
基于空间自相关模型的西北太平洋日本鲭渔场时空变动研究
2022-01-22 05:01赵国庆吴祖立崔雪森樊伟石永闯肖戈唐峰华
海洋学报 2022年1期
赵国庆,吴祖立,崔雪森,樊伟,石永闯,肖戈,唐峰华*
( 1. 上海海洋大学 海洋科学学院,上海 201306;2. 中国水产科学研究院东海水产研究所,上海 200090)
1 引言
日本鲭(Scomber japonicus)是一种具有重要商业价值的大洋暖水性中上层鱼类,沿大陆架广泛分布于太平洋、大西洋和印度洋及其邻近海域,横跨温带和亚热带[1-2],垂直分布范围为0~300 m[3],具有趋光、垂直移动特性,在产卵期(3-6月)和越冬季(12月至翌年2月)会进行长距离洄游[4]。日本鲭是西北太平洋公海灯光围网的主要捕捞对象,日本在该海域捕捞历史悠久,另外,在该海域进行生产的国家还有中国、韩国和俄罗斯[5-6]。根据季节性洄游模式和产卵场位置的不同,西北太平洋日本鲭被划分为对马暖流群体和太平洋系群体[7],前者产卵地为东海南部到对马海峡之间的海域,后者往往在黑潮洋流边缘产卵[1],我国在西北太平洋公海的主要捕捞对象为太平洋系群体。据研究,20世纪70年代该群体的资源量一直处于较高水平,80年代初开始下降,直至2012年一直处于较低水平,自2014年开始,其资源量大幅提升,目前,太平洋系群体的利用程度较低,捕捞潜力较大[6]。自20世纪70年代以来,太平洋系群体资源量一直处于剧烈变动中,且变动强度要远大于对马暖流群体[6],据推测,这可能与海洋环境的改变和渔业管理制度的实施有很大关系。
资源量变化是渔业生产者、管理者主要的关注内容,但渔场变动也应同样受到重视。渔场变动研究可以较好地展示渔场的变动方向、分布格局、时空演变等特征,对捕捞生产、保护管理等意义重大。灯光敷网和灯光围网是我国在西北太平洋的主要作业方式,日本鲭是这两种作业方式的主要目标种,渔获组成占比高达80%以上[8],因此,日本鲭渔场变动一直是西北太平洋渔业研究的重要内容。目前,相关研究多利用渔业生产数据来计算渔场重心以探讨渔场变动的特征,如采用渔获量重心法、地统计插值法等研究日本鲭渔场重心的季节性变动、渔场区域的变化等[9-10],同时也多结合环境数据探讨渔场的变动是否与环境因子存在相关性,如将海面温度、叶绿素浓度、海流、温跃层等数据与日本鲭渔场变动特征结合起来进行研究[10-11]。Wang等[1]通过对越冬和产卵地指数分析后指出,大尺度气候因子对日本鲭越冬场和产卵场的温度变化具有重要影响,并认为该指数可以较好地解释其资源丰度的变化。但是目前有关渔场变动的研究多缺乏对变动特征的描述,如分布格局、扩张范围、方向、冷热点分析等。渔场变动是时间和空间互相叠加产生的结果,因此,地理上的空间统计方法对渔场变动分析具有很大优势。空间统计是分析特定时间范围内某一空间尺度下某要素是否存在统计意义上的聚集或离散性质的一种有效方法,已发展成为研究自然界具有随机性和规律性变化的普遍性的科学方法,在诸多领域如经济[12]、医学[13]、生态[14]等方面都得到成功应用,目前在渔业中也已得到应用[15]。全局莫兰指数和热点分析是研究某一要素全局和局部分布模式的有效方法,可以得到变量的全局空间分布特征和具有统计显著性的热点和冷点,因此,可以清晰展示渔场变动的相关特征[16]。标准差椭圆模型是一种定量描述研究对象空间分布整体特征及时空演变过程的空间格局统计分析方法,可以直观地展现渔场在空间上的变动方向、扩张海域和分布格局[17]。
基于商业捕捞渔船进行渔业研究对了解渔业动态、量化生物资源量具有很大优势[18],然而,目前基于长时间序列渔业数据对西北太平洋日本鲭渔场变动的研究相对较少。基于此,本研究以我国2014-2019年西北太平洋日本鲭捕捞统计资料为基础,首先采用全局空间自相关工具全局莫兰指数和局部空间自相关工具局部热点分析分析了日本鲭全局和局部空间分布模式,然后采用重心迁移轨迹模型和标准差椭圆模型分析了统计年间日本鲭渔场重心的时空变动轨迹和分布范围的变迁。本研究综合运用了多种时空分析工具,对西北太平洋日本鲭渔场的时空分布特征进行了研究,以期为日本鲭合理开发利用、保护和管理等提供借鉴。
2 材料与方法
2.1 数据来源及渔场区域
本文所用统计数据来源于中国远洋渔业协会公海围拖网渔业技术组所收集的西北太平洋公海灯光围网日本鲭生产统计资料,该资料共由国内14家远洋渔业公司提供,统计内容包括作业日期(年、月、日)、作业位置(经度和纬度)、投网次数、渔获量等信息。本文所用统计数据的时间跨度为2014-2019年,主要作业时间为每年的4-11月,作业区域分布在35°~45°N,145°~165°E(图1)。本文在进行产量分布分析时将经纬度0.5°×0.5°定义为一个小渔区,统计每个小渔区的产量之和来进行产量空间分布研究,同时,为兼顾每个捕捞点对渔场的贡献,在进行空间自相关研究、产量重心轨迹迁移和标准差椭圆分析时使用产量的点要素。
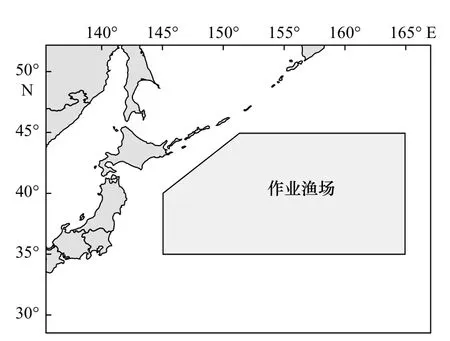
图1 西北太平洋公海日本鲭作业海域Fig. 1 Fishing area of chub mackerel in the high seas of the Northwest Pacific
2.2 渔获数据处理
对渔获量数据进行处理,利用单位捕捞努力量渔获量(Catch Per Unit Effort,CPUE,单位:t/网次)表示渔获效率[17],计算公式为
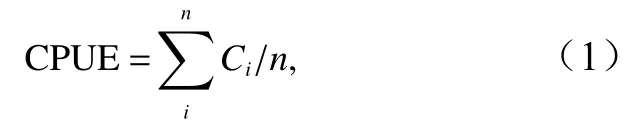
式中,Ci为每艘渔船第i网次的渔获量(单位:t);n是总网次。本文在进行年份和月份的渔获量、CPUE统计和计算时,使用的均是累加数据。
2.3 核密度估计
核密度估计(Kernel Density Estimation,KDE)是一种常用的点密度方法,在概率论中被用于概率密度函数的估算[19],本文采用KDE模型探索了西北太平洋日本鲭捕捞密度的变化情况,计算公式为
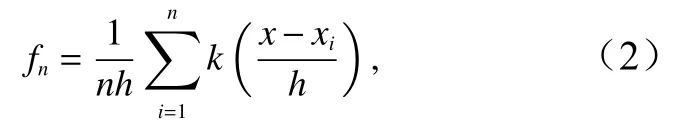
式中,fn为捕捞点的核密度估计值;n为捕捞点的数量;k( )为权重函数,通常取对称的单峰概率密度函数;x-xi为估计捕捞点x到样本捕捞点xi处的距离;h为搜索半径。
2.4 空间自相关检验模型
2.4.1 全局空间自相关
全局空间自相关是对地理属性要素值在整个区域的空间特征描述,用来度量整个研究区域上空间对象之间的关联程度,以表明空间对象之间是否存在显著的空间分布模式,通常采用全局莫兰指数、全局Gear’sC和全局Getis-OrdG等统计量来进行分析[20]。本文采用全局全局莫兰指数来度量日本鲭产量在空间上的全局自相关程度,并利用标准化Z值检验全局莫兰指数的显著水平,计算公式为[21]
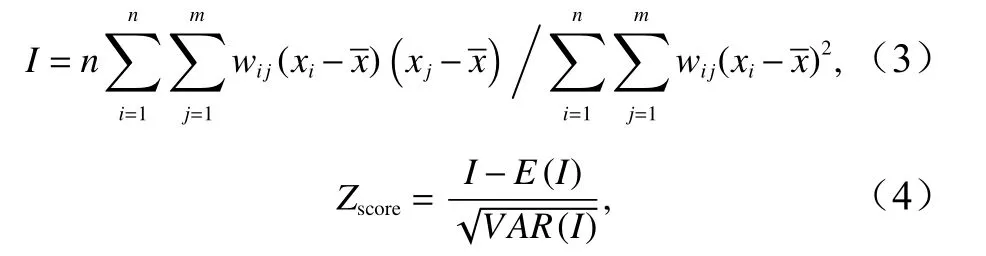
式中,I为全局莫兰指数;n为研究单元总个数;x为产量的平均值;xi、xj分别为第i、j个研究区域日本鲭的产量;wij为i、j区域的空间权重系数,它反映第i、j区域在空间上的关系,并定义为:若区域相邻,wij=1,否则,wij=0;x为产量均值;E(I)、VAR(I)分别表示莫兰指数的期望值和方差。全局莫兰指数取值范围为[-1,1],定义为当全局莫兰指数大于0(p<0.05),表示存在空间正相关,即产量高值(或低值)在空间上显著聚集;等于0或接近0,表示相邻区域不存在空间自相关性,产量随机分布;小于0(p<0.05),表示空间负相关,即相邻区域的产量不具相关性。在显著性为0.05水平下,|Zscore|>1.96,则表明全局莫兰指数显著[21]。
2.4.2 局部空间自相关
全局空间自相关描述了整个研究区域日本鲭分布的自相关程度,但不能有效表达研究区域内不同空间单元与邻近区域在空间上的自相关水平。本文利用局部热点分析(Getis-OrdGi*方法)来展现日本鲭产量在局部区域的空间集聚特征,进而得到具有统计显著性的热点和冷点,以计算日本鲭渔场产量的高值(或低值)在空间上聚类的位置,局部热点分析公式为[22]
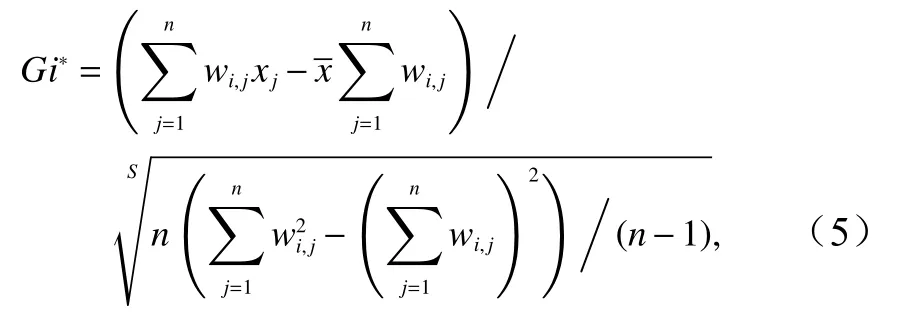
式中,Gi*为局部热点分析值;xj为要素j的属性值;wi,j表示要素i和j之间的空间权重(相邻为1,不相邻为0);S为研究时间段内产量的标准差;n为样本点总数。Gi*统计结果为Z得分,Z得分为正且显著,则表示热点,值越大热点聚集越紧密;Z得分为负且显著,则表示冷点,值越小冷点聚集越紧密[23]。根据Gi*的高低及显著性水平,结合ArcGIS中自然间断点分级法将渔获量区域分为5类:冷点区、次级冷点区、随机分布区、次级热点区和热点区。
2.5 重心迁移轨迹与标准差椭圆分析
重心迁移轨迹模型可基于地理要素的加权中心表达时空迁移规律[24],本文将日本鲭产量作为变量,用该模型来进行渔场重心时空变化计算。标准差椭圆(Standard Deviational Ellipse, SDE)是一种定量描述研究对象空间分布整体特征及时空演变过程的空间格局统计分析方法,其结果能够准确地描述中心性、分布、研究对象的空间分布方向和形状(图2),并能够精确表达地理要素空间分布的整体特征[25],本文采用产量作为权重字段来揭示日本鲭渔场的变动方向和分布格局。相关参数计算公式为
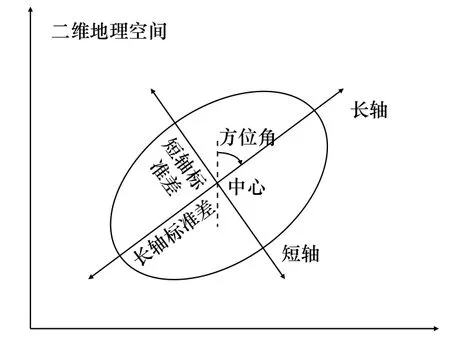
图2 标准差椭圆基本参数Fig. 2 Basic parameters of the standard deviational ellipse
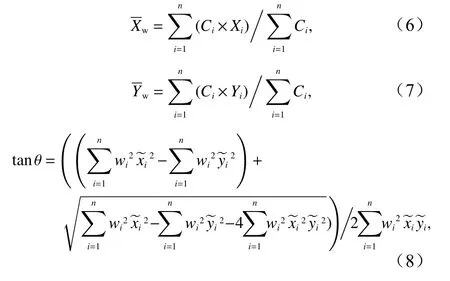
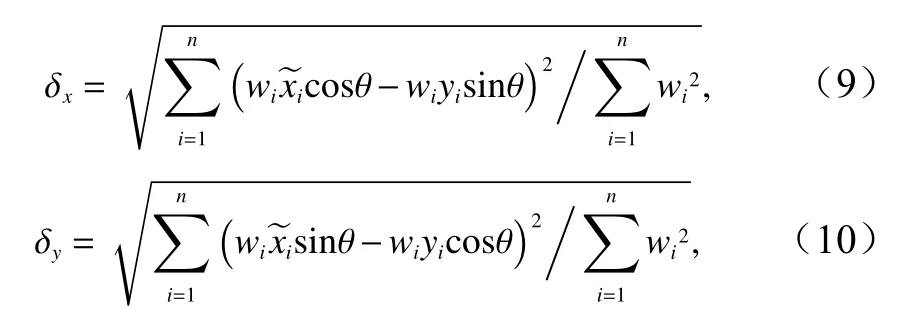
3 结果与分析
3.1 产量和CPUE
2014-2019年,从年间变化来看,日本鲭产量在前5年逐年增长,2018年达到最大值,2019年骤降,恢复到2014年水平;CPUE逐年递减,在8.77~22.92 t/网之间。从月间变化来看,日本鲭产量最低值发生在4月份,6-10月产量相差不大且均维持在较高水平,11月产量下降至与5月产量相当;总体上来看,CPUE呈波动增长趋势,10月份达到最大值,CPUE值的范围为11.96~20.15 t/网(图3)。统计年间,日本鲭产量分布在35°~45°N,145°~164°E范围内,产量高值主要集中的海域范围为39°~44°N,147°~155°E,沿陆地呈带状分布,自155°E往东,产量低值占比高(图4)。以自然间断点分级法对渔获网次的空间分布做核密度可视化分析,结果如图5所示,核密度高值发生在39°~43°N,147°~155°E范围内,且沿陆地呈带状分布,与产量高值的分布范围具有高度一致性。
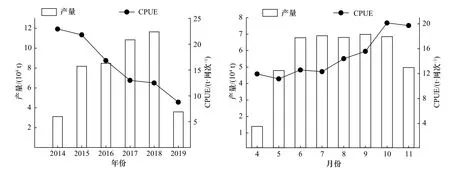
图3 西北太平洋公海日本鲭产量和CPUEFig. 3 Catch and CPUE of chub mackerel in the high seas of the Northwest Pacific
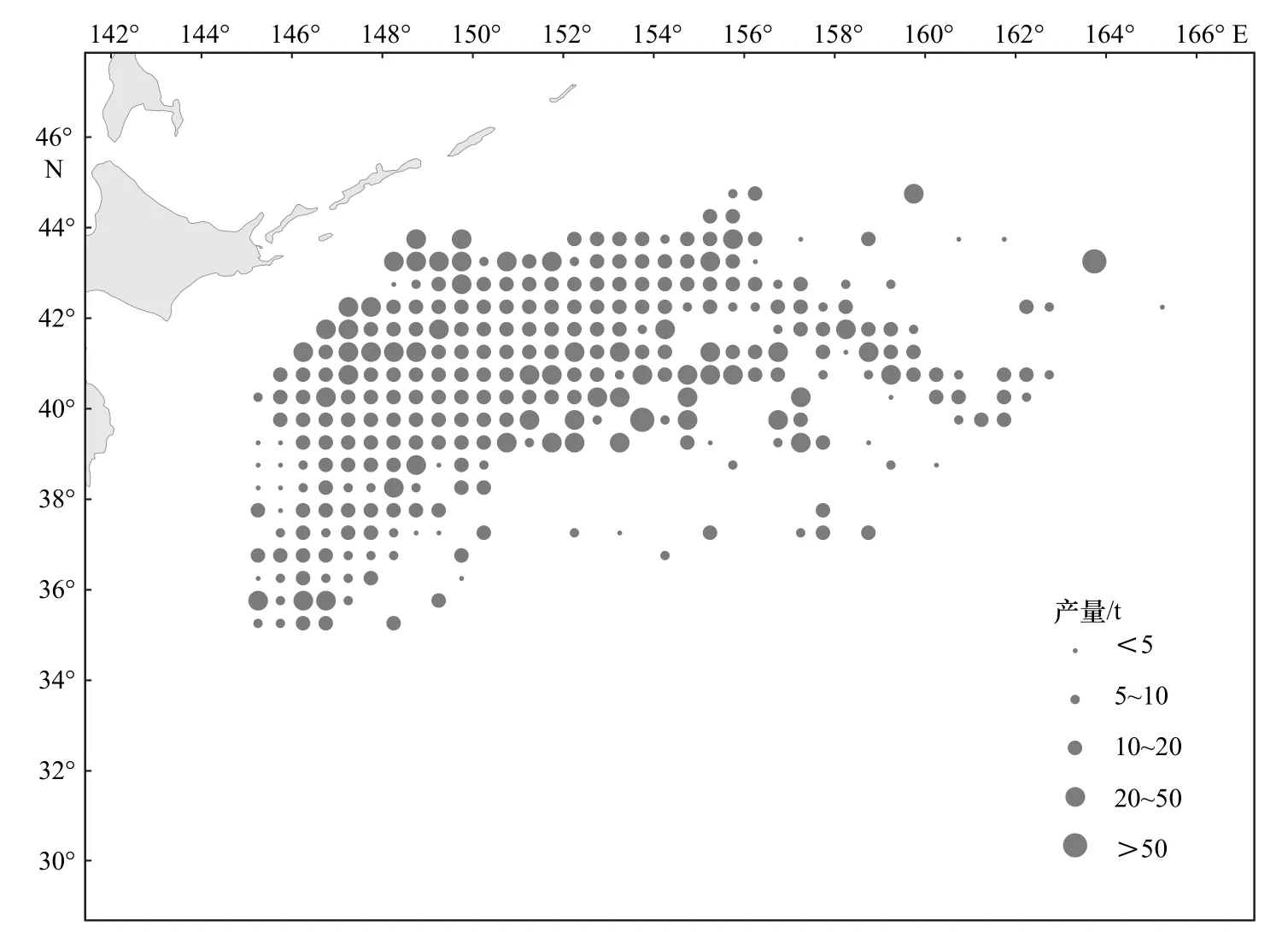
图4 2014-2019年西北太平洋公海日本鲭产量分布Fig. 4 Catch distribution of chub mackerel in the high seas of the Northwest Pacific from 2014 to 2019
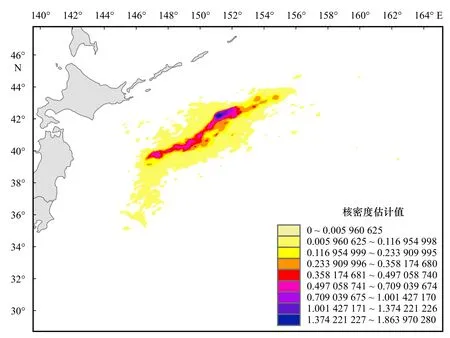
图5 2014-2019年西北太平洋公海日本鲭渔获网次核密度Fig. 5 The kernel density of fishing haul numbers of chub mackerel in the high seas of the Northwest Pacific from 2014 to 2019
3.2 日本鲭渔场分布空间关联性分析
3.2.1 日本鲭渔场的空间聚集性
由表1可知,所有年份偏态大于1,表明年间产量频数分布呈正偏且渔场为高度偏态分布;峰态值大于3,峰陡峭,表明日本鲭年间产量较为集中,渔场具有很强的聚集性;各年变异系数均在0.1~1之间,表明各区域产量差异水平为中等;标准差平方与均值的比值大于1,表明年间各区域日本鲭产量呈强烈聚集分布;各年份全局莫兰指数均为正值,Z得分均大于1.96且p<0.05,因此年间日本鲭渔获量存在空间自相关并呈现显著聚集分布模式,但是2015年、2016年和2018年的全局莫兰指数均小于0.1,因此这3年的空间自相关性较弱。
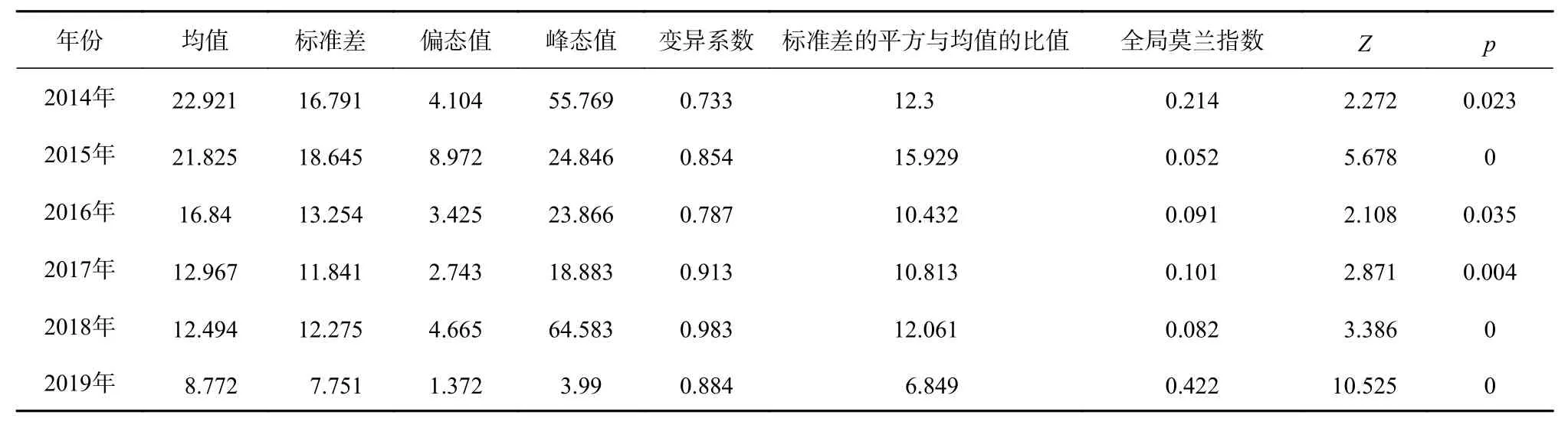
表1 日本鲭产量年间常规统计和全局空间自相关参数Table 1 Ordinary statistics and global spatial autocorrelation parameters of annual catches of chub mackerel
由表2可知,所有月份偏态大于1,表明月间产量频数分布呈正偏且渔场为高度偏态分布;峰态值大于3,峰陡峭,表明日本鲭月间产量较为集中,渔场具有很强的聚集性;4月和5月的变异系数大于1,表明4月、5月各区域产量差异较大,6-11月的变异系数均在0.1~1之间,表明6-11月各区域产量差异水平为中等;标准差平方与均值的比值大于1,表明月间各区域日本鲭产量呈强烈聚集分布;各月份全局莫兰指数均为正值且较大,Z得分均大于1.96且p<0.05,因此月间日本鲭产量存在空间自相关并呈现显著聚集分布模式,且空间自相关性较强。
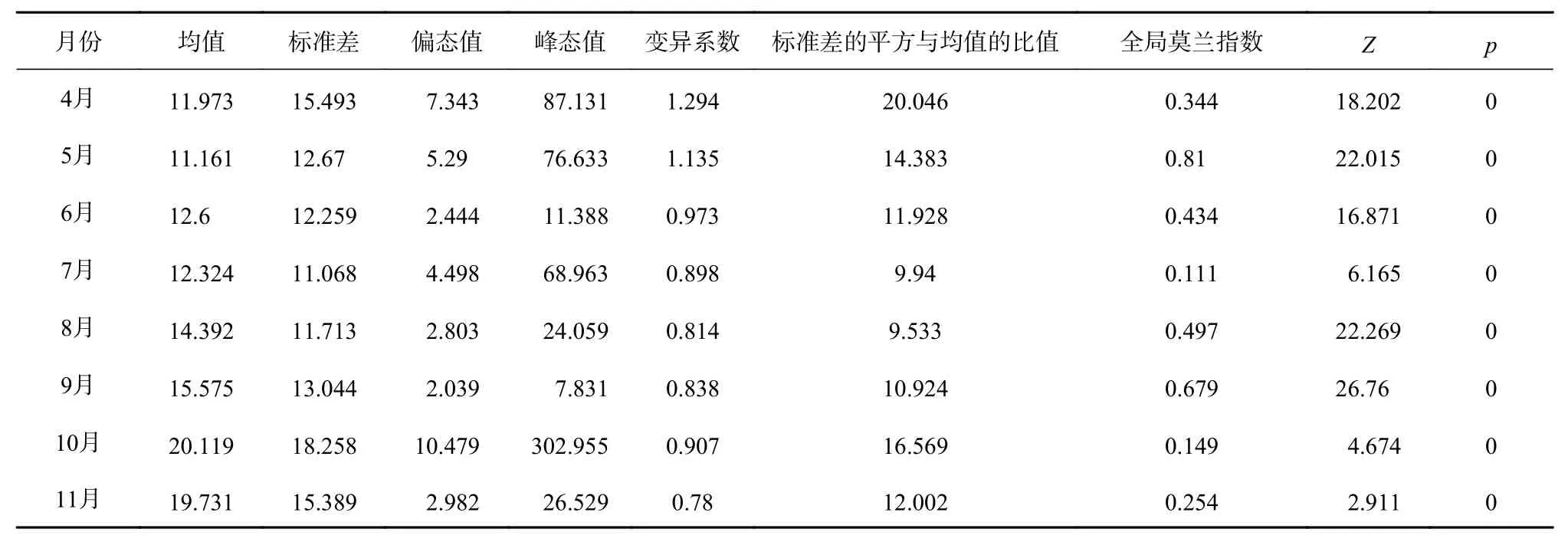
表2 日本鲭产量月间常规统计和全局空间自相关参数Table 2 Ordinary statistics and global spatial autocorrelation parameters of monthly catches of chub mackerel
3.2.2 日本鲭渔场局域空间分布格局
分析2014-2019年的年间和月间西北太平洋产量空间格局热点演化情况,结果如图6和图7所示。年间、月间日本鲭产量分布的热点区和冷点区均表现出一定空间集聚特征,但是不同年份、月份的热点区和冷点区的分布区域、面积均存在较大差异。具体来看,年间热点分析显示,2014年热点区和冷点区的分布海域面积相差不大,且热点区集中在东北方,冷点区集中在西南方;2015年热点区集中在39°~40°N,148°~151°E,冷点区集中在36°~40°N,145°~148°E;2016年热点区主要集中在38°~41°N,146°~151°E,冷点区位于热点区的东北方,集中在40°~43°N,150°~154°E;2017年热点区和冷点区具有间隔分布的特点,从西南往东北逐渐表现为冷点区、热点区、冷点区和热点区;2018年几乎全由热点区和冷点区组成,随机分布区海域面积占比较小,冷点区集中在38°~41°N,146°~152°E,热点集中在41°~43°N,148°~153°E;2019年热点区和冷点区海域面积占比均较小且集中在西南方,大部分海域为随机分布区。
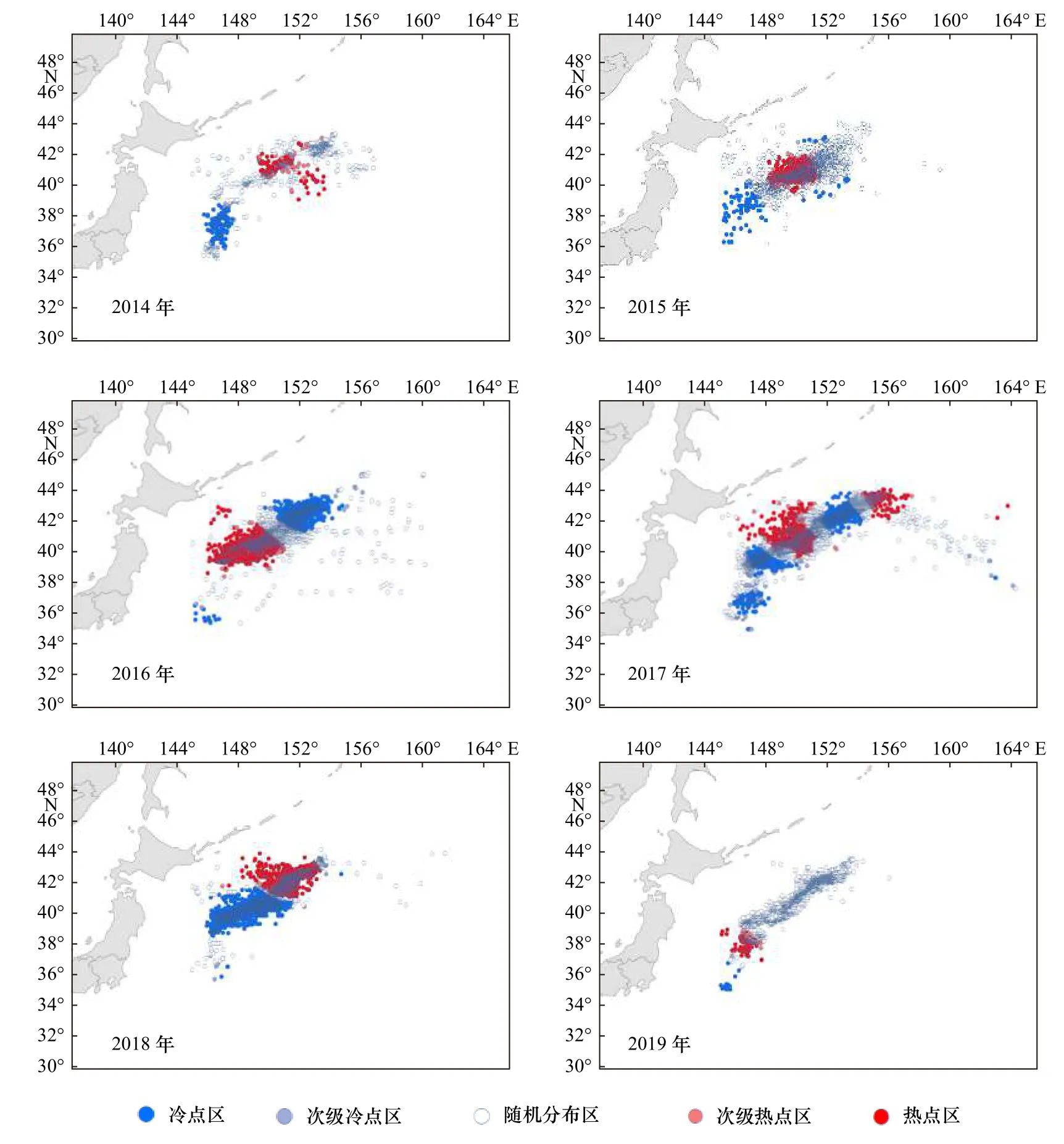
图6 西北太平洋日本鲭产量年间热点分析Fig. 6 Hotspots of annual catches of chub mackerel in the Northwest Pacific

图7 西北太平洋日本鲭产量月间热点分析Fig. 7 Hotspots of monthly catches of chub mackerel in the Northwest Pacific
月间热点分析显示,4月随机分布区远大于热点区和冷点区海域面积之和,热点区和冷点区主要分布在捕捞海域的东北方;5月冷点区位于热点区之间,集中在37°~41°N,145°~149°E,热点区由两部分组成,分别为35°~37°N,145°~148°E和39°~42°N,149°~151°E;6月和7月热点区和冷点区在分布格局上基本一致,热点区占比较大,分别集中在39°~42°N,147°~152°E和40°~43°N,149°~154°E,冷点区都由两部分组成,分布在热点区的东北和西南;8月和9月热点区和冷点区的分布表现出一定的相似性,冷点区均分布于捕捞海域的东北,热点区均分布于西南,8月冷点区占比较大,9月反之;10月冷点区位于热点区之间,集中在41°~43°N,149°~153°E,热点区主要集中在冷点区的西南,集中在39°~41°N,146°~150°E;11月冷点区较为分散,热点区主要集中在39°~41°N,148°~151°E。
3.3 日本鲭渔场变动分析
3.3.1 日本鲭产量重心时空变化
通过重心迁移轨迹模型得到日本鲭产量重心的变化,结果如图8所示。从年间变化来看,产量重心集中在40.8°~41.6°N,150.1°~151.2°E范围内,移动轨迹呈右转约90°的扁“W”形状,总体上往西北移动(图8a)。从月间变化来看,产量重心集中在39.4°~42.1°N,147.7°~151.9°E范围内,从4月份开始逐渐向东北方向移动,8月到达最东北端后,9月开始向西南方向折返(图8b)。
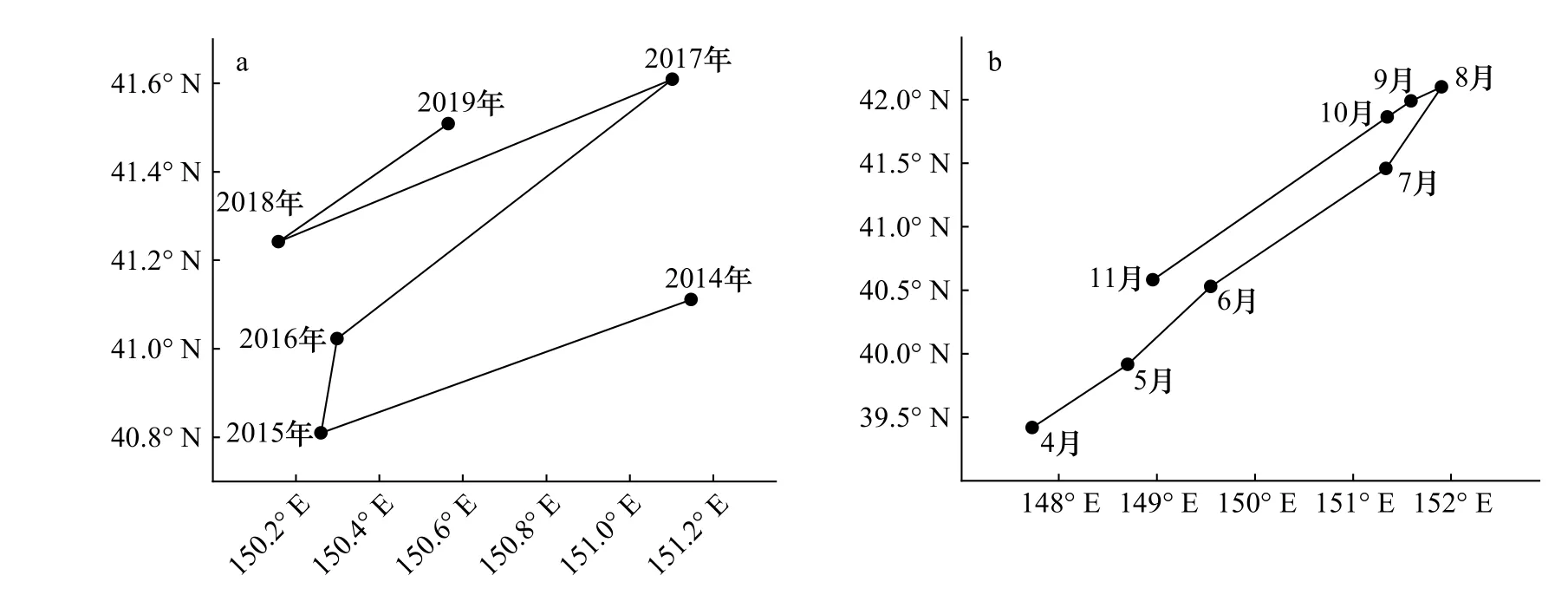
图8 西北太平洋日本鲭渔场重心年间(a)和月间(b)变化Fig. 8 Interannual variation (a) and monthly variation (b) of fishing ground gravity centers of chub mackerel in the Northwest Pacific
3.3.2 日本鲭渔场变动方向和分布格局
日本鲭渔场的时空分异特征如图9所示。从年间变化来看,渔场年间分布方位角变化较小,均在52.84°~67.62°之间变动,表明日本鲭渔场年间分布方向性较强,且具有很强的一致性,均呈现西南-东北格局;年间扩张海域面积变化较小,最小值、最大值分别发生在2018年、2014年;标准差椭圆扁率在3.04~5.88之间,均保持在较高水平,表明日本鲭渔场具有很强的向心力(图9a,表3)。从月间变化来看,渔场月间分布方位角变化较小,分布在47.01°~67.64°之间,表明日本鲭渔场月间分布具有较强的方向性,呈现西南-东北格局;月间扩张海域面积变化较小,最小值、最大值分别发生在4月、9月;标准差椭圆扁率在2.13~4.06之间,向心力较强(图9b,表3)。总的来看,年间、月间日本鲭渔场变动方向一致,均呈西南-东北格局,具有较强的方向性和向心力,表明日本鲭渔场具有较高的聚集性。
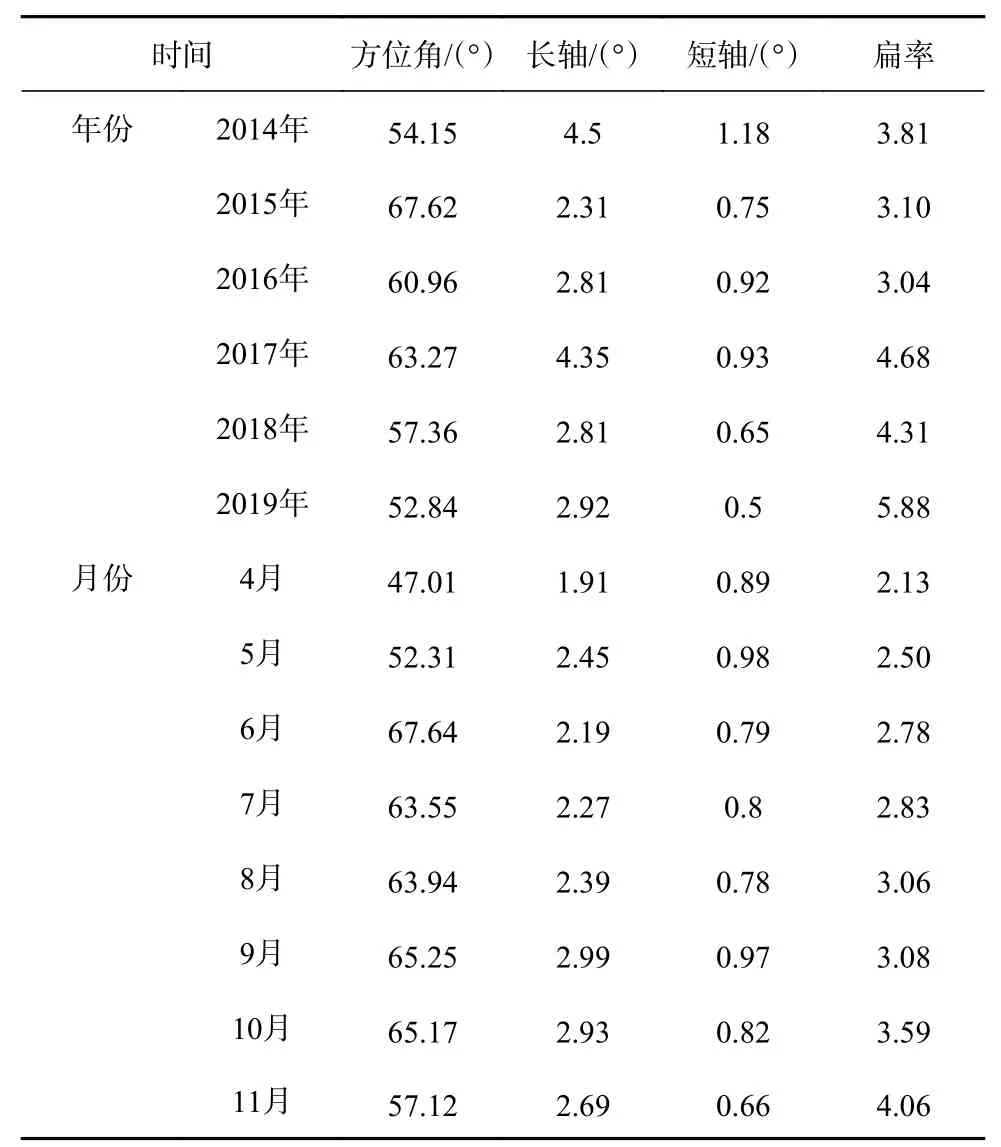
表3 日本鲭年间、月间产量分布标准差椭圆形状参数Table 3 Parameters of standard deviational ellipse of annual and monthly catch distributions of chub mackerel
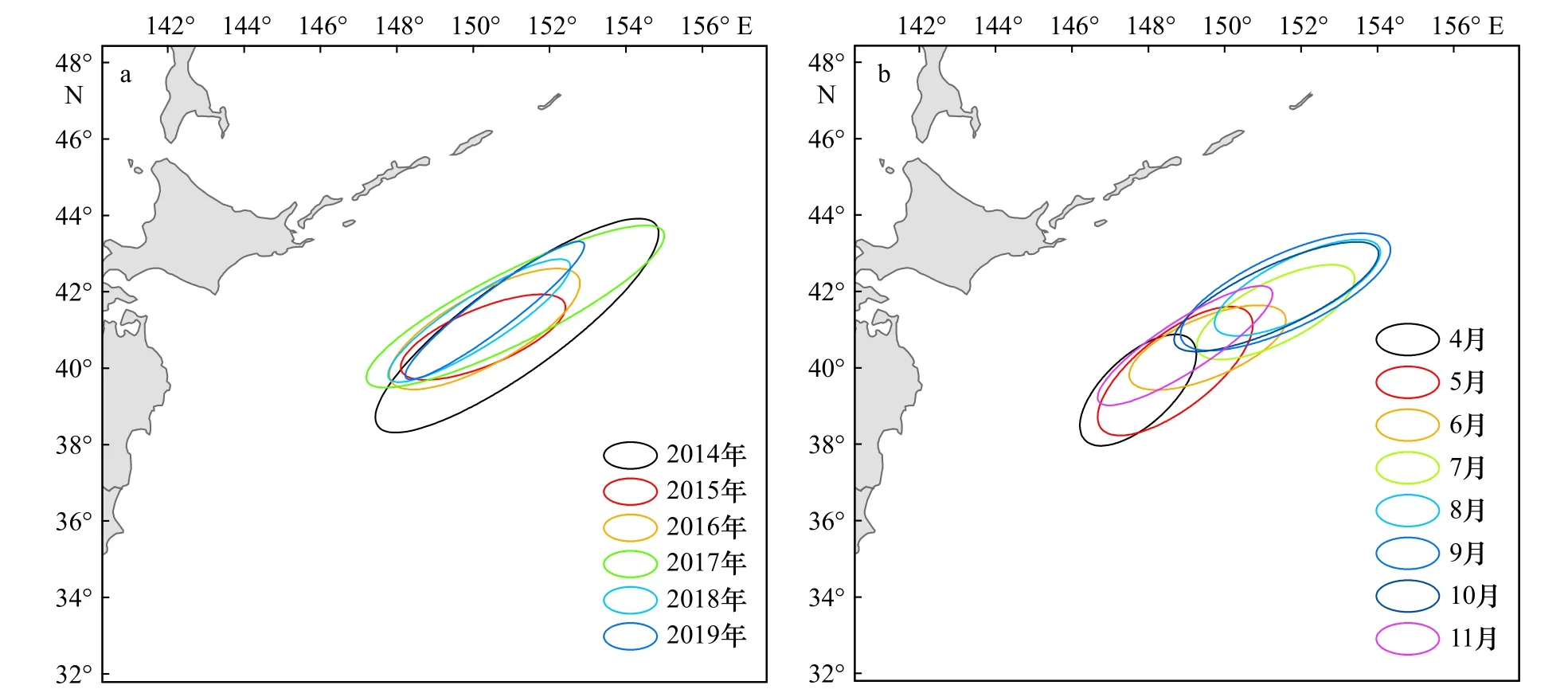
图9 西北太平洋日本鲭渔场年间(a)和月间(b)标准差椭圆Fig. 9 Annual (a) and monthly (b) standard deviation ellipses of chub mackerel fishing ground in the Northwest Pacific
4 讨论
4.1 产量和CPUE时空变化
渔业资源的丰度、产量是渔业生产者和管理者所关注的主要内容,并影响着渔业资源的开发利用。日本鲭是西北太平洋灯光围网的主要捕捞鱼种,其资源丰度和产量均具有明显的季节性变化[8-10]。本文对2014-2019年日本鲭进行统计分析后得出了年间产量先增后降、CPUE逐年递减的结论,日本中央水产研究所年度报告[6]所得结论的产量变动趋势与本文具有一定相似性,但CPUE的差异性较大,究其原因,本文与日方统计的捕捞海域[6,26-27]、作业方式和数据来源[6]等都存在较大差异,可能产生了一定影响。月产量在6-10月均维持在较高水平,且CPUE逐月增加,往年研究也具有此特性[9,28],进一步表明日本鲭的季节性变化规律明显。唐峰华等[29]研究发现,1-5月为日本鲭的成长期,其在成长过程中不断寻找适宜生存的渔场,中心渔场的不确定性较高,导致渔船也跟随进行渔场的找寻,实际作业时间较短,因此,初始捕捞月份的产量和CPUE往往较低。庄之栋等[30]指出,日本鲭在8-10月处于索饵洄游期,分布相对集中,且处于生物量较大值,因此产量和CPUE均较大。11月较大的风浪对捕捞产生了一定影响,导致产量和CPUE均有所下降[9]。
西北太平洋日本鲭产量也具有很明显的地理分布特点。产量高值集中分布于39°~44°N,147°~155°E范围内,沿陆地呈带状分布。薛嘉伦等[31]基于最大熵模型对西北太平洋日本鲭潜在栖息地分布研究后发现,渔场区域与其栖息地分布区域变化基本一致,主要集中在40°~42°N,147.5°~152.5°E海域,同时指出海表面温度是影响西北太平洋栖息地分布的首要因子,海表面温度极大值和极小值的交会对中心渔场的分布有一定影响。而捕捞海域的人为选择性、捕捞渔船的增减、管理政策的实施均是造成产量分布不均的重要因素[32],本文发现,日本鲭产量高值与捕捞网次高值的分布范围具有高度一致性,一方面可以说明渔船可能对于渔场具有较好的掌控能力,但是从另一方面考虑,其产量也可能高度依赖于大量捕捞努力量的投入。
4.2 渔场变动的空间自相关分析
在渔场研究中,利用的往往都是点数据,而掌握整个渔场的资源分布情况对渔业资源、渔业捕捞、渔业管理等研究更具意义,因此,渔业资源平面分布的可视化就显得尤为重要[33]。地理学上的空间自相关模型能够度量某位置的点数据与其他位置点数据之间潜在的相互依赖性[34],能够较好地对空间数据进行统计分析并得出要素之间的关系[35]。本文利用2014-2019年西北太平洋日本鲭渔获数据,通过空间自相关分析模型对该海域日本鲭渔业资源的空间分布特征进行了分析,得到了日本鲭的空间聚集程度和局部空间分布的冷热点特征。研究发现,日本鲭年际、月间产量呈现显著的聚集分布,但个别年份的空间自相关性较弱,不同年份、月份渔场的热点区和冷点区的分布区域、面积均存在较大差异,说明日本鲭渔场具有很明显的空间自相关性。目前,空间自相关已开始被应用于渔业资源分布特征的研究中,其结果对于资源分布、渔场变动等的深入研究具有重要意义[36-38]。
本文所研究的区域位于远离大陆的西北太平洋公海,人类活动影响相对较小,所以日本鲭资源的空间聚集程度可能与种内竞争、环境因素存在一定关系。2015年、2016年和2018年的全局莫兰指数均小于0.1,空间自相关性较其他年份较弱,总体上空间分布格局趋于分散,但是统计结果表明,这3年的产量、CPUE均处于较高水平,据推测,该现象可能会在一定程度上降低鱼类资源因密度过大而加剧的种内资源竞争[36-37],这一现象也可描述为-通过降低地理分布上的空间自相关性来缓解密度过大而导致的种内竞争。而相关研究也发现,其他种类诸如南海的大眼鲷(Priacanthusspp.)[36]、海州湾的日本枪乌贼(Loligo japonica)和短蛸(Octopus ochellatus)[37]等也在季节性分布上存在该特征,并指出该特征可能在渔业资源的分布上具有普遍性。将2014-2019年内同一月份的数据分别累加进行分析可得,各月份渔场的自相关性和聚集性均较强,证明了西北太平洋日本鲭渔场具有明显的季节性变化,究其原因,因为不同年份内同一月份的环境条件相对较为相似,也从侧面验证了薛嘉伦等[31]关于海表面温度与日本鲭栖息地显著相关的结论。
冷热点分析发现,盛渔期6月和7月、8月和9月的热点、冷点的分布分别表现出一定的相似性:6月和7月热点区集中于渔场中部,冷点区集中于渔场两端,热点区占比远高于冷点区;而8月和9月的冷点区分布于渔场的东北部,热点区分布于西南部,8月冷点区占比较大。各年份的冷热点的分布区域、分布范围、所占比例等均表现出较大的差异性。另外,不管是冷点区、热点区还是随机分布区,均各自表现出较为明显的空间聚集性,呈团块状分布,符合水生生态系统研究中普遍存在的球形半方差函数的特征,即当样点间隔距离达到变程之前,样点的空间依赖性随样点的距离增大而逐渐降低[39],这也是海洋渔业资源分布普遍存在的现象,东海带鱼(Trichiurus japonicus)[40]、西北太平洋柔鱼(Ommastrephes bartramii)[41]、中西太平洋鲣鱼(Katsuwonus pelamis)[38]、南海北部大眼鲷[36]等冷热点的分布均表现出团块状聚集状态。渔业资源在地理上冷热点格局的变动在很大程度上反映了渔场中心的变动特征,而引起这种变化的原因是多方面的,如船队动态变化[15]、海洋环境因素(海流[40]、海表温度[33,40]、叶绿素浓度[33])、气候变暖[33]等。冷点区和热点区均具有较强的聚集性,渔船在冷点区和热点区都具有较高的作业频率,本质区别在于热点区作业频次和产量都较高,而冷点区的作业频次高而产量低,但是两者均为中心作业渔场。对于随机分布区,冯永玖等[33]提出两种猜测,一种是作业频次较低,而单船单日的产值可能高也可能低,一种是作业频次较高但空间自相关性较低。渔场冷热点的分布具体受何种因素的影响仍有待继续深入研究。
4.3 渔场时空变异特征分析
2014-2019年,西北太平洋日本鲭渔场的变动范围为35°~44°N,145°~164°E。本文利用重心迁移轨迹模型得到了西北太平洋日本鲭的年间、月间产量重心变动情况,年间产量重心总体上往西北方向移动,月间产量重心显示,从4月开始逐渐向东北方向移动,8月到达最北端,然后向西南方向折返,表现出明显的季节性变化。据研究,气候变暖可能是造成年间渔场重心变动的主要原因。Kang等[42]对1970年以来日本和韩国统计的日本鲭的捕捞情况进行了研究,指出在气候变化的大背景下,随着海温的升高,日本鲭的栖息地似乎正逐渐向北扩展。Cheung等[43]指出,海洋鱼类通常会前往高纬度和更深水域来应对海洋变暖,并提出了渔获物平均温度(Mean Temperature of the Catch,MTC)的概念来计量气候变暖对各海区产量的影响,得出在1970-2006年间,全球范围内非热带地区的MTC以0.19℃/(10 a)的速度增长,气候变暖已显著影响到全球渔业。本文月间产量重心变化的结论与戴澍蔚等[9]、徐博等[10]、崔国辰等[28]分别对2014年、2015年、2016-2017年的研究结果具有一致的变动规律,验证了西北太平洋日本鲭渔场重心具有明显季节性变动趋势的结论。月间渔场重心的变动与日本鲭自身的生态学特征和习性息息相关。日本鲭每年都会进行生殖和越冬的远距离洄游[11],每年的4-6月太平洋系群体在日本中部沿海进行产卵,而后向东北方向洄游至黑潮和亲潮的交汇区甚至到达更北的亲潮海域的索饵场进行索饵育肥[44-45]。海流对推动日本鲭渔场形成和消失、季节性变动起到了一定程度的影响[46]。黑潮暖流和亲潮寒流在西北太平洋海域交汇[47],大气与海洋强烈作用,水温合适,小型浮游生物众多,是多种小型中上层鱼类的重要索饵场,王良明等[11]指出,渔场位置会随两个水系强弱的不同而发生变化,同时表明,海流对于日本鲭渔场的作用机制还有待深入研究。另外,环境因子诸如海表面温度、水温的垂直结构等,对日本鲭渔场季节性变动也同样起着较大作用[11,31]。
本文利用SDE模型首次对西北太平洋渔场的变动情况进行了深入分析,可视化了渔场的变动方向和分布格局,得到了表征渔场变动的相关参数。结果表明,西北太平洋日本鲭渔场的扩张方向为严格的西南-东北格局,渔场各年份的扩张方向极为一致,且具有很强的向心力,结合产量分布、渔获网次核密度可以看出,每年在西北太平洋进行灯光围网作业的路线没有大的变化,即每年都延续上一年的捕捞路线。月间渔场方向性也较强,与年间的扩张方向一致,保持西南-东北格局,但在一致性上较年间弱-各月份的扩张方向稍有差异,另外,各月份渔场的扩张海域差别较大,从而验证了渔场具有明显季节性变化的结论。年间和月间的渔场变动方向均为严格的西南-东北格局,且相关研究也得到了相同的结论[9-10,28],究其原因,从表面上看主要受船队作业动态的影响,从更深层次来看,日本鲭渔场的变动是日本鲭聚集状态的结果,从而受其生态学特征及其习性的支配,但是具体原因仍有待深入研究。由于SDE模型在空间结构和空间区位研究上可视化的优越性,已在多个领域得到了应用[48],近年来也已被应用到渔业资源分布研究中,比如Guan等[49]分析了渤海湾13 种主要鱼类和无脊椎动物种群的分布,蔡研聪等[17]分析了南海北部近海苏门答腊金线鱼(Nemipterus mesoprion)资源的分布特征。本文将SDE模型首次应用到了西北太平洋日本鲭渔场分布的研究上,对远洋渔业资源的渔场分布提供了一种新的思路。
4.4 不足与展望
本文使用了地理上的空间自相关模型和空间统计方法中的SDE模型等多种时空分析方法,对西北太平洋日本鲭的渔场变动特征进行了深入的分析,得到了日本鲭渔场年间以及季节性的变化特征,对渔场的选择具有一定意义。但是本文还存在一定局限性:首先,本文主要从时间和空间上对日本鲭渔场的变动特征进行了研究,对于影响因素和驱动机制仅在讨论部分进行了剖析,而并未深入研究,在后续研究中应加强研究各影响因素的作用;其次,本文虽然研究了西北太平洋日本鲭渔场变动特征,但是并未对渔业的实际发展提出相关建议;最后,本文并未考虑数据本身可能存在的不确定性以及模型参数的敏感性,在以后研究中,应着重对其进行深入的分析,以使渔场分布特征分析的结果更可信。
猜你喜欢
中国水产(2021年12期)2021-12-06
大自然探索(2021年12期)2021-02-07
学生天地(2020年19期)2020-06-01
戏剧之家(2019年30期)2019-11-28
金山(2018年8期)2018-11-09
散文诗(2017年15期)2018-01-19
消费电子(2017年4期)2017-05-04
考试周刊(2016年37期)2016-05-30
火花(2015年7期)2015-02-27
中学生英语·阅读与写作(2008年7期)2008-12-19
