的可溶性分析及纯化
挑取阳性单克隆于4 mL LB培养基中,37 ℃振荡培养过夜,第2天以1 ∶100的比例扩大培养至菌液为0.6~1.0,加入终浓度为2 mmol/L的IPTG,37 ℃、120 r/min诱导表达8 h后8 000 r/s离心 10 min,弃上清,沉淀用PBS反复洗涤3次,重悬于480 μL PBS中,间歇超声波冰浴破菌后,12 000 r/s 离心10 min,分别对上清和沉淀进行 SDS-PAGE 检测。根据可溶性分析的结果,按照碧云天纯化试剂盒的步骤对蛋白进行纯化。
1.8 纯化蛋白的Western blot分析
将凝胶上的蛋白转至PVDF膜,转膜完成后加入含5%脱脂奶粉的PBST在37 ℃培养箱中封闭 2 h,后与1 ∶1 000稀释的His单克隆抗体4 ℃反应过夜,用PBST洗涤3次,每次5 min,再与辣根过氧化物酶标记的二抗在37 ℃温箱中反应1 h,用PBST洗涤3次后显色,进行免疫印记分析。
1.9 纯化蛋白的免疫原性分析
采用纯化的NS1重组蛋白多次免疫小鼠制备血清,以其作为一抗对感染H9N2禽流感病毒8 h 的MDCK细胞进行免疫荧光试验,测定所制备的血清能否识别H9N2禽流感病毒在MDCK细胞复制过程中产生的NS1蛋白。
2 结果与分析
2.1 NS1基因的RT-PCR扩增与保真克隆的筛选
提取H9N2 AIVs的基因组,用NS1引物进行RT-PCR扩增,由图1可知,该试验扩增出与预期大小一致的片段。将该片段进行胶回收后将产物连接到pMD18-T载体上,通过序列测定筛选出保真基因克隆。

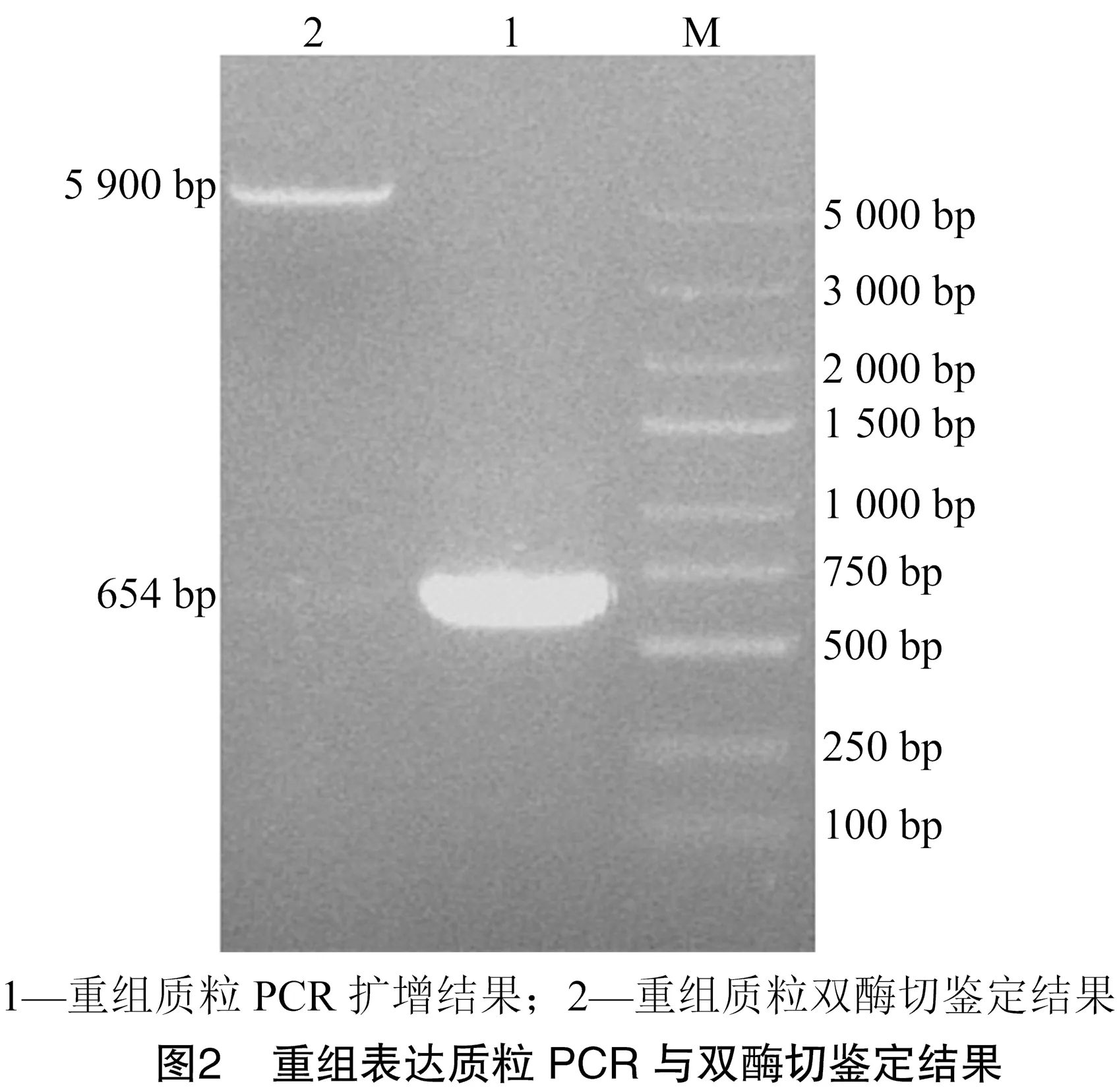
2.2 重组表达质粒的PCR与双酶切鉴定结果
对克隆质粒pMD18-T-进行双酶切,将切下的基因连接到原核表达载体pET32a,然后对重组表达质粒进行PCR和双酶切鉴定。由图2可知,基因成功连接到质粒pET32a,重组表达质粒构建成功,命名为pET32a-。
2.3 NS1蛋白表达条件的优化
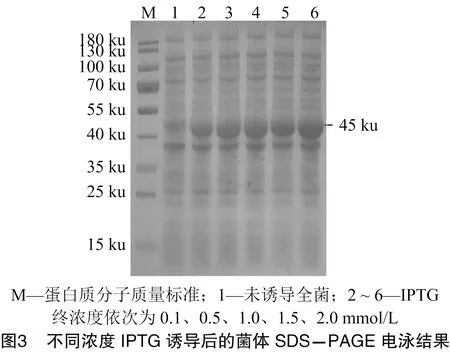
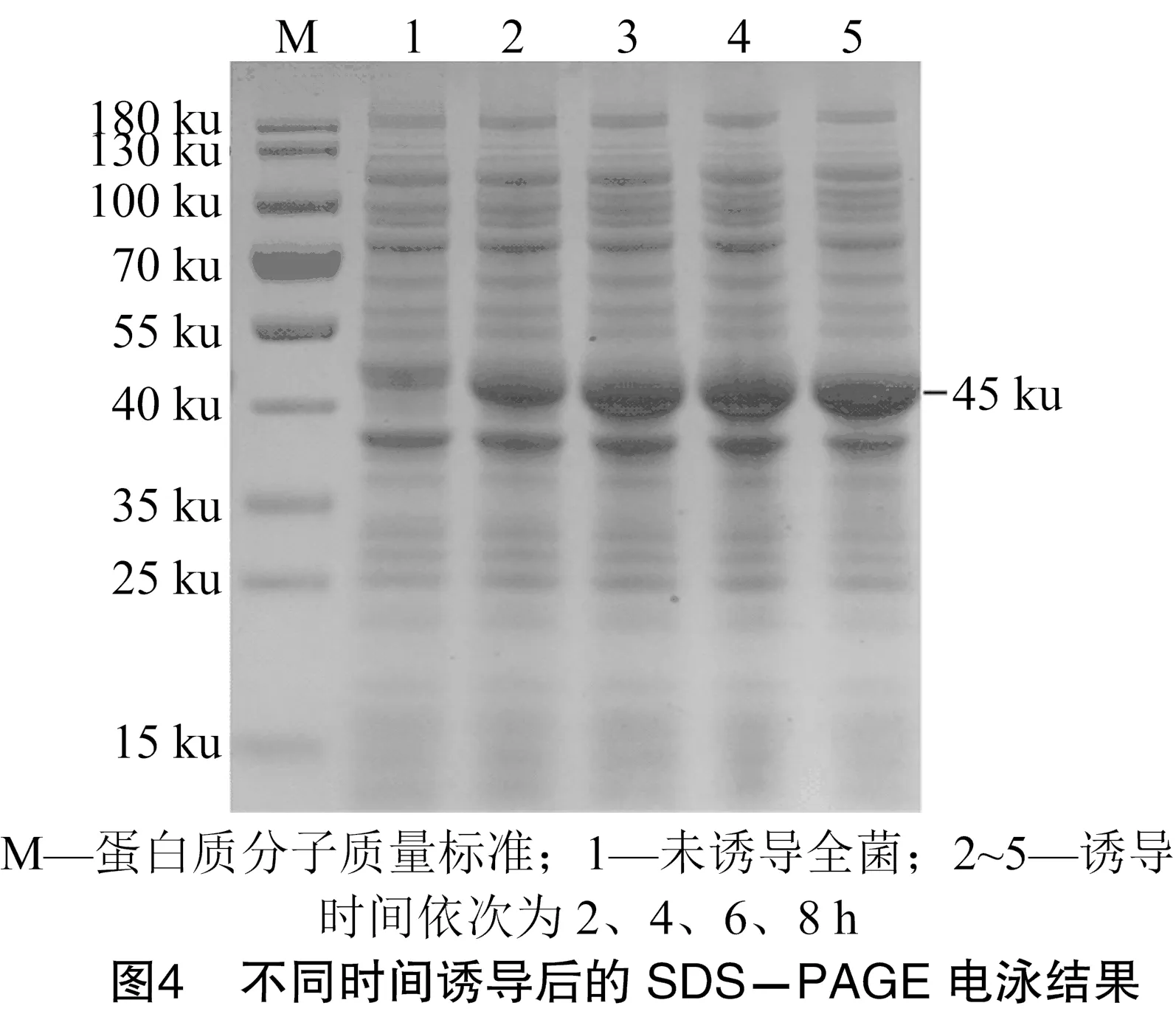
将构建好的重组质粒pET32a-转化至表达感受态BL21(DE3),分别对IPTG的使用浓度和诱导时间进行最佳条件探索。由图3、图4可知,最佳IPTG的使用浓度为2 mmol/L,最佳诱导时间为8 h。
2.4 表达产物的可溶性分析
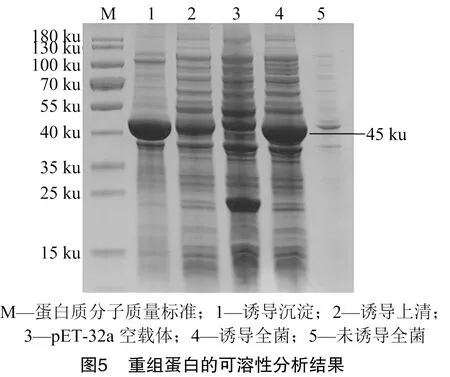
确定重组蛋白的最佳诱导表达条件后,对重组蛋白进行大量表达,并对表达产物的可溶性进行分析。由图5可知,重组蛋白主要存在于细胞裂解沉淀中, 即NS1重组蛋白在细胞中主要以包涵体的形式存在。
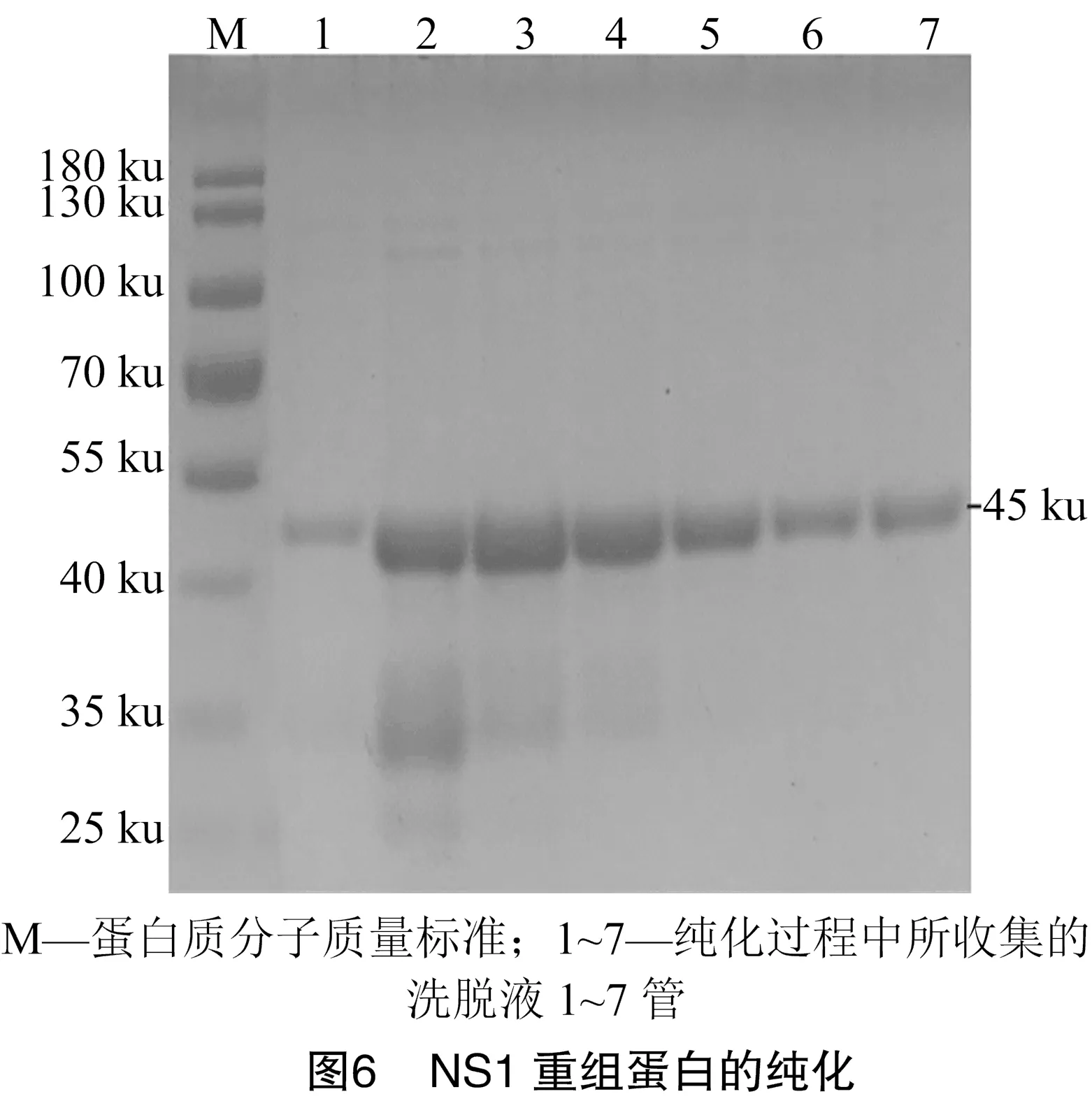
2.5 NS1重组蛋白的纯化与鉴定
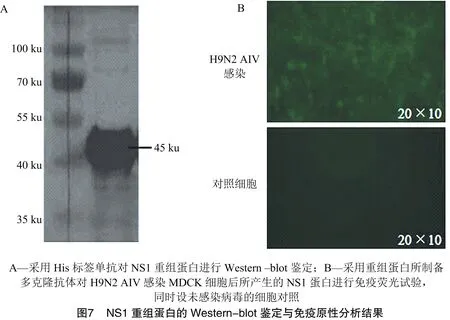
由图6可知,将收集的包涵体用裂解液进行溶解,用镍离子亲和层析柱对溶解上清中的重组蛋白进行纯化,并对纯化后的重组蛋白进行Western-blot鉴定。由图7-A可知,重组蛋白可以被抗His标签的单抗识别,提示NS1重组蛋白表达、纯化成功。
2.6 重组蛋白的免疫原性分析
采用纯化的NS1重组蛋白多次免疫小鼠制备血清,采用该抗体对感染H9N2 AIV的MDCK细胞进行免疫荧光试验。由图7-B可知,所制备的多克隆抗体可以识别H9N2 AIV感染MDCK细胞后所产生的的NS1蛋白,提示本试验所表达、纯化的NS1重组蛋白具有良好免疫原性。
3 讨论
H9N2亚型禽流感病毒是最常见的低致病性禽流感病毒,最早于1966年从北美的火鸡体内分离到;我国于1994年在广东某一鸡场首次分离出H9亚型禽流感病毒;随后,该型禽流感在我国养禽地区呈大流行趋势。流行病学调查显示,H9亚型禽流感病毒的主要宿主是野生水禽,而家养水禽和鸡对H9N2 AIVs也易感,这些宿主在H9N2 AIVs向哺乳动物的跨物种传播中发挥重要作用。自1999年以来,我国不断有报道人感染H9N2亚型禽流感病毒的病例,这使H9N2亚型禽流感受到广泛关注和高度重视。H9N2亚型禽流感病毒不仅给养禽业造成极大损失,也严重威胁到人类健康,所以对H9N2亚型禽流感病毒的研究具有重要的公共卫生学意义。NS1是甲型流感病毒的一种非结构多功能调节蛋白,在流感病毒的致病性等方面发挥着重要的调节作用,对NS1蛋白进行深入研究有助于更好地理解流感病毒与宿主细胞间的相互作用,为揭示禽流感病毒的致病分子机制奠定了理论基础。
本研究利用RT-PCR技术扩增出H9N2 AIVs的基因片段,连接pMD18-T载体进行克隆,然后将克隆的保真基因与载体pET32a连接,构建重组质粒pET32a-,并转化到BL21(DE3)感受态细胞中进行诱导表达,确定最优的诱导表达条件,利用His-tag镍柱纯化重组蛋白,利用SDS-PAGE和Western-blot进行鉴定。结果显示,采用pET32a载体可在感受态BL21(DE3)中进行NS1蛋白的表达,NS1蛋白的表达量与诱导剂IPTG的浓度和诱导时间有一定相关性,当IPTG浓度为2 mmol/L、诱导时间为8 h时,NS1融合蛋白的表达量最大。由于pET32a带有His标签,所以本研究对融合蛋白用镍柱进行了纯化,并对纯化的融合蛋白用针对His标签的单克隆抗体进行了鉴定,结果表明融合蛋白可以与His单抗进行结合。而免疫荧光试验结果显示,利用纯化的重组蛋白制备的多克隆抗体可识别H9N2 AIVs在宿主细胞内产生的NS1蛋白。以上结果说明,本研究成功表达和纯化了H9N2禽流感病毒的NS1蛋白,并且重组NS1蛋白具有良好的免疫原性,这为下一步研究NS1蛋白的功能,以及以NS1蛋白为靶标的禽流感病毒临床感染的诊断和抗病毒药物的研制奠定了基础。
猜你喜欢
环球时报(2022-09-20)2022-09-20
今日农业(2020年24期)2020-12-15
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
启蒙(3-7岁)(2017年4期)2017-06-15
兽医导刊(2016年12期)2016-05-17
实用皮肤病学杂志(2015年4期)2015-12-22
医学研究杂志(2015年12期)2015-06-10
现代检验医学杂志(2015年4期)2015-02-06
当代畜禽养殖业(2014年7期)2014-02-27
