
2003~2018年干旱对中国森林绿度异常的影响
2022-01-21 02:11:44赵洪飞他志杰
中国环境科学 2022年1期
侯 鑫,赵 杰,赵洪飞,他志杰,岳 超,3*
2003~2018年干旱对中国森林绿度异常的影响
侯 鑫1,赵 杰1,赵洪飞1,他志杰2,岳 超1,3*
(1.西北农林科技大学资源环境学院,陕西 杨凌 712100;2.西安外国语大学旅游学院·人文地理研究所,陕西 西安 710128;3.中国科学院水利部水土保持研究所,陕西 杨凌 712100)
基于卫星观测和气象再分析数据提取2003~2018年间中国森林绿度异常现象(月尺度LAI长期趋势的累积偏差),并分析干旱对森林绿度异常影响的时空动态变化,探究不同森林类型的干旱敏感性.结果表明:①干旱导致的绿度异常现象频率分布呈现出南高北低的空间格局,而高强度异常现象主要分布于中国的东北与西南地区.②2003~2018年间绿度异常严重程度呈现显著增加趋势(即绿度异常强度指数下降),变化率为:-0.06/a(<0.05),而绿度异常面积比率下降,变化率为-0.0049/a.③中国森林对干旱胁迫较为敏感的区域主要分布于云南北部与大兴安岭北部,这是由区域的地质和气候特点以及植被类型共同作用导致的.④干旱主导区不同森林类型的干旱敏感性依次为:阔叶林>针叶林>混交林,绿度异常与SPEI回归斜率依次为1.8>1.3>1.2.⑤针叶林遭受的干旱强度最高,2003~2018年间统计的SPEI中位数为-1.65,绿度异常现象也最为严重,多年绿度异常指数中位数为-1.81,说明干旱强度而非敏感性主导了森林绿度异常的现象.
中国;绿度异常;干旱;森林;敏感性
干旱是影响森林绿度异常主要的胁迫因子之一[1],其通过影响植物光合作用与呼吸作用对森林的生长带来负面影响[2-3].持续性严重干旱甚至会造成森林大面积死亡[4].
不同森林类型对干旱的敏感性存在差异.Liu等[5]基于不同生物群落中极端低降水频率与植被绿度异常频率的回归斜率定义敏感性,发现阔叶林的干旱敏感性高于针叶林.Li等[6]通过树木年轮和植物性状数据,表明裸子植物具有更高的干旱抵抗力.与之相反的是,Huang等[7]的研究表明阔叶林在干旱期间保持原有增强型植被指数(EVI)水平的能力比针叶林更强.以上结果的差异说明有必要在全国范围内开展不同森林类型的干旱敏感性研究.
中国是干旱灾害发生频率最高且影响最严重的国家之一,近年我国南方地区干旱频次和强度明显增加[8-9],干旱已经造成了我国森林的大面积死亡[10].国内有关干旱胁迫对森林影响的研究多位于干旱与半干旱区域[11-13].由于干旱半干旱区的气候条件限制了森林的最大盖度[14],在大尺度研究中可被遥感产品识别的森林多位于湿润区.目前星载遥感技术已经被广泛应用于研究森林扰动[15-17],因此本研究采用一种新的方法通过2003~2018年间中分辨率成像光谱仪(MODIS)叶面积指数刻画干旱影响下中国森林绿度异常的时空动态,量化干旱对中国森林的影响,并更加深入的探究不同森林类型干旱敏感性差异,提高对于干旱引起的森林生长异常甚至死亡的风险认识;为森林管理者对森林实施区域化管理,制定合适的策略以提高森林生态系统韧性提供理论参考.
1 研究方法与数据来源
1.1 数据来源与预处理
叶面积指数(LAI)是核算生态系统光合作用、呼吸作用等过程的关键变量[18].作为全球植被的基本属性,LAI已经被全球气候变化研究界列为必需气候变量[19].McDowell等[20]的研究表明干旱时高温缺水的环境条件使得饱和水汽压差增加,致使树木叶面积减少以适应环境.Liu等[21]的研究也指出干旱导致了树木茎叶生长的减少.因此本研究选取LAI作为反应森林冠层状况的指标,用于定量描述植被绿度异常现象.Steltzer等[22]的研究发现不同植被类型的光合特性会影响归一化植被指数(NDVI)对叶面积的估算.本文数据选用美国国家航空航天局(NASA)发布的MODIS的叶面积指数遥感产品MCD15A2H(https://ladsweb.modaps.eosdis.nasa.gov/).产品算法选用美国宇航局的Terra和Aqua卫星在8d时段中的最佳影像数据.时间跨度为2003年1月~2018年12月,空间分辨率为500m,时间间隔为8d,共计736期影像.通过求均值将LAI的空间分辨率聚合到0.1°,保留原时间分辨率不变.
土地覆盖类型数据使用第6版MODIS土地覆盖类型(MCD12Q1)数据产品.该产品提供2003~ 2018年空间分辨率为500m的年尺度土地覆盖类型数据.该产品给出了基于6种不同土地覆盖分类方案的数据,本研究采用国际地球生物圈计划(IGBP)分类方案.选定土地覆盖类型中的1~5(常绿针叶林、常绿阔叶林、落叶针叶林、落叶阔叶林、混交林)作为森林.通过众数法将空间分辨率转为0.1°,获得0.1°的中国森林类型分布.
气象数据采用中国区域地面气象要素驱动数据集(CMFD),该数据是专门为研究中国陆地表层过程而开发的高时空分辨率近地表气象数据集[23].本研究采用的温度和降水数据时间跨度为1979~2018年,空间分辨率为0.1°,时间分辨率为1d.
利用CMFD气象数据集中的降水和温度估算了1979~2018年的标准化降水蒸散发指数(SPEI),选取其中的2003~2018年用作研究.SPEI是利用降水和潜在蒸散发(PET)之间的月差值来量化水分盈余或亏缺的程度,其中月尺度PET按照Thornthwaite[24]的方法估算获得,然后将月差值最终归一化为对数逻辑概率分布,得到SPEI序列[25].本研究中,SPEI的时间尺度选取为12个月,代表了数据所在月前1a截止至该月的累计水量平衡.长时间尺度(12个月)的SPEI表征更多的是水文干旱,能够反映深层土壤水分状态变化[26].森林具有较深的根系,可以汲取深层土壤水,使其更倾向于对长时间尺度的干旱产生响应,因此采用12月时间尺度的SPEI是适当的[7].
本研究将不同尺度的数据通过GIS实现数据的投影转换与重采样,使得数据空间分辨率统一为0.1°,投影统一为WGS84.通过将土地覆盖类型数据与LAI数据叠加,提取中国森林覆盖区域的LAI数据集.
1.2 研究方法
1.2.1 森林绿度异常数据的提取 绿度异常指的是在极端气候影响下,植被叶面积指数对于其长期生长趋势的负向偏离.由于人类土地利用和森林管理、气候变化、二氧化碳施肥以及氮沉降等诸多因素的作用,2000~2017年间中国森林LAI总体呈现增长趋势[27].因此本文采用去除线性趋势法,对LAI数据的每个步长(8d)去趋势.该方法除了去除LAI的时间增长趋势以外,也一并去除了森林生长的季节周期[28-29],便于观察年际间植被LAI的异常表现.对去趋势后LAI进一步统计获得最终绿度异常指数 (GAI),便于量化分析中国森林绿度异常的时空动态.
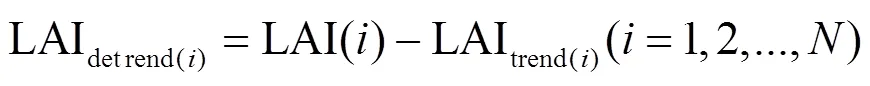
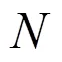


图1 中国某0.1°像元2005年去除线性趋势的值

1.2.2 干旱影响下绿度异常时空分异特征分析 森林的绿度异常现象可能由多种极端气候导致,为获取仅发生于干旱情况下的绿度异常,采用与Schwalm类似的方法,将SPEI<-1定义为像元出现中度或重度干旱[30],结合SPEI与绿度异常指数,提取干旱情况下的森林绿度异常现象.在此基础上,统计了2003~2018年间绿度异常的频率和强度,探究干旱导致绿度异常的频率和强度的空间分布特征

式中:表示多年间平均绿度异常强度,表示2003~ 2018年间绿度异常现象出现的年份次数(即频率).
为了探究干旱情况下发生森林绿度异常的面积、强度的时间变化趋势,累加获取全国干旱情况下逐年绿度异常面积比率(绿度异常面积与森林总面积的比值),并求取每年全国绿度异常强度的均值,采用线性回归分析(最小二乘法拟合)获取随时间的变化趋势.
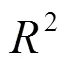
2 结果与分析
2.1 我国森林类型空间分布特征
如图2所示,我国森林空间分布差异化明显,针叶林主要分布于大兴安岭北部以及四川省西南部.混交林主要分布我国中部一带,在我国云南东北部也存在混交林聚集分布.阔叶林分布最为广泛,北至我国黑龙江、吉林、辽宁一带,中至在我国陕西省南部,同时我国贵州、广西、广东、福建、江西几省主要森林类型均为阔叶林.

图2 中国森林类型的空间分布
基于国家测绘地理信息局标准地图服务网站下载的审图号为GS(2020)4632号的标准地图制作,底图无修改,下同
2.2 干旱影响下我国森林绿度异常时空分异特征
森林绿度异常现象可能由多种极端气候事件(干旱、台风)驱动,研究仅保留干旱情况下(SPEI<-1)出现的森林绿度异常.2003~2018年间,累计出现绿度异常现象的森林面积约有606万km2,说明干旱是致使树木生长异常的重要因素.本文结果成功刻画了受到广泛关注与报道的我国森林受干旱影响的事件(图3),包括:2004年我国南方发生大范围绿度异常现象,涉及广东、浙江、福建和江西省[32].全国森林受干旱影响(即出现绿度异常)总面积约63万km2,平均绿度异常指数为0.98,森林出现较大面积绿度异常现象但程度较轻.2010年我国云南发生了严重的绿度异常现象[33].绿度异常总面积约47万km2,像元平均绿度异常指数为-0.72,即干旱程度低于通常严重程度约一个标准差,受灾森林LAI显著降低.2012, 2013年云南地区又连续发生严重的绿度异常现象[33-34].2018年我国东南部(广东、福建和湖南)出现绿度异常现象[35].总体绿度异常面积约47万km2,平均绿度异常指数为-0.25.东南地区出现绿度异常面积较大但强度较低.这些绿度异常事件的成功识别表明本研究中所用绿度异常的刻画方法具有较强的可靠性.

图3 干旱条件下森林绿度异常分布
2003~2018年间我国干旱情况下绿度异常强度较高(<0)的区域主要分布在藏东南、云南省大部、四川南部、贵州广西和湖南三省交界、华北地区西部以及大兴安岭一带(图4a).绿度异常强度较高的森林面积占全部有林区的31.05%.绿度异常强度总体上呈现出东北与西南较高,其他地区较低的空间格局.2003~2018年16a间,绿度异常频率较高(出现次数>4)的区域多位于云南西部与东部.此外,西藏东部、四川南部、湖北与重庆交界以及浙江与安徽交界处也有少量绿度异常现象聚集(图4b).绿度异常频率较高的像元面积占总体有林区的45.71%.空间分布呈现出南高北低的格局并且高频像元集中于我国西南一带.这表明我国南方尤其是西南地区森林受干旱影响明显较为严重,绿度异常频率与强度均较高.干旱情况下极端绿度异常(I<-2)的像元2003~2018年间出现频率均低于3次,干旱频次7次以上的像元未出现过极端绿度异常现象,这意味着高频率与高强度绿度异常事件往往不在同一区域同时发生.

图4 干旱条件下森林绿度异常强度与频率分布

图5 干旱条件下森林绿度异常的时间变化趋势

2.3 绿度异常受干旱影响的空间格局
森林绿度异常指数与SPEI的回归拟合优度(<0.05)空间差异化明显.SPEI对于云南省大部、大兴安岭北部以及广东、福建和浙江南部的绿度异常解释程度较高(图6a),表明上述地区森林冠层叶面积指数与水分条件的耦合程度更高.回归斜率则表明绿度异常随SPEI的变化,即绿度异常对干旱的敏感性.其中云南省北部与大兴安岭北部斜率为正值,表明随着干旱程度加重绿度异常加重.广东、福建和浙江南部斜率为负值,则说明随着干旱的出现绿度异常现象反而减轻(图6b),干旱不是森林绿度异常的主要驱动因素.结合两者判断,云南省北部和大兴安岭北部和中部的森林绿度异常受到干旱影响较为严重.
2.4 不同森林类型绿度异常及对干旱敏感性的差异
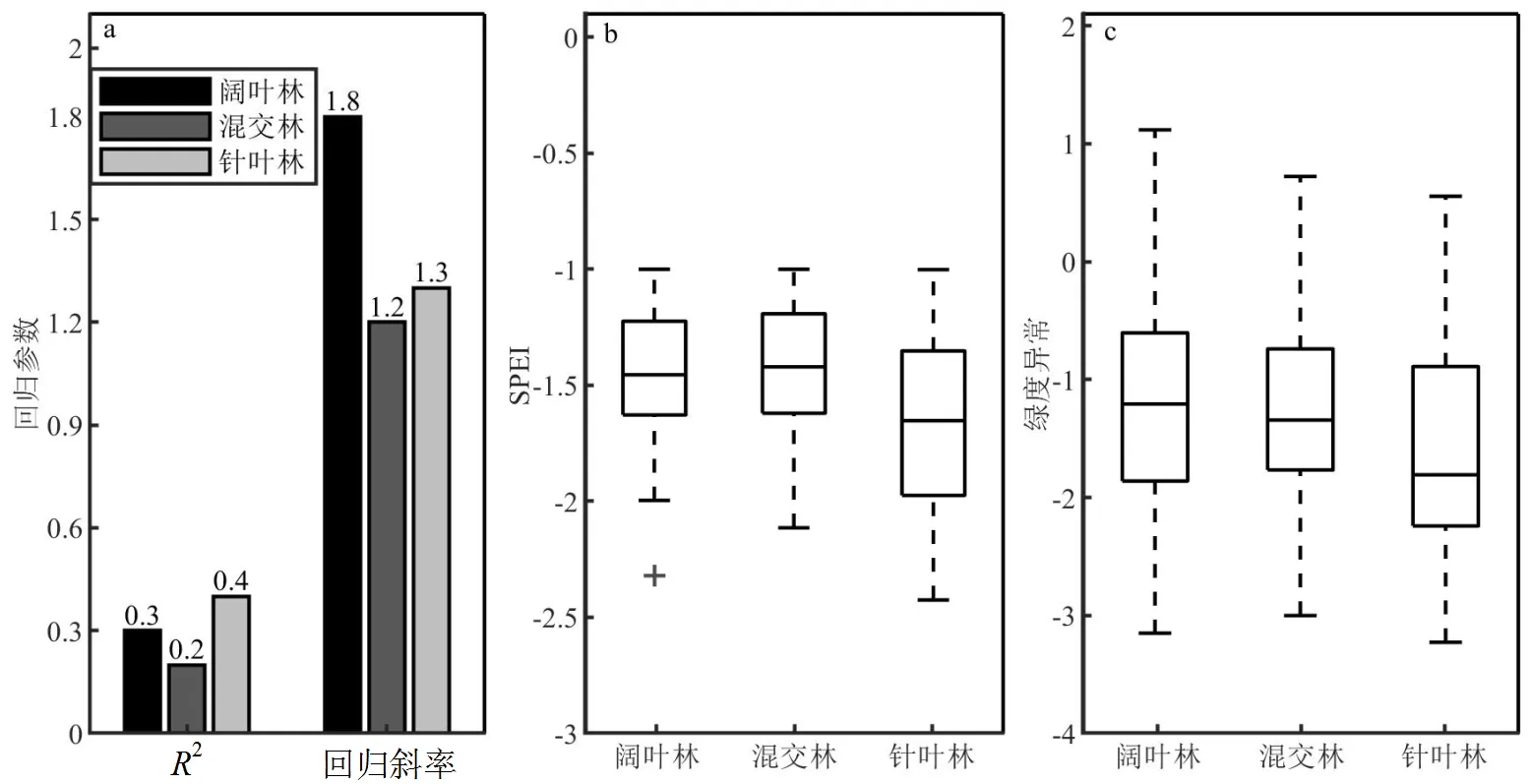
图7 2003~2018年SPEI分布与干旱导致的绿度异常指数分布统计
在分析不同森林类型的干旱敏感性时将研究区限制在干旱主导区,即上述绿度异常受干旱影响的空间格局中回归分析的2>0.4,且回归斜率为正值的区域,以排除其他极端气候事件的干扰.按照不同森林类型划分干旱主导区的绿度异常数据,并通过线性回归斜率的大小来表征不同森林类型对干旱的敏感性.图7a中阔叶林的回归斜率为1.8,针叶林的回归斜率为1.3,混交林的回归斜率为1.2.对应不同森林类型的敏感性强弱依次为:阔叶林>针叶林>混交林.这表明处于相同的干旱强度时,针叶林的绿度异常现象要比阔叶林轻微,混交林绿度异常表现最小.与干旱敏感性的结果相反,阔叶林的多年绿度异常指数中位数为-1.21,针叶林的多年绿度异常指数中位数为-1.81(图7c).这说明2003~2018年间干旱敏感性较低的针叶林实际产生的绿度异常现象比阔叶林更加严重.图7b的结果部分解释了这一现象:阔叶林多年间SPEI值的中位数为-1.45,针叶林多年间SPEI值的中位数为-1.65.表明2003~2018年间,针叶林区域的干旱强度相对阔叶林区域更高.意味着可能干旱强度主导了森林绿度异常表现,而非干旱敏感性.
3 讨论
为了探究干旱对于森林绿度异常的影响,结合SPEI与绿度异常指数获得我国2003~2018年干旱影响下绿度异常的空间分布.结果表明干旱影响下绿度异常现象的频率与强度具有明显的空间异质性.极端绿度异常现象往往发生于干旱频率相对较低的区域.这可能是由于这些区域的植被缺少对于极端水分亏缺的应对机制,这种现象同样有可能发生于干旱适应性强的裸子植物[36].对绿度异常时间变化趋势的研究表明,绿度异常的强度显著增强.这可能是由于我国南方极端干旱事件明显增多,西南地区特大干旱事件频发,干旱中心的南移使得中度到重度干旱事件的频率与强度增加[37].绿度异常面积的下降趋势可能是由于面积变化存在周期性波动,王强等[17]统计的农作物旱灾受灾面积在1951~ 2015年存在总体增大趋势,在这期间的2000~2015年受灾面积却表现出显著下降趋势.本文森林干旱影响下绿度异常面积可能与之相似,选取的时间序列处于大周期的波动下降阶段.
绿度异常的干旱主导区主要分布于云南省北部和大兴安岭北部和中部.从云南地质和气候特点角度考虑,云南地区干季持续时间长,从11月到次年4月,长达6个月[38],干旱事件持续时间长,强度大[9].该地区的岩溶地貌地表水易下渗成为地下水,使得土壤水分难以保持,容易导致土壤水分亏缺[39].从森林类型角度考虑,我国南方多为被子植物,相较于裸子植物对干旱的抵抗能力更弱[6,40].这些条件共同造成了云南北部干旱主导区的集中现象.大兴安岭中部和北部地区干季为11月到次年3月,占年降水量的约10%~12%[41].落叶松是大兴安岭森林生态系统的优势树种.大兴安岭干旱主导区单一的生态系统格局导致物种多样性较低,减弱了对于干旱的抵抗力[42].虽然研究普遍认为裸子植物本身相较于被子植物具有更高的抵抗力,但森林在频繁或长期的干旱下,仍有可能产生绿度异常现象[10,6,36,43-44].另一方面,大兴安岭区域的绿度异常可能来自于火灾这种通常与干旱并发的灾害.火灾是兴安落叶松生长的主要干扰之一.由于干旱导致的气温升高、枯落物堆积以及盛行强风,使得春季很容易发生森林大火[45].
我国东南部沿海广东、福建和浙江南部的SPEI与绿度异常同样存在高相关性,但SPEI与绿度异常呈现负相关.这可能是由于该区域辐射为森林生长的主要限制因子[10].干旱情况下降水减少,随着蒸散发降低,更透明的非降雨云与云量降低使得太阳辐射强度增加,促进了植被生长[46]. Morimoto等[47]的研究表明降水是预测伴随暴雨的台风灾害对森林干扰的重要变量,结合该区域的绿度异常与水热条件的高2表现(图6a)以及该区域台风灾害频发的特点[48],伴随暴雨的台风灾害可能是该区域森林绿度异常的主要驱动因素.
研究通过SPEI与绿度异常数据的回归斜率来表征干旱敏感性,以保证在同一干旱严重程度下展开分析[5].结果表明针叶林对干旱的敏感性更低,这可能源于针叶林木质部水力特征使其在干旱情况下不易产生空化或者栓塞[6].除此之外,其能通过调节气孔开闭控制水分流失[49].该结果与Huang等[7]的差异可能因为其对干旱敏感性的定义未考虑森林是否处于同一干旱强度下.不同森林类型2003~ 2018年间的总体绿度异常表明,针叶林在干旱期间保持原有LAI水平的能力弱于阔叶林,这与两者的敏感性表现相反.这说明植被LAI对于干旱事件的响应可能受到干旱强度、环境和植被敏感性等因素的复合影响[7].图7b也印证了多年间针叶林遭受的干旱程度更高.在更高强度的干旱下,抵抗力更强的针叶林表现出比阔叶林更严重的异常现象.
本研究还存在一些不足之处.首先,限于目前气象数据的空间分辨率较低以及遥感数据的精度限制无法在大尺度上开展更为精细的相关研究.其次,本研究仅通过SPEI所表征的水量平衡来定义干旱,具有一定局限性.因为绿度异常可能是由任何与干旱同时发生的次生灾害(如火灾、虫灾)造成的[50].更为复杂的干旱影响机制需要进一步细化干旱情况下绿度异常的驱动因素.因此,今后有必要结合其他森林干扰分布数据厘清干旱对森林绿度异常的影响及其驱动机制.
4 结论
4.1 干旱情况下中国东北与西南地区绿度异常强度较高,绿度异常频率空间分布呈现出南高北低的格局.
4.2 干旱导致的绿度异常严重程度呈显著上升趋势,变化率为-0.06/a,绿度异常面积比率呈下降趋势,变化率为-0.0049/a.
4.3 中国森林绿度异常的干旱主导区集中于云南省北部和大兴安岭北部.
4.4 干旱主导区不同森林类型对于干旱的敏感性为:阔叶林>针叶林>混交林,3种森林类型绿度异常与SPEI回归斜率依次为1.8>1.3>1.2.2003~2018年间干旱主导区总体干旱强度为:针叶林>阔叶林>混交林,统计的SPEI中位数依次为:-1.65<-1.45<-1.42.针叶林总体干旱强度最高,阔叶林与混交林相当.多年间由干旱导致的森林绿度异常现象强度为:针叶林>阔叶林>混交林,绿度异常指数中位数依次为:-1.21> -1.34>-1.81.针叶林由干旱导致的绿度异常现象最为严重,阔叶林和混交林表现相当.受干旱强度影响,总体绿度异常表现与敏感性并不完全一致.
[1] Seidl R, Thom D, Kautz M, et al. Forest disturbances under climate change [J]. Nature Climate Change, 2017,7(6):395-402.
[2] Griffin J J, Ranney T G, Pharr D M. Heat and drought influence photosynthesis, water relations, and soluble carbohydrates of two ecotypes of redbud (Cercis canadensis) [J]. Journal of the American Society for Horticultural Science, 2004,129(4):497-502.
[3] Ciais P, Reichstein M, Viovy N, et al. Europe-wide reduction in primary productivity caused by the heat and drought in 2003 [J]. Nature, 2005,437(7058):529-533.
[4] Young D J, Stevens J T, Earles J M, et al. Long‐term climate and competition explain forest mortality patterns under extreme drought [J]. Ecology letters, 2017,20(1):78-86.
[5] Liu G, Liu H, Yin Y. Global patterns of NDVI-indicated vegetation extremes and their sensitivity to climate extremes [J]. Environmental Research Letters, 2013,8(2):279-288.
[6] Li X, Piao S, Wang K, et al. Temporal trade-off between gymnosperm resistance and resilience increases forest sensitivity to extreme drought [J]. Nature Ecology & Evolution, 2020,4(8):1075-1083.
[7] Huang K, Xia J. High ecosystem stability of evergreen broadleaf forests under severe droughts [J]. Global Change Biology, 2019,25(10): 3494-3503.
[8] 任国玉,封国林,严中伟.中国极端气候变化观测研究回顾与展望 [J]. 气候与环境研究, 2010,15(4):337-353.
Ren G Y, Feng G L, Yan Z W, et al. Progresses in observation studies of climate extremes and changes in mainland China [J]. Climatic and Environmental Research, 2010,15(4):337-353.
[9] 张 强,姚玉璧,李耀辉,等.中国干旱事件成因和变化规律的研究进展与展望 [J]. 气象学报, 2020,78(3):500-521.
Zhang Q, Yao Y B, Li Y H, et al. Progress and prospect on the study of causes and variation regularity of droughts in China [J]. Acta Meteorologica Sinica, 2020,78(3):500-521.
[10] Allen C D, Macalady A K, Chenchouni H, et al. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests [J]. Forest Ecology and Management, 2010, 259(4):660-684.
[11] 何志斌,杜 军,陈龙飞,等.干旱区山地森林生态水文研究进展 [J]. 地球科学进展, 2016,31(10):1078-1089.
He Z B, Du J, Chen L F, et al. Review on montane forest eco- hydrology in arid area [J]. Advances in Earth Science, 2016,31(10): 1078-1089.
[12] 赵鹏武,管立娟,刘兵兵,等.我国半干旱区东段森林动态研究现状及展望 [J]. 世界林业研究, 2021,34(2):74-79.
Zhao P W, Guan L J, Liu B B, et al. Current research and prospect of forest dynamics in eastern section of Semi-arid Area in China [J]. World Forestry Research, 2021,34(2):74-79.
[13] 郭永强,王乃江,褚晓升,等.基于Google Earth Engine分析黄土高原植被覆盖变化及原因 [J]. 中国环境科学, 2019,39(11):4804-4811.
Guo Y Q, Wang N J, Chu X S, et al. Analyzing vegetation coverage changes and its reasons on the Loess Plateau based on Google Earth Engine. China Environmental Science, 2019,39(11):4804-4811.
[14] Gunin P D, Vostokova E A, Dorofeyuk N I, et al. Vegetation dynamics of Mongolia [M]. Springer Science & Business Media, 2013:79-129.
[15] Chen J, Sun L. Using MODIS EVI to detect vegetation damage caused by the 2008 ice and snow storms in south China [J]. Journal of Geophysical Research: Biogeosciences, 2010,115(G3):1-12.
[16] Hansen M C, Potapov P V, Moore R, et al. High-resolution global maps of 21st-century forest cover change [J]. Science, 2013,342 (6160):850-853.
[17] 王 强,张 勃,戴声佩,等.三北防护林工程区植被覆盖变化与影响因子分析 [J]. 中国环境科学, 2012,32(7):1302-1308.
Wang Q, Zhang B, Dai S P, et al. Analysis of the vegetation cover chang and its relationship with factors in the Three-North Shelter Forest Program [J]. China Environmental Science, 2012,32(7):1302- 1308.
[18] Fang H, Baret F, Plummer S, et al. An overview of global leaf area index (LAI): Methods, products, validation, and applications [J]. Reviews of Geophysics, 2019,57(3):739-799.
[19] Mason P, Zillman J, Simmons A, et al. Implementation plan for the global observing system for climate in support of the UNFCCC (2010Update) [Z]. GCOS Rep. 2009:28-29.
[20] Mcdowell N G, Allen C D. Darcy's law predicts widespread forest mortality under climate warming [J]. Nature Climate Change, 2015, 5(7):669-672.
[21] Liu J F, Arend M, Yang W J, et al. Effects of drought on leaf carbon source and growth of European beech are modulated by soil type [J]. Scientific Reports, 2017,7(1):1-9.
[22] Steltzer H, Welker J M. Modeling the effect of photosynthetic vegetation properties on the NDVI–LAI relationship [J]. Ecology, 2006,87(11):2765-2772.
[23] Yang K, He J, Tang W J, et al. On downward shortwave and longwave radiations over high altitude regions: Observation and modeling in the Tibetan Plateau [J]. Agricultural and Forest Meteorology, 2010,150(1): 38-46.
[24] Thornthwaite C W. An approach toward a rational classification of climate [J]. Geographical Review, 1948,38(1):55-94.
[25] Vicente-Serrano S M, Beguería S, López-Moreno J I. A multiscalar drought index sensitive to global warming: the standardized precipitation evapotranspiration index [J]. Journal of Climate, 2010, 23(7):1696-1718.
[26] Wang H, Vicente-Serrano S M, Tao F, et al. Monitoring winter wheat drought threat in Northern China using multiple climate-based drought indices and soil moisture during 2000~2013 [J]. Agricultural and Forest Meteorology, 2016,228:1-12.
[27] Chen C, Park T, Wang X, et al. China and India lead in greening of the world through land-use management [J]. Nature Sustainability, 2019,2(2):122-129.
[28] Zhang Y, Liu C, Tang Y, et al. Trends in pan evaporation and reference and actual evapotranspiration across the Tibetan Plateau [J]. Journal of Geophysical Research: Atmospheres, 2007,112:D11210.
[29] Cuo L, Zhang Y, Gao Y, et al. The impacts of climate change and land cover/use transition on the hydrology in the upper Yellow River Basin, China [J]. Journal of Hydrology, 2013,502:37-52.
[30] Schwalm C R, Anderegg W R, Michalak A M, et al. Global patterns of drought recovery [J]. Nature, 2017,548(7666):202-205.
[31] 赵 杰,刘雪佳,杜自强,等.昼夜增温速率的不对称性对新疆地区植被动态的影响 [J]. 中国环境科学, 2017,37(6):2316-2321.
Zhao J, Liu X J, Du Z Q, et al. Effects of the asymmetric diurnal- warming on vegetation dynamics in Xinjiang [J]. China Environmental Science, 2017,37(6):2316-2321.
[32] 王维国,饶晓琴,康志明,等.2004年我国重大灾害性天气和大尺度环流特征 [J]. 气象, 2005,31(5):26-31.
Wang W G, Rao X Q, Kang Z M, et al. Analysis of the calamitous weather and large-scale circulation in 2004 [J]. Meteorological Monthly, 2005,31(5):26-31.
[33] 黄荣辉,刘 永,王 林,等.2009年秋至2010年春我国西南地区严重干旱的成因分析 [J]. 大气科学, 2012,36(3):443-457.
Huang R H, Liu Y, Wang L, et al. Analyses of the causes of severe drought occurring in Southwest China from the fall of 2009 to the Spring of 2010 [J]. Chinese Journal of Atmospheric Sciences, 2012, 36(3):443-457.
[34] 段海霞,王素萍,冯建英.2013年全国干旱状况及其影响与成因 [J]. 干旱气象, 2014,32(2):310-316.
Duan H X, Wang S P, Feng J Y. Drought events and its influence in 2013 in China [J]. Journal of Arid Meteorology, 2014,32(2):310-316.
[35] 张 宇,王芝兰,沙 莎,等.2018年夏季全国干旱状况及其成因 [J]. 干旱气象, 2018,36(5):884-892.
Zhang Y, Wang Z L, Sha S, et al. Drought events and its causes in Summer of 2018 in China [J]. Journal of Arid Meteorology, 2018, 36(5):884-892.
[36] Zhang X, Li X, Manzanedo R D, et al. High risk of growth cessation of planted larch under extreme drought [J]. Environmental Research Letters, 2021,16(1):014040.
[37] 韩兰英,张 强,贾建英,等.气候变暖背景下中国干旱强度、频次和持续时间及其南北差异性 [J]. 中国沙漠, 2019,39(5):1-10.
Han L Y, Zhang Q, Jia J Y, et al. Drought severity, frequency, duration and regional differences in China [J]. Journal of Desert Research, 2019,39(5):1-10.
[38] 黄中艳.1961~2007年云南干季干湿气候变化研究 [J]. 气候变化研究进展, 2010,6(2):113-118.
Huang Z Y. Dry-wet climate change of the dry season in Yunnan During 1961~2007 [J]. Climate Change Research, 2010,6(2):113-118.
[39] Liu M, Xu X, Wang D, et al. Karst catchments exhibited higher degradation stress from climate change than the non-karst catchments in southwest China: An ecohydrological perspective [J]. Journal of Hydrology, 2016,535:173-180.
[40] Lü L, Cai H, Yang Y, et al. Geographic patterns and environmental determinants of gymnosperm species diversity in China [J]. Biodiversity Science, 2018,26(11):1133-1146.
[41] Jiang Y, Zhang J, Han S, et al. Radial growth response of larix gmelinii to climate along a latitudinal gradient in the Greater Khingan Mountains, Northeastern China [J]. Forests, 2016,7(12):295.
[42] Grossiord C, Granier A, Ratcliffe S, et al. Tree diversity does not always improve resistance of forest ecosystems to drought [J]. Proceedings of the National Academy of Sciences, 2014,111(41): 14812-14815.
[43] Trugman A T, Detto M, Bartlett M K, et al. Tree carbon allocation explains forest drought-kill and recovery patterns [J]. Ecology Letters, 2018,21(10):1552-1560.
[44] Li X, Li Y, Chen A, et al. The impact of the 2009/2010 drought on vegetation growth and terrestrial carbon balance in Southwest China [J]. Agricultural and Forest Meteorology, 2019,269:239-248.
[45] Fan Q, Wang C, Zhang D, et al. Environmental influences on forest fire regime in the Greater Hinggan Mountains, Northeast China [J]. Forests, 2017,8(10):372.
[46] Josefsson W, Landelius T. Effect of clouds on UV irradiance: As estimated from cloud amount, cloud type, precipitation, global radiation and sunshine duration [J]. Journal of Geophysical Research: Atmospheres, 2000,105(D4):4927-4935.
[47] Morimoto J, Aiba M, Furukawa F, et al. Risk assessment of forest disturbance by typhoons with heavy precipitation in northern Japan [J]. Forest Ecology and Management, 2021,479:118521.
[48] 殷 洁,戴尔阜,吴绍洪.中国台风灾害综合风险评估与区划 [J]. 地理科学, 2013,33(11):1370-1376.
Yin J, Dai E F, Wu S H. Integrated risk assessment and zoning of typhoon disasters in China [J]. Scientia Geographica Sinica, 2013, 33(11):1370-1376.
[49] Brodribb T J, Mcadam S A M, Jordan G J, et al. Conifer species adapt to low-rainfall climates by following one of two divergent pathways [J]. Proceedings of the National Academy of Sciences, 2014,111(40): 14489-14493.
[50] 马明卫,韩宇平,严登华,等.特大干旱事件灾害孕育机理及影响研究进展 [J]. 水资源保护, 2020,36(5):11-21.
Ma M W, Han Y P, Yan D H, et al. Research progress on the mechanism and influence of extreme drought-induced disasters [J]. Water Resources Protection, 2020,36(5):11-21.
Drought effects on forest greenness anomalies in China from 2003 to 2018.
HOU Xin1, ZHAO Jie1, ZHAO Hong-fei1, TA Zhi-jie2, YUE Chao1,3*
(1.College of Natural Resources and Environment, North West Agriculture and Forestry University, Yangling 712100, China;2.School of Tourism & Research Institute of Human Geography, Xi'an International Studies University, Xi'an 710128, China;3.Institute of Soil and Water Conservation, CAS & MWR, Yangling 712100, China)., 2022,42(1):336~344
This study was to evaluate the effects of drought on forest greenness anomalies in China. Satellite observations of forest leaf area index (LAI) and climate reanalysis data were used to examine forest canopy damage induced by drought during the period from 2003 to 2018. We assigned the forest Greenness Anomaly index (GAI) as a proxy for the forest canopy damage, defined as cumulative monthly negative LAI anomalies deviated from the long-term monthly LAI trend, then followed by standardization using the standard deviation. Using this index, we analyzed the spatiotemporal dynamics of drought-induced forest damage and the drought sensitivity of different forest types. Our results showed that: (1) The drought-induced forest canopy damage was more widespread in South China than in North China, while high-intensity drought damage mainly occurred in the Northeast and Southwest of China; (2) From 2003 to 2018, the severity of drought-induced canopy damage increased significantly with the GAI change rate of -0.06/a (a lower index means a higher severity), while the forest area ratio subjected to the drought damage decreased with the change rate of -0.0049/a; (3) Forests with a high sensitivity of drought damage mainly occurred in the north of Yunnan and the north part of the Greater Khingan Mountains in China, due to the common effects of geological, climate and vegetation type characteristics of the region; (4) Broadleaf forest was most sensitive to drought, followed by coniferous forest, while mixed forest was the least sensitive forest type and the corresponding regression slopes of GAI and SPEI for these three forest types were 1.8, 1.3 and 1.2, respectively; and (5) from 2003 to 2018, coniferous forest suffered the highest drought intensity and forest damage, with the median of SPEI as -1.65. As a result forest damage was more dominated by the drought intensity than by drought sensitivity.
China;greenness anomaly;drought;forest;sensitivity
X173
A
1000-6923(2022)01-0336-09
侯 鑫(1995-),男,河南新乡人,西北农林科技大学硕士研究生,主要从事气候变化与植被响应方向研究.
2021-06-08
陕西省自然科学基础研究计划(2021JQ-771)
* 责任作者, 研究员, chaoyue@ms.iswc.ac.cn
猜你喜欢
水土保持研究(2024年2期)2024-03-18 07:58:20
中国环境科学(2023年12期)2024-01-06 01:15:02
节水灌溉(2023年1期)2023-02-03 12:53:36
现代农业研究(2018年4期)2018-08-11 10:25:02
中国绿色画报(2018年5期)2018-08-07 07:12:26
森林工程(2018年4期)2018-08-04 03:23:16
丝路艺术(2018年1期)2018-04-01 18:10:15
电子制作(2017年7期)2017-06-05 09:36:14
海洋气象学报(2016年3期)2016-02-28 14:27:42
食管疾病(2015年3期)2015-12-05 01:45:11
