
三角梅芽变突变体光合特性分析
2022-01-18 14:31杜学林刘悦明余铭杰黄子锋蔡振远王凤兰
热带农业科技 2022年1期
杜学林,刘悦明,余铭杰,黄子锋,蔡振远,王凤兰*
(1.仲恺农业工程学院,广州510225;2.广州市林业和园林科学研究院,广州510405;3.东莞市农业科学研究中心,广东东莞523083)
叶色突变是很多植物都存在的常见现象,主要特征为绿色叶的黄化、白化或产生斑点及条纹等[1]。导致植物叶色突变的原因众多,涉及机制复杂,其内在因素包括光合色素合成相关基因突变、叶绿素形成过程中相关基因突变等;外部环境则会受到光照、温度、外源激素等的影响[2]。开展叶色突变体的研究对探究植物光形态建成、叶绿素合成和代谢以及叶绿体发育等具有重要意义;也可作为标记性状用于辅助育种及杂种一代的培育[3-6],不仅能提高育种效率,提高种子纯度[7],还有可能带来更大的经济效益。因此,很多学者对植物叶色突变现象开展过一系列研究,目 前 已 涵 盖 了 拟 南 芥[8]、水 稻[9]、黄 瓜[10]、兰花[11]、桑树[12]、大豆[13]、烟草[14]等。
三角梅中也存在叶色突变的现象,其叶色突变形式一般以黄化、白化为主,在叶形态上也会有出现变小的情况。目前关于三角梅芽变突变体的研究较少,多集中在人工杂交[15-16]、抗性[17-19]、栽培管理[20-22]等方面。本文以3年生三角梅芽变突变体植株上的黄化叶片以及正常叶片为材料,对处于生长期的2种叶片进行光合色素含量和叶绿素荧光参数测定,并对突变植株进行园艺性状调查,探讨了正常叶片和黄化叶片之间的光合性能差异,为三角梅叶色突变相关研究奠定基础。
1 材料和方法
1.1 试验材料
试材于2021年1月取自广州市林业和园林科学研究院白云苗圃防雨大棚,选取3年生有芽变现象的三角梅植株,共3个品种,分别为:银边浅紫(Bougainvillea glabra‘Mrs Eva Mauve Variegata’)、金边浅紫(Bougainvillea glabra‘Mrs Eva Mauve VariegataⅡ’)、金 边 水 红(Bougainvilleaxbuttiana‘Miss Manila Variegata’)。
1.2 试验方法
1.2.1 园艺性状观测
在三角梅生长期间,观测比较叶色突变体植株黄化枝条与正常枝条的叶色变化、茎粗、叶长、叶宽等园艺性状。3次重复,每个重复测定2株。
1.2.2 光合色素含量测定
采用王力源等[23]测定光合色素的方法。分别选取三角梅芽变植株上黄化枝条与正常枝条上4~7叶位(从顶芽依次往下数)的功能性叶片,放入冰盒内带回试验室,蒸馏水洗净,吸干表面水分。将叶片剪碎混合,各称取0.05 g,放入10 mL离心管中,加入10 mL 95 %乙醇,用黑色塑料袋密封保存,黑暗避光浸提至叶片完全变白,然后用分光光度计分别测定在663 nm、645 nm和470 nm下的OD值。每个处理3次重复。根据公式分别计算叶绿素a(Chla)、叶绿素b(Chlb)、类胡萝卜素(Caro)以及总叶绿素(Chla+b)的含量。计算公式如下:
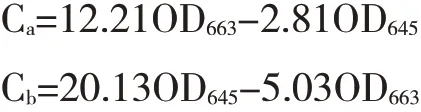
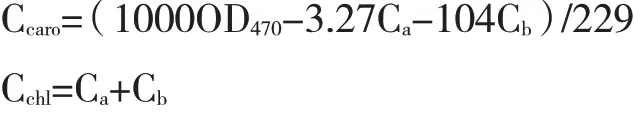
1.2.3 叶绿素荧光参数测定
在晴朗天气的9:00—12:00时间段内,选取三角梅芽变植株上正常枝条与黄化枝条上4~7叶位的功能性叶片进行黑暗处理20 min,然后用Opti-Sciences OS30p+手持叶绿素荧光仪分别测定三角梅芽变植株的正常叶片与黄化叶片的初始荧光(Fo),最大荧光(Fm)以及可变荧光(Fv),从而得出光系统Ⅱ(PSⅡ)原初光化学效率(Fv/Fm)、光系统Ⅱ(PSⅡ)的潜在光化学活性(Fv/Fo)。
1.2.4 数据分析方法
应用WPS Office和SPSS 26软件对试验数据进行分析,SigmaPlot 10.0软件制图。采用独立样本t检验法分析三角梅芽变突变体的变异情况及不同指标之间的显著性差异。
2 结果与分析
2.1 园艺性状比较
通过田间实地测量观察,与正常枝条相比,芽变枝条整枝布满黄色叶片,且叶色在整个生长过程中不会逆转。不同品种叶色黄化程度不同,其中金边浅紫品种呈金黄色,其余则呈黄偏白色。不同品种的新芽表现为不同颜色,金边水红芽变枝条新芽呈黄棕色,且新芽长大后逐渐偏粉红色,后又转为黄色,而正常枝条则表现为叶边缘粉红色中部暗绿色(图1a);金边浅紫芽变新芽呈黄棕色,正常枝条则呈中部绿色,边缘带少量棕色(图1b);银边浅紫芽变枝条新芽呈黄色,正常枝条新芽则为中部绿色,边缘带不规则棕黄色(图1c)。芽变枝条落叶情况比正常枝条严重,个别黄化枝条在1月叶片几乎掉光,而正常枝条仍然生长良好。

图1 芽变与正常枝条新芽叶色比较
从观测结果(表1)来看,每个品种芽变植株正常枝条与黄化枝条在茎粗、叶长、叶宽等方面差异不显著。说明三角梅芽变现象的产生不会影响芽变枝条的生长发育,推测其原因可能是芽变枝条非单独存在,仍可吸收母体植株的营养成分而维持正常生长发育。
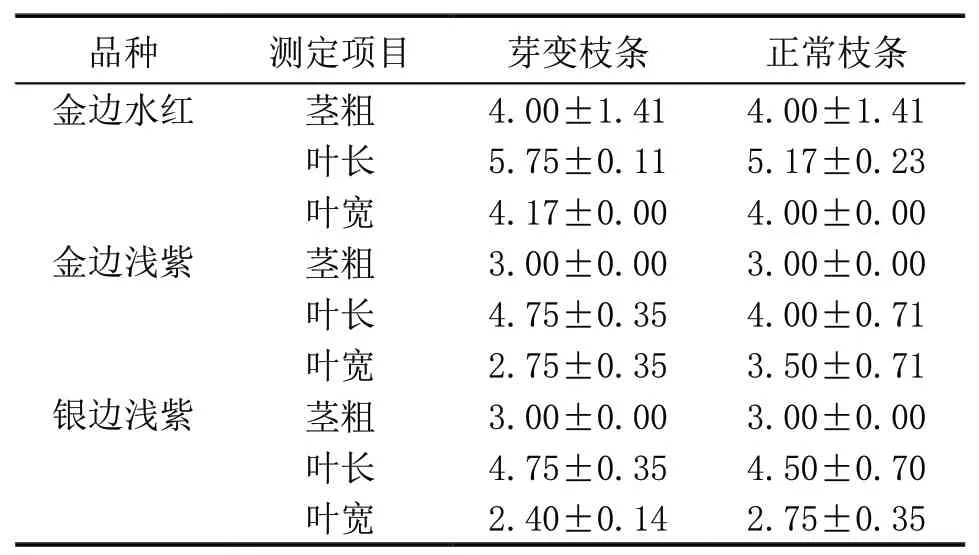
表1 三角梅正常与芽变枝条形态指标的比较 cm
2.2 光合色素含量差异分析
从表2可见,3种三角梅芽变植株黄化叶片与正常叶片的各光合色素含量均差异显著。黄化叶片的叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量均低于正常叶片。与正常叶片相比,金边水红黄化叶片的叶绿素a和叶绿素b分别降低了97.52 %和78.26 %,总叶绿素和类胡萝卜素含量的则分别降低了94.57%和96.64 %。金边浅紫黄化叶片的叶绿素a、叶绿素b和总叶绿素含量均低于正常叶片,分别降低65.49%、47.62%和60.65%;银边浅紫中黄化叶片与正常叶片相比,叶绿素a与叶绿素b分别降低了95.60%和80.58%,总叶绿素则降低了92.83%,而类胡萝卜素差异最明显,减少了98.54%。推测三角梅芽变产生黄化叶片的原因可能与上述3种光合色素的减少有关。
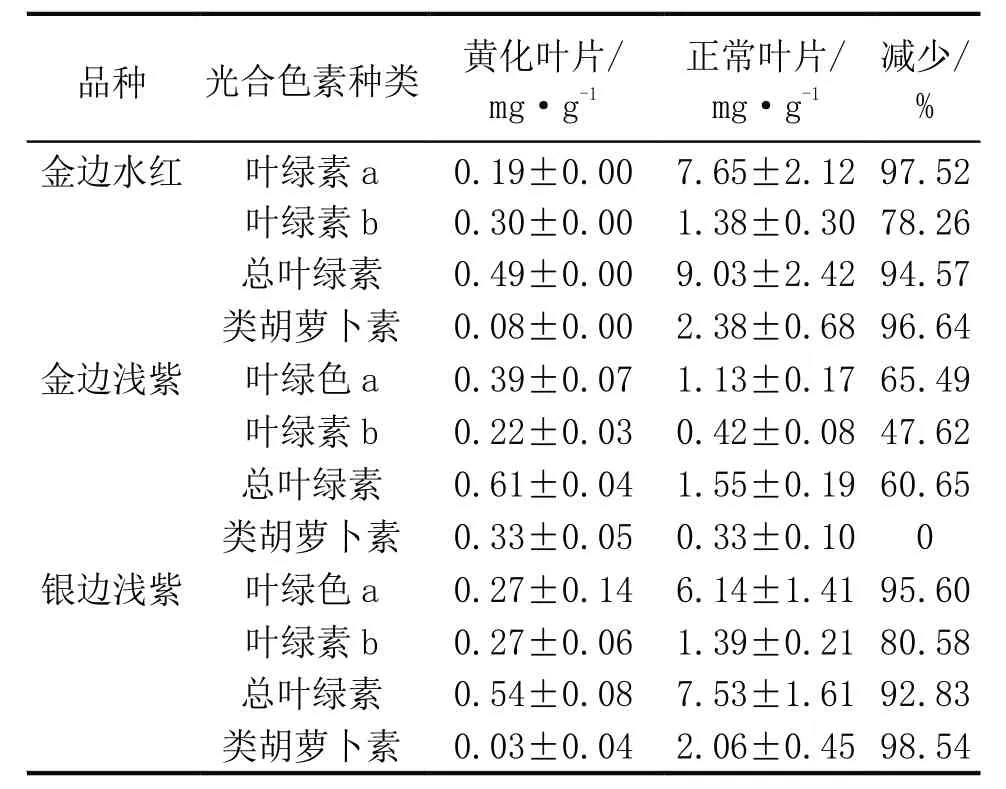
表2 三角梅黄化叶片与正常叶片光合色素含量的比较
2.3 叶绿素荧光参数差异分析
由图2可见,各品种黄化叶片Fo值均低于正常叶片,其中银边浅紫黄化叶片仅为正常叶片的18.92%;金边浅紫为正常叶片的57.97%。
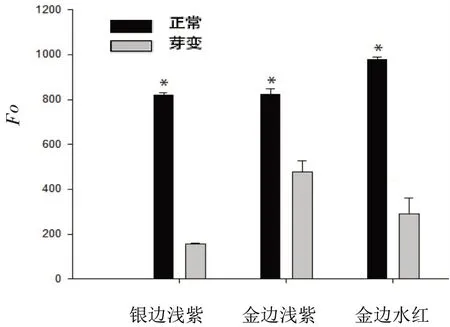
图2 不同三角梅品种黄化叶与正常叶Fo值的比较
图3 显示,3种三角梅黄化叶片Fm均小于正常叶片,其中银边浅紫Fm最小,仅占该品种正常叶片的14.47%;金边浅紫黄化叶片Fm相对较大,占该品种正常叶片的38.04%。
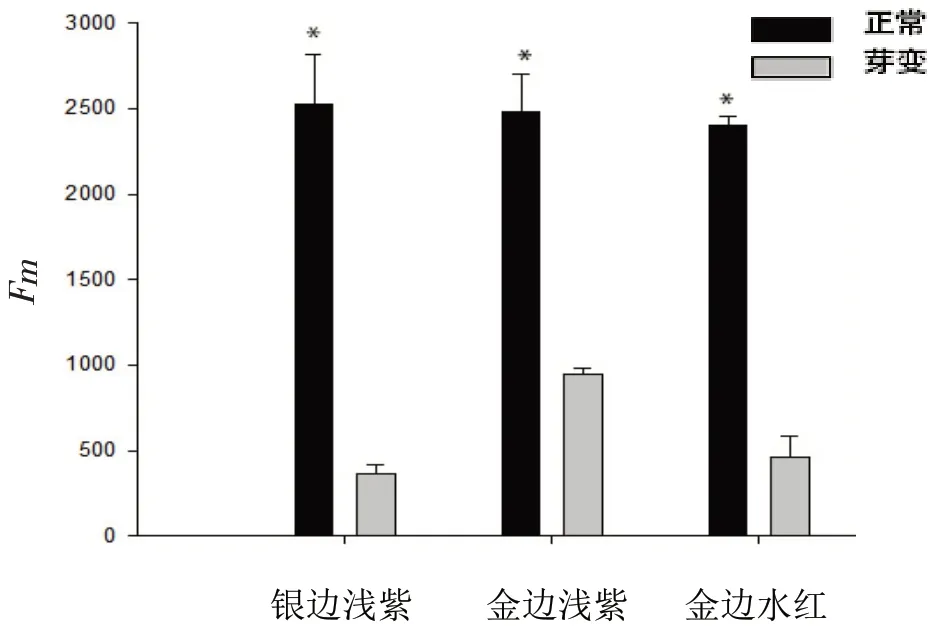
图3 不同三角梅品种黄化叶与正常叶Fm值的比较
银边浅紫芽变品种黄化叶片Fv/Fm比正常叶片低14.93%;金边浅紫低25.76%,金边水红低38.98%(图4)。说明三角梅黄化叶片PSⅡ反应中心可能受到了胁迫或损害。
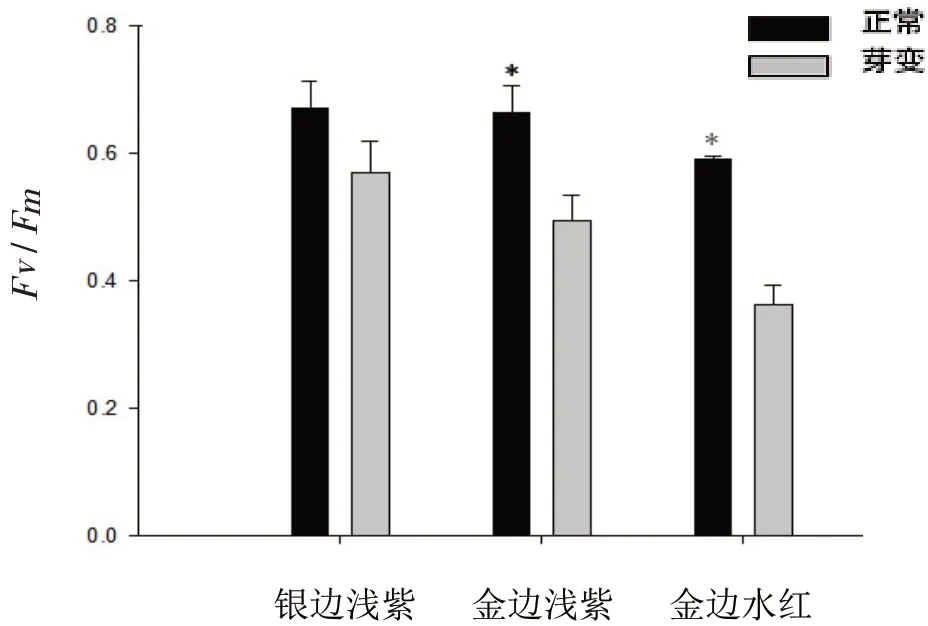
图4 不同三角梅品种黄化叶与正常叶Fv/Fm值的比较
银边浅紫、金边浅紫、金边水红黄化叶片Fv/Fo值均低于正常叶片,分别为正常叶片的65.07%、49.01%、39.46%(图5)。
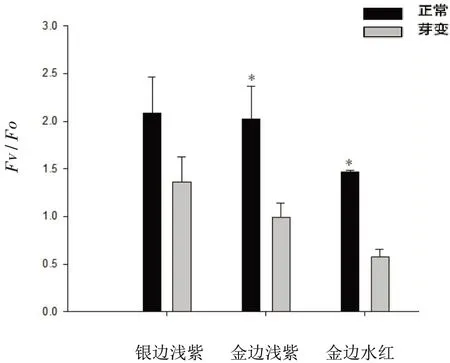
图5 不同三角梅品种黄化叶与正常叶Fv/Fo值的比较
综上可知,3个品种黄化叶片的各叶绿素荧光参数均低于正常叶片,与叶光合色素的测定结果一致。说明黄化叶片光合色素的缺失会同步影响叶绿素荧光参数值,同时,叶绿体膜上的光系统Ⅰ(PSⅠ)光系统Ⅱ(PSⅡ)反应中心可能受到了胁迫或破坏。
3 结论与讨论
植物叶色突变机制复杂,变异种类繁多,有的体现为质量性状,而有的则为数量性状。要更加深入的了解三角梅属植物的叶色突变特性,探究其园艺性状和生理特性就显得十分必要[24]。本研究中,芽变突变体植株上的黄化叶片与正常叶片园艺性状相比较,除了在叶色上有显著区别外,其他性状并无显著差异,说明芽变枝条在不离开母株时,其正常的生长发育不会受到阻碍,母株仍可为其提供足够的营养,至于其脱离母株后能否继续生长则还需进一步试验来加以验证。光合色素具有收集和传递光能的作用,其含量多少能够反映植株的光合性能的强弱。本试验中,三角梅黄化叶片与正常叶片相比,叶绿素a、叶绿素b、类胡萝卜素含量均低于正常叶片,说明叶片颜色黄化的原因与光合色素含量的降低有关,而叶绿素含量的降低可能会导致叶绿体光合性能降低,同时影响芽变突变体的光合速率,此结果与其他学者研究的其他植物黄化结果一致[25-26]。对三角梅来说,黄化叶与白化叶不同,黄化叶是斑叶品种的最高形态,仍然含有少量的叶绿素,而白化叶片则不含叶绿素,若植株全株白化则无法存活[27]。
高等植物活体状态下,叶绿素荧光几乎全部来源于PSⅡ的叶绿素a(Chla)。光合作用的各个过程密切关联,任何一步的变化都会影响到PSⅡ从而引起荧光变化。通过对芽变突变体叶绿素荧光参数的差异分析,三角梅芽变植株黄化叶片叶绿素荧光参数含量均低于正常叶片,说明黄化叶片的光合性能受到了一定影响,同时也能够说明黄化叶片光系统Ⅱ反应中心的蛋白合成受到了一定阻碍。
自然界中叶色突变现象广泛存在,一般发生在植株的顶芽、腋芽部位,原因是该部位生命力最为旺盛,已成熟或老化的部位则很少发生。而三角梅的芽变现象不仅体现在叶色上,还有可能产生苞片的变异,造成一株多色或一朵花多种颜色。对三角梅来说,芽变现象的产生对其新品种的选育有促进作用,若变异性状表现稳定则能提高其观赏价值,同时产生一定的经济效益。本研究结果可为今后开展三角梅叶色突变机理研究等工作提供参考。
猜你喜欢
作物学报(2022年2期)2022-11-06
作物学报(2022年8期)2022-05-29
语文世界(小学版)(2021年5期)2021-06-04
热带作物学报(2020年7期)2020-09-02
山东农业科学(2018年3期)2018-03-29
山东农业科学(2018年11期)2018-01-17
安徽农学通报(2017年18期)2017-10-18
现代农业科技(2017年11期)2017-07-14
热带农业科学(2014年6期)2014-08-07
江苏农业科学(2014年3期)2014-07-16
