
玉树牦牛USP9Y与ND1基因多态性分析及遗传进化研究
2022-01-14 13:28李文浩金夏阳肖立成魏廷虎郭卫兴张雁平
江西农业大学学报 2021年6期
李文浩,金夏阳,任 越,肖立成,魏廷虎,桑 巴,郭卫兴,张雁平*
(1.青海大学 畜牧兽医科学院,青海 西宁 810016;2.西藏自治区农牧科学院,西藏 拉萨 850000;3.青海省玉树市畜牧兽医工作站,青海 玉树 815000)
【研究意义】牦牛遗传资源是青藏高原及其毗邻地区其它畜种资源无可替代的种质资源之一,是我国畜禽遗传资源库中不可或缺的宝贵资源。然而,牦牛受分布地区特殊的自然环境条件以及长期自然选择和人工选育的影响,分布范围较窄,属“区域物种”。因此,和其他家畜(如黄牛、绵羊、猪等)相比,牦牛的研究和开发利用相对比较滞后,至今,仍然是一个以自然选择大于人工选择的牛种[1]。人们对牦牛遗传资源的现状、起源和驯化、分化程度、多样性丰富程度及系统发育关系等问题的研究还不够深入,这极大地限制了牦牛遗传资源的合理保护和开发利用。基于现代分子遗传学的原理和方法,利用测序技术发展带来的机遇,对牦牛遗传资源进行分子评估和系统综合分析,并提出合理的保护和开发利用对策,对促进牦牛业的良性发展和推进民族地区的畜牧业可持续发展具有十分重要的理论和现实意义,有助于牦牛遗传资源的合理保护和开发利用。玉树牦牛产于青海省玉树藏族自治州,属青藏高原腹地的三江源头,平均海拔4 200 m 以上,是“世界屋脊”青藏高原的主要组成部分,也是世界牦牛的主产区之一,不仅具有牦牛产业发展得天独厚的自然资源,还拥有中国数量最多、品质最好的牦牛资源,数量占30%以上,位居全国第一[2-5]。【前人研究进展】近年来,众多学者基于USP9Y基因(Ubiquitin-Specific Protease 9 gene on the Y)在不同动物父系遗传多样性、起源和进化研究中进行了大量研究[6-10]。而对于牦牛USP9Y基因的研究中,Ma 等[11]使用5 种Y-SNP 标记(USP9Y,SRY,UTY,AMELY和OFD1Y)对9 个品种的322 头牦牛进行了Y 染色体单倍型多样性及群体遗传结构研究,表明青海牦牛具有2 个不同的父系遗传谱系,个体间存在显著差异,青海牦牛群体的遗传结构较弱。Y 单倍型总体多样性为0.538±0.028,表明青海牦牛具有较高的父系遗传多样性。Li 等[12]根据USP9Y、UTY19和SRY4等标记对65 头公牦牛Y 染色体多态性进行分析,发现牦牛拥有2 个高度分化的父系支系(即YH1和YH2 vs.YH3)。Ma 等[13]基于USP9Y,SRY4,UTY19,AMELY3,OFD1Y10和INRA189Y染色体特异性标记对8 头公野牦牛进行分析,结果显示野牦牛具有丰富的父系遗传多样性,其Y 染色体单倍型多样性(Hd)为0.821±0.101,野牦牛拥有2 个父系遗传分支。而对于牦牛ND1基因的研究仅见于Shi等[14]对2 个高低海拔的牛属群体(70 头西藏牦牛和70 头宣汉牛)的ND1基因进行测序分析,结果表明MT-ND1和MT-ND2被认为是适应高海拔环境的候选基因。【本研究切入点】综上所述,目前基于USP9Y和ND1基因已在牦牛群体内部进行不同方面的研究[6-14],但是尚未见基于2 种标记结合对玉树牦牛群体的多态性研究。鉴于此,本研究以分布在青海玉树地区的牦牛群体为研究对象,选择USP9Y和ND1基因并对其PCR 扩增产物直接测序检测多态位点,探讨USP9Y和ND1基因在玉树牦牛群体中的多态性。【拟解决的关键问题】对玉树牦牛内部的遗传信息基于育种建议,为进一步对牦牛保护性开发利用牦牛遗传资源及选育新品种提供理论科学依据。
1 材料与方法
1.1 样本采集与基因组DNA提取
随机采集青海省玉树市100 头牦牛(公牦牛40 份、母牦牛60 份)血样带回实验室,采用“两步法”从FTA 卡中提取基因组DNA,提取方法参照Zhou 等[15],后对基因组DNA 进行质量与浓度检测,合格的基因组DNA保存于-20 ℃备用。
1.2 引物合成与PCR扩增
参照牦牛USP9Y、ND1基因编码序列(GenBank NM_001145509、NC_006380),利用primer3.0在线软件设计引物。引物信息见表1。引物由西安杨凌天润奥科生物科技有限公司合成。使用时先将引物干粉稀释成100 pmol/μL作为原液,再取出10 μL引物原液加入90 μL dd H2O,工作液的最终浓度为10 pmol/μL。

表1 引物信息Tab.1 Primer information
PCR 反应体系为20 μL。反应体系如下:dd H2O 7.6 μL,TaqMaster Mix 10.00 μL,上下游引物各0.8 μL(10 μmol/L),基因组DNA 0.8 μL。反应条件为:94 ℃预变性5 min,94 ℃变性30 s,57°C 退火30 s(USP9Y引物)或62 ℃退火30 s(ND1引物),72 ℃延伸30 s,共35 个循环,72 ℃延伸10 min 后4 ℃保存。PCR 扩增产物用1.5%琼脂糖凝胶电泳检测。
1.3 基因分析及SNP突变位点检测
将40份公牦牛、60份母牦牛PCR 扩增产物(USP9Y和ND1)送至天津金唯智测序公司进行测序。应用MEGA7 软件比对等位基因序列,并下载青海牦牛相应序列进行亲缘关系的构建,探明遗传距离并揭示牛种内序列遗传变异。
2 结果与分析
2.1 牦牛USP9Y和ND1基因PCR扩增产物琼脂糖电泳检测
PCR 扩增产物用1.5%的琼脂糖凝胶电泳检测,紫外灯照射下结果均和目的片段大小相一致,条带单一且清晰,表明PCR扩增获得目标片段,可以进行测序。
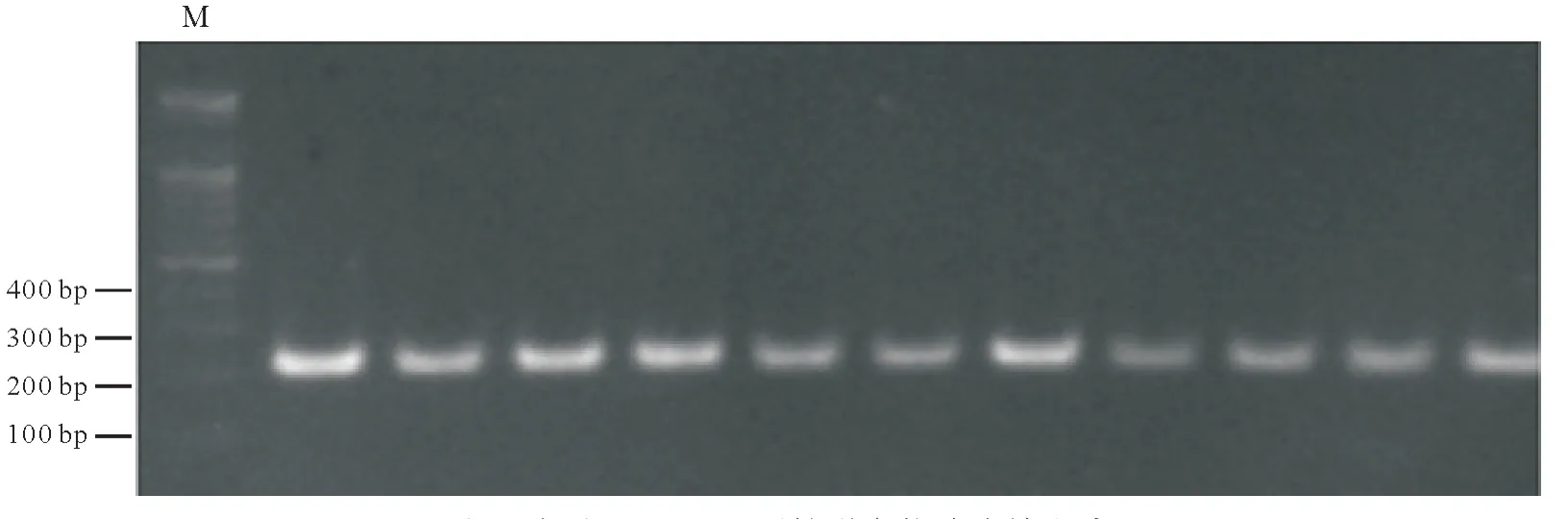
图1 牦牛USP9Y基因扩增产物琼脂糖电泳Fig.1 Agarose electrophoresis of amplification products of yak USP9Y gene
2.2 USP9Y基因SNP位点的筛查与检测
将牦牛USP9Y扩增后,PCR 扩增产物送天津金维智生物技术有限公司进行正反双向测序,运用Chromas 2.3对扩增序列的测序结果与NCBI上参考目的片段序列进行比对并校正拼接,得出序列长度为210 bp;使用MEGA7.0 软件打开测序结果图谱寻找双峰位点(突变位点)。结果发现在40 份公玉树牦牛中均未发生碱基突变。

图2 牦牛ND1基因扩增产物琼脂糖电泳Fig.2 Agarose electrophoresis of amplified products of yak ND1 gene
2.3 ND1基因SNP位点的筛查与检测
对玉树牦牛ND1基因扩增后,运用Chromas 2.3 对扩增序列的测序结果与NCBI 上参考目的片段序列进行比对并校正拼接,得出序列长度为220 bp,并使用MEGA7.0 软件打开测序结果图谱寻找双峰位点(突变位点)。结果发现玉树牦牛中均未发生碱基突变。
2.4 青海牦牛部分USP9Y基因多态位点分析
根据马志杰[16]使用USP9Y基因标记对中国的682 头公牦牛(青海、甘肃、云南、新疆和西藏)以及8 头野牦牛公牛进行多态性分析,得出在470 bp 的USP9Y基因中仅有1 个多态位点(g.249C>T)(图3),本实验扩增的目的片段大小为210 bp,未检测到该多态位点,表明玉树牦牛USP9Y 序列前210 bp 处较为保守,无突变位点。
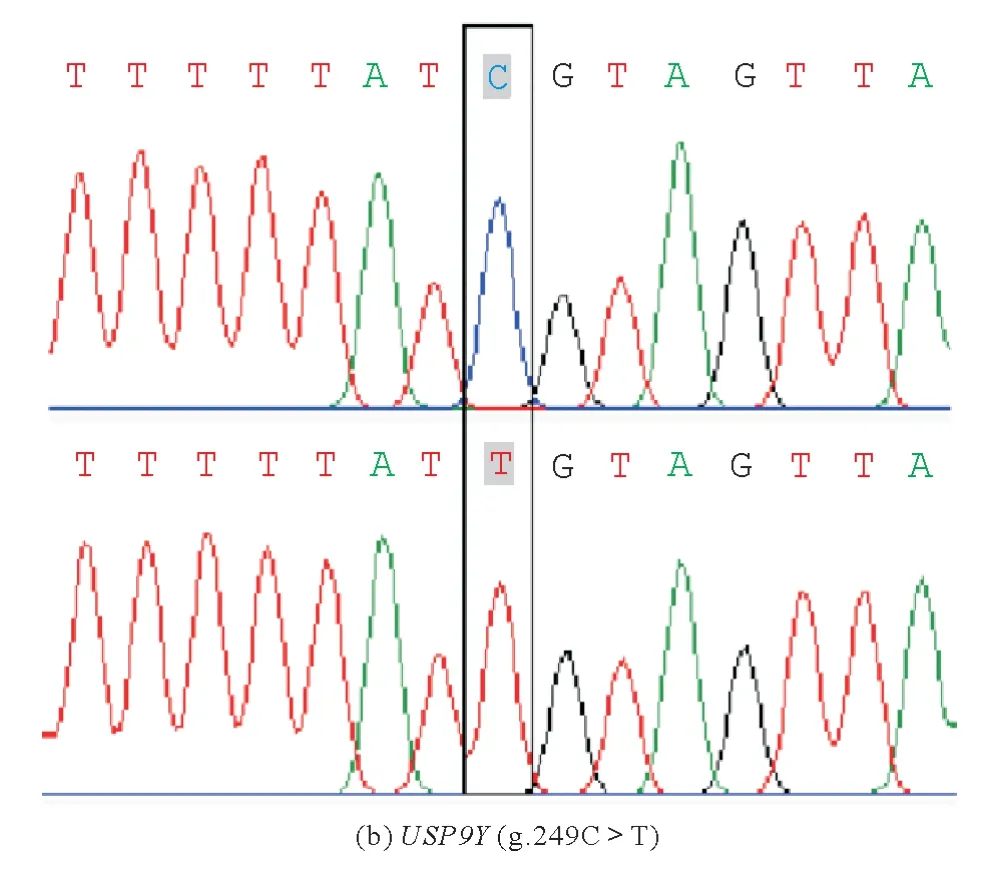
图3 牦牛USP9Y基因多态位点Fig.3 Polymorphic loci of USP9Y gene in yak
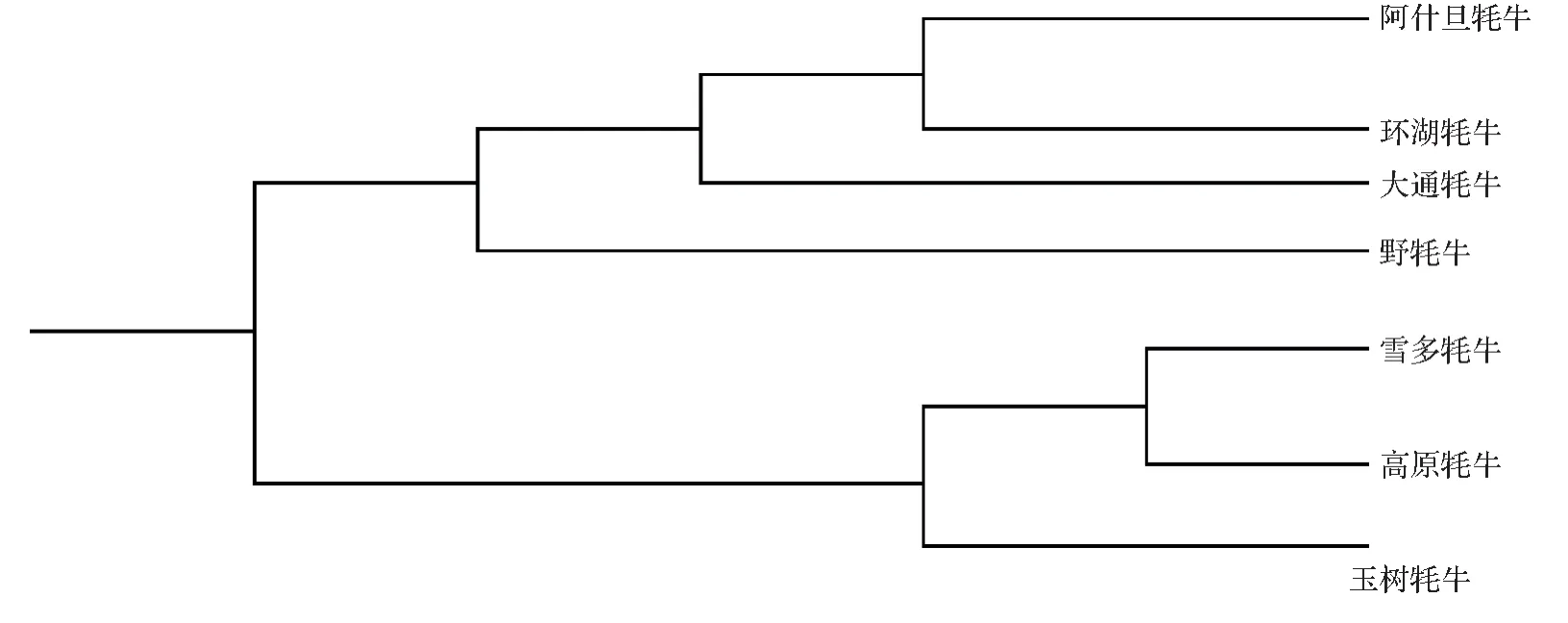
图4 基于ND1基因构建玉树牦牛与其他青海牦牛的遗传关系Fig.4 The genetic relationship between Yushu yak and other Qinghai yaks was constructed based on ND1 gene
2.5 玉树牦牛与其他牦牛部分ND1基因亲缘关系分析
通过下载青海范围内牦牛ND1基因序列(高原、雪多、环湖、阿什旦、大通和野牦牛)并与本实验中的玉树牦牛的ND1序列进行构树,并探明其亲缘关系,实验结果显示,玉树牦牛与高原和雪多牦牛距离较近,而与阿什旦、环湖、大通和野牦牛遗传距离较远,故建议玉树牦牛与环湖牦牛种公牛进行适当的杂交,提高玉树牦牛的遗传性能。
3 结论与讨论
牦牛毛长皮厚,体矮身健,能够适应特殊的高原环境,包括缺氧、高寒和低气压,被称为我国的高原之舟。长期的地域分布局限使牦牛的遗传背景较为清晰,是探明高海拔低氧适应性很好的模式动物。Qiu 等[17]在2012 年组装出第一个牦牛的参考基因组,且分析得出牦牛和普通牛约在490 万年前就已分化,在牦牛基因组中共鉴定出85 个正向选择基因(positive selected genes,PSGs)均与缺氧应激和能量代谢有关,包括2 个重要的调控子(Adam17和Arg2)和1 个靶基因(Mmp3)与缺氧功能有关。Ding 等[18]通过采用反转录定量聚合酶链反应和western-blot 方法检测牛和牦牛脑组织中AQP4mRNA 和蛋白的表达是否存在差异,此外也利用免疫组化技术分析了AQP4在牦牛和牛脑中的定位和表达情况。AQP4可能通过牦牛大脑低表达和维持正常生理功能在抵抗脑水肿中发挥重要作用,对高海拔与缺氧应激有着积极作用。在对牦牛组织形态学对于高海拔与缺氧方面也有学者进行研究,Xiong 等[19]克隆了牦牛中HIF-1和HIF-2完整编码区,确定其在几个组织中的mRNA 表达,并与亲缘关系密切的低海拔牛的表达水平进行比较,研究结果表明在牦牛中HIF-1和HIF-2的表达量明显高于普通牛,除肺外,牦牛各器官中HIF-1的表达量均显著高于普通牛(P<0.05),但心脏、脾脏和肾脏中HIF-2脾的表达量显著高于牛(P<0.05)。
遗传多样性研究可以评估动物遗传变异的程度大小,进而反映动物遗传多样性水平的高低,从而为动物遗传资源的保护和选育策略制定提供科学依据。在以往对牦牛USP9Y基因多态性中,Ma 等[11]在牦牛USP9Y基因中仅发现1处多态位点(即g.249 C>T),而本研究中玉树牦牛群体中均未发现多态位点;对于ND1基因,Shi 等[14]研究结果为在所有的西藏牦牛中均有MT-ND1(SNP m.3,907 C>T),与高原适应呈正相关(P<0,006),而在本研究中玉树牦牛群体中也未发现多态位点,究其原因,可能与采样点与所取的片段长度有关,或是由于玉树牦牛所处的位置较为封闭从而使其内部的核苷酸多态性低,与外界牦牛的基因交流较少导致的,Ma等[11]认为青海牦牛具有较为丰富的父系遗传信息,但是本研究中显示玉树牦牛作为青海牦牛的代表品种,其USP9Y基因多态性低,从而导致其遗传多样性低。其他作者对牦牛母系遗传研究结果认为,我国牦牛遗传多样性丰富,有2~3 个母系起源[20-27]。但是本研究得出ND1作为线粒体基因组上的一段区域,在玉树牦牛群体内未检测到多态位点,表明该区域不适于做群体遗传多态性研究,而后续的研究应当多基于Cyt-b、D-loop 或是线粒体基因组来评估群体的母系遗传多样性。根据马志杰[16]使用USP9Y基因标记对中国的682头公牦牛和8头野牦牛公牛进行牦牛的父系遗传多样性分析,得出在扩增的470 bp的USP9Y基因中仅有1个多态位点(g.249 C>T),本研究中,由于扩增片段的长度受限,未扩增出该多态位点。由于USP9Y是一个较为保守的基因序列,故今后的研究中应当调整引物,增加扩增长度。
牦牛种质资源可持续利用对于高海拔地区的经济是必不可少的,已有不少研究者应对不同地区不同品种的牦牛遗传资源提出了相应的育种建议[16,28],目前青海公认的牦牛品种有4 个,分别为高原牦牛、玉树牦牛、环湖牦牛和雪多牦牛。本研究基于母系ND1基因与USP9Y基因对玉树牦牛品种进行多态性检测时,发现该品种内部父系与母系遗传结构较为单一,原因可能与该品种生活在海拔较高的地区,生存环境较为封闭,导致其内部的父本较为单一,本研究结果显示,玉树牦牛与高原和雪多牦牛距离较近,而与阿什旦、环湖、大通和野牦牛遗传距离较远,鉴于此,建议应当适度的调用环湖种公牦牛或其他省份的种公牦牛对玉树牦牛进行杂种优势的选育,使其充分的发挥其应有的遗传潜力。
猜你喜欢
青海湖(2022年3期)2022-06-09
滇池(2022年5期)2022-04-30
散文诗(2021年22期)2022-01-12
小哥白尼(野生动物)(2020年9期)2021-01-18
新世纪智能(高一语文)(2020年12期)2020-06-01
小小说月刊(2019年20期)2019-10-19
科技视界(2016年27期)2017-03-14
中国医药导报(2015年27期)2015-02-28
火花(2015年1期)2015-02-27
中国神经精神疾病杂志(2013年1期)2013-03-11
