
油茶NAC基因鉴定及对干旱胁迫响应分析
2022-01-14 13:28曹瑞兰李知青欧阳雯婷胡冬南周增亮苏文娟
江西农业大学学报 2021年6期
曹瑞兰,李知青,欧阳雯婷,胡冬南,周增亮,苏文娟,陈 霞,刘 娟*
(1.江西农业大学 林学院/江西省森林培育重点实验室,江西 南昌 330045;2.福建农林大学 林学院,福建 福州 350028)
【研究意义】油茶(Camellia oleiferaAbel.),是我国特有的木本油料树种,与油橄榄、油棕和椰子并称为世界四大木本油料植物,综合利用价值极高[1-2]。油茶生长在山地或丘陵地带,适生性强,耐干旱贫瘠,且不与粮棉争耕地,近年来已成为我国南方山区重点发展的油料作物。发展油茶产业也成为保障国家粮油安全、提高食用油自给率的重要途径。截至2019年底,我国油茶种植面积约453.3万hm2,其中高产油茶林93.3万hm2,产油62.7万t,全国油茶产业总产值达到1 160亿元[3]。
干旱是影响植物生长发育的主要逆境胁迫之一,也是全球作物栽培面临的最严重的环境压力之一[4]。油茶是我国南方山地的乡土树种,一般认为其耐旱性较强,但是其主产区在6—9 月常出现持续干旱,同时伴随着极端高温,这将严重影响油茶的集约化栽培和生产[5]。干旱胁迫将降低油茶光合效率,影响花芽分化和油脂转化,导致大量落花落果,进而造成油茶林大面积减产[6]。民间谚语“七月干果,八月干油”,说的就是夏季高温干旱对油茶产量和茶油品质的影响。大量研究发现,在长期进化过程中,油茶已形成一系列形态、生理机制来应对或适应干旱胁迫,例如,通过促进根系生长,提高根冠比[7];关闭部分叶片气孔,破坏光合色素,降低光合效率,减缓植株生长[8];减小花芽分化,降低油脂转化,抑制生殖生长等[9]。在分子调控水平上,诱导或抑制一系列干旱胁迫相关的基因表达,这包括直接参与干旱胁迫相关基因,以及调节这些基因表达水平的转录因子。【前人研究进展】NAC 转录因子家族是植物界数量最大、分布最广的转录因子家族之一,主要参与调控植物的生长发育和形态建成[10-12]。Souer[13]和Aida等[14]先后从矮牵牛和拟南芥中发现了NAM、ATAF1/2 和CUC2 等3 个NAC 转录因子。序列比对分析结果发现,NAM、ATAF1/2 和CUC2 所编码的蛋白质具有较高的同源性。即使它们有不同的生物学功能,但其N 端结构相似,故以其首字母命名为NAC[15]。研究发现NAC 家族NAM和CUC基因对顶端分生组织的形成和分化具有重要调控作用[16],启动子区富含逆境响应元件,如低温响应元件[17]、干旱响应元件[18]、损伤响应元件[19]等,是植物应答生物或非生物胁迫的重要方式之一。
【本研究切入点】研究表明,NAC 转录因子在植物应对干旱的过程中具有重要的调节作用。例如,Hu等[20]发现,NAC转录因子能在气孔保卫细胞中被诱导表达,调控叶片气孔运动,进而提高植株抗旱性。Duan 等[21]研究表明,苜蓿在受到干旱胁迫时,MfNACsa去棕榈酰化易位至细胞核,通过调节相关基因表达维持植株体内谷胱甘肽库处于还原状态,保证植株在干旱胁迫下正常生长发育。拟南芥中转录因子ANAC096与ABRE结合蛋白共同激活植株体内ABA诱导基因,促使植株体内ABA合成,保证在干旱时植株体内渗透压平衡,增加抗旱性[22]。【拟解决的关键问题】本研究从课题组油茶三代全长转录组数据分离得到CoNAC转录因子家族成员,明确各个基因的系统发育进化关系、理化性质、Motif 预测、亚细胞定位等,并基于油茶干旱转录组数据,分析CoNAC在不同干旱胁迫程度和复水下的表达水平,最后采用q-PCR 验证CoNAC家族成员在不同耐旱油茶品种和不同干旱胁迫强度下的表达特征,进而探讨油茶响应干旱胁迫的分子调控机理,为水分高效利用油茶基因型的筛选提供了理论基础和科学依据,具有重要的现实意义。
1 材料与方法
1.1 材料
取生长较一致的一年生油茶苗为试验材料,供试品种选取课题组前期干旱胁迫研究中筛选的相对耐旱油茶品种长林53 和相对不耐旱的长林18。2019 年7—9 月在江西农业大学科技园大棚内进行盆栽试验,每3株一盆,每盆装相等的质量的河沙为基质,测定土壤田间持水量和含水量。试验设置充分供水(对照)、自然干旱(8 d)、复水(自然干旱8 d后)3个处理。采用称量控水法于每天17:00称量测定土壤含水量,并将同一处理补水至相同含水量值。实验对照组的土壤含水量维持在田间持水量的85%~90%;处理组土壤经自然干旱,第8 天时土壤含水量下降至田间持水量的35%,处理第9 天复水至对照水平。分别在处理前、处理第8 天、复水后第5 天采样,采样时间为每天08:00—09:00。采集油茶苗幼叶,立即放入液氮中冷冻,然后置于-80 ℃冰箱冷藏,直至RNA提取。
1.2 方法
1.2.1 油茶NAC 基因家族成员的筛选及基本信息分析 下载拟南芥NAC 蛋白序列(https://www.arabidopsis.org/)和茶树NAC蛋白序列[23],使用本地blast-2.9.0+程序中tblastn对课题组已有的油茶三代测序转录组数据进行检索,参数设置为-evalue 1e-5,-max_target_seqs 100,-outfmt 6。随后,将初始搜索中鉴定出的所有油茶NAC基因序列上传至NCBI 数据库中的CDD(http://www.ncbi.nih.gov/structure/cdd/wrpsb.cgo)进行在线验证,确定存在NAC结构域,去除冗余和无效基因模型。将CDD验证的所有候选基因上传到ORFfinder(http://www.ncbi.nih.gov/orffinder/),获得CDS 序列和氨基酸序列。最后将这些氨基酸序列再次使用CDD 和Pfam 验证,得到油茶NAC基因家族候选基因。ProtParam(http://web.expasy.org/protparam)在线分析分子量(MW)、理论等电点(pI)等基因的一级结构,利用CELLO v.2.5(http://cello.life.nctu.edu.tw/)进行亚细胞定位预测。
1.2.2 多序列比对及系统进化树构建 利用ClustalX2.1软件将本研究鉴定出的油茶CoNAC蛋白序列同拟南芥(A.thaliana)和茶树(C.sinensis)的NAC 蛋白序列进行多重比对分析,然后运用MEGA10.0软件打开,在氨基酸序列中使用邻近法(Neighbor-Joining method)构建系统进化树,参数设置为泊松矫正(Poission correction)、成对删除(pairwise deletion)和1 000 次重复的Bootstrap 值。使用在线程序iTOL(https://itol.embl.de/)对进化树进行美化。根据序列比对结果和进化树,通过引进已知亚家族分类的拟南芥NAC蛋白和茶树NAC蛋白的NAC区域,分类方法综合Hisiko[24]文献,对油茶NAC基因进行亚家族的分类。
1.2.3 基因家族保守结构域分析 利用保守元件分析工具MEME 5.0.5(http://meme-suite.org/tools/meme)在线对油茶NAC 家族蛋白进行保守序列分析,设定参数motif 的最大数目为20,motif 长度设置为6~60个氨基酸,其他则为默认值。使用TBtools软件绘图。
1.2.4 基因表达谱分析 根据已发表的油茶干旱转录组数据,使用Galaxy 生信在线分析平台的Salmon quant 工具进行转录组reads 的mapping,将不同处理的所有可用读取数据分别映射到对应的转录组数据库,计算油茶NAC基因的TPM值,最后使用TBtools绘制表达图谱。
1.2.5 候选基因的表达模式分析 提取油茶长林18 和长林53 幼叶在不同试验处理下的RNA,使用iScriptTMcDNA Synthesis Kit 进行cDNA 的合成,于-20 ℃保存,用于差异表达分析。挑选CoNAC28、CoNAC51、CoNAC52、CoNAC56、CoNAC60和CoNAC61基因进行q-PCR 验证,引物序列如表1所示,内参基因为EF1a1。将所有样品都放在冰盒,按照所需的反应体系进行实验,每个样品设置3个重复。设置好荧光定量仪程序,待反应结束后,采用2—△△CT方法进行计算实时荧光定量反应中候选基因的相对表达量。

表1 实时荧光定量PCR引物Tab.1 Primers for quantitative real-time PCR
2 结果与分析
2.1 油茶NAC家族的筛选与基本信息分析
本研究在已成功构建的RNA-seq 本地blast 文库的基础上进行NAC基因相似序列检索与搜索,经CCD 和Pfam 在线数据库进一步筛选分析后最终确定了67 个含有NAM 保守结构域的序列。根据转录组数据库中unigenes 对67 个NAC 基因进行编号,即CoNAC1~CoNAC67(表2)。油茶NAC 蛋白质理化性质分析显示,67 个CoNAC基因大小为423(CoNAC48)~1911(CoNAC7)bp,编码的氨基酸长度在140(CoNAC48)~636(CoNAC7),这些油茶NAC 蛋白的氨基酸长度变化差异较大;相对分子质量介于15.65(CoNAC48)~388.73(CoNAC13)ku,与氨基酸含量成正比;等电点分布在4.54(CoNAC11)~9.65(CoNAC65),其中26 个为碱性蛋白质(pI>7),41 个为酸性蛋白质(pI<7);有18 个CoNAC 的不稳定指数均小于40,是稳定蛋白,约占NAC基因家族的26.87%。CoNAC成员的总平均亲水系数(GRAVY值)都小于0。因此,推断这些油茶NAC 家族蛋白表现均为亲水蛋白,并且多数稳定。亚细胞定位预测表明,除了CoNAC24 定位在叶绿体和细胞核,其余的油茶CoNAC蛋白都定位在细胞核。

表2 油茶NAC蛋白基本信息Tab.2 Basic information of C.oleifera NAC proteins
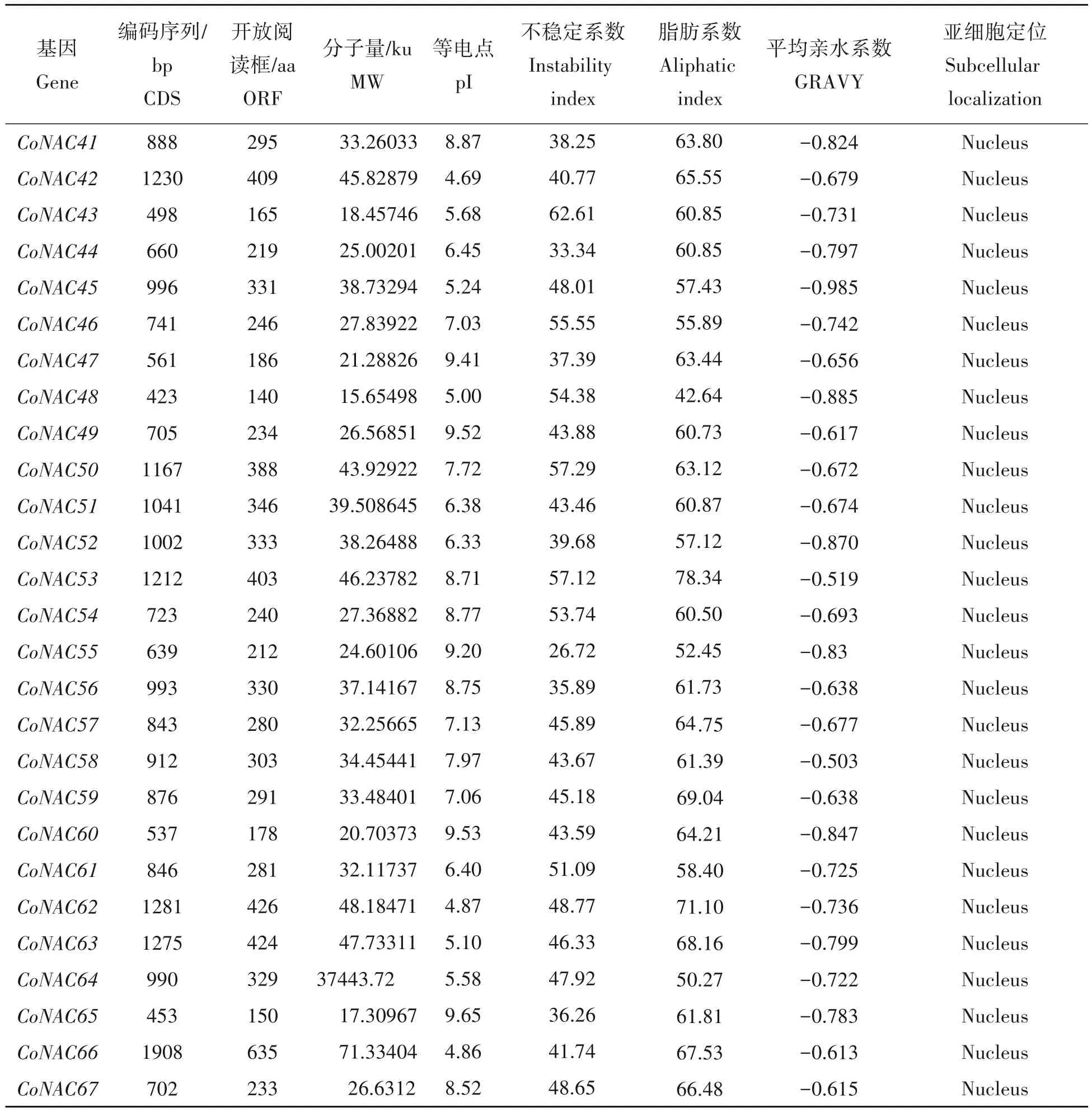
续表Continued tab.
2.2 油茶NAC蛋白保守结构域分析
保守结构域是蛋白质中能够折叠成特定三维结构的一段相对区域,其结构亚单位称为基序(Motif),组成这些基序的氨基酸区段行使特异的功能,并蕴含各自的遗传进化信息[26]。通过GeneDoc软件对油茶NAC转录因子进行序列比对(图1),发现油茶NAC转录因子的蛋白质的N端存高度保守的NAC结构域(约150~160个氨基酸),这与拟南芥也相吻合[24,27]。此外,油茶NAC家族成员中除了少数基因具有不完整的NAC结构域外,大多数基因具有5个保守的亚结构域(A~E),说明CoNAC基因在进化过程中具有高度保守性。

图1 油茶NAC蛋白序列比对Fig.1 Sequence alignment of C.oleifera NAC proteins
2.3 油茶NAC蛋白Motif分析
CoNAC基因家族中选用了20个Motif对油茶的NAC蛋白分析显示(图2),油茶NAC家族单独建树与拟南芥、茶树和油茶共同构建的物种间系统进化树总体趋势相吻合(图3),表明通过建树预测基因功能稳定可靠。按照Motif 在CoNAC基因家族中出现次数进行排序,其中Motif 5 出现65 次,Motif 6 出现62 次,Motif 1 出现58 次,Motif 2 和Motif 4 出现56 次,Motif 7 和Motif 10 出现12 次,Motif 12 和Motif 13 出现7次,Motif 8和Motif 9出现6次,Motif 11、Motif 14、Motif 19和Motif 20出现5次,Motif 15、Motif 16、Motif 17和Motif 18 出现4 次,Motif 在CoNAC基因家族中出现频率越高说明其越重要。通过分析Motif 与蛋白结构域,发现Motif 5、6、1、2、3和4分别对应CoNAC转录因子N端5个保守的亚结构A~E。

图2 油茶中NAC蛋白保守基序Fig.2 Sequence logos of CoNAC domains in C.oleifera
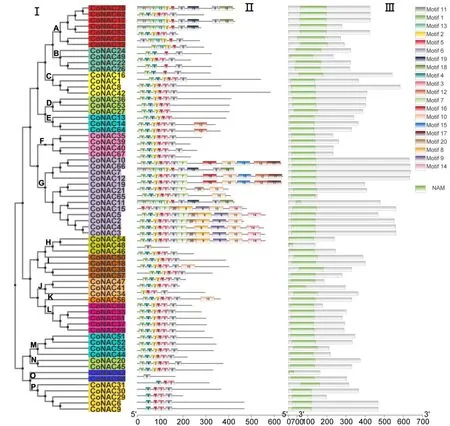
图3 NAC基因家族成员保守基序分析Fig.3 Distribution of conserved Motifs in the NAC gene family members
2.4 NAC基因家族的多序列比对和系统进化树构建
为充分明确油茶NAC基因家族各基因间亲缘关系与生物学功能的关系,对鉴定得到的油茶、茶树和拟南芥的NAC 基因家族成员进行进化关系分析,根据Hisako 等[24]的分类方法,对CoNAC家族进行了分类,结果显示(图4):进化树共分为17 个亚组,其中ANAC2 最多有13 个成员,ONAC003 次之有8 个CoNAC成员,ATAF 和ANAC63 含有5个CoNAC成员,NAM、Unclassified、TIP 和SENU5含有4 个CoNAC成员,NAP、TERN、ONAC22、ANAC011和OSNAC8含有3个CoNAC成员,OSNAC7、NAC1含有2个CoNAC成员,ATNAC3和ONAC022亚组最少,含有1个CoNAC成员,只有ANAC001亚组中不包含CoNAC基因。在进化关系上比较相近的一类NAC 基因在结构与功能上均比较相似,可以作为推测油茶中NAC 基因功能的参考依据。

图4 油茶、茶树和拟南芥NAC的邻接法进化树Fig.4 Neighbor-joining phylogenetic tree of NAC in C.oleifera,C.sinensis and A.thaliana
2.5 油茶NAC基因家族表达分析
基于油茶不同干旱胁迫处理下转录组数据,使用TBtools 分析软件进行Row-Scale 均一化处理(图5),发现在中度干旱、重度干旱和复水后,大部分CoNAC基因间的表达模式存在一定差异。其中,CoNAC1、CoNAC8、CoNAC9、CoNAC27、CoNAC34、CoNAC43、CoNAC45、CoNAC47和CoNAC58等基因在干旱胁迫下,下调表达;CoNAC15、CoNAC16、CoNAC18、CoNAC28、CoNAC49、CoNAC51、CoNAC52、CoNAC55、CoNAC56、CoNAC57、CoNAC60、CoNAC61、CoNAC65和CoNAC67等基因上调表达,其中CoNAC56在中度干旱胁迫下的表达量比对照增加7 倍,CoNAC60基因的表达量增加4 倍,CoNAC61增加12 倍,达到最高峰。随着干旱胁迫程度加强,CoNAC15、CoNAC16、CoNAC18、CoNAC28、CoNAC51、CoNAC60、CoNAC61等基因表达量也持续增加。在复水后,CoNAC18、CoNAC28、CoNAC49、CoNAC51、CoNAC52、CoNAC56、CoNAC57、CoNAC59、CoNAC60、CoNAC61和CoNAC65等基因表达呈明显下降趋势,而CoNAC12、CoNAC14、CoNAC23、CoNAC30、CoNAC36、CoNAC41、CoNAC50、CoNAC53、CoNAC54、CoNAC62、CoNAC63和CoNAC64等基因表现为上调表达,表达量增加。分析油茶干旱转录组数据发现,CoNAC基因家族中有大部分CoNAC会响应干旱胁迫。为了验证RNA-seq数据,本研究挑选了CoNAC28、CoNAC51、CoNAC52、CoNAC56、CoNAC60和CoNAC61基因进行q-PCR验证。
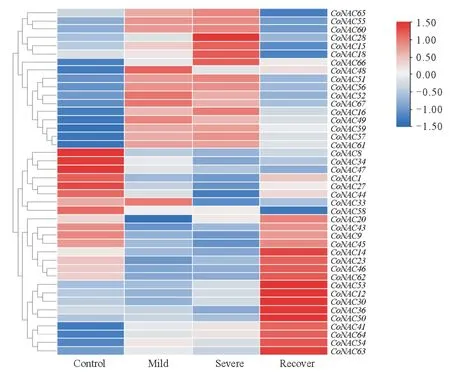
图5 CoNAC基因在不同干旱胁迫下的表达特征Fig.5 Expression profiles of CoNAC genes under different concentration of drought
2.6 候选基因在干旱胁迫下的表达模式分析
为了研究CoNAC基因在油茶不同干旱时期的表达情况,本研究挑选在干旱胁迫表达谱中表达量较高的6 个CoNAC基因,利用q-PCR 检测它们在2 个不同耐旱油茶品种(长林18 和长林53)中处于适宜水分、干旱、复水试验下叶片中表达量。结果显示,在干旱胁迫和复水后,大部分CoNAC基因在2个油茶品种中均表现出较为相似的表达趋势,即基因在干旱处理时表达量均呈显著上调趋势,复水后呈现下降趋势,这与表达谱(图5)结果基本相同。例如,在长林53和长林18中,在干旱处理时CoNAC28、51、56、61等4个CoNAC基因候选表达量在干旱胁迫处于最高峰,复水后表达量有所下降。
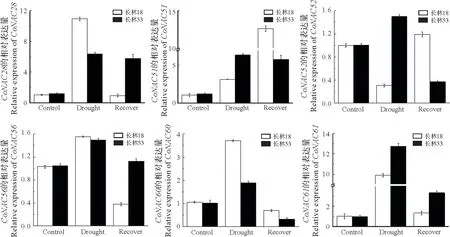
图6 6个CoNAC基因表达特征Fig.6 Expression patterns of 6 CoNAC genes
同时,笔者也发现CoNAC51和CoNAC52基因表达模式呈现不同趋势,而且在不同品种间的表达模式不同。例如,在长林18 中,CoNAC52基因在干旱胁迫时表现为下调表达,复水后显著上调,而CoNAC51基因在干旱胁迫下表达量呈上升趋势,复水后表达量持续增加。长林53中,这2个基因的表达模式均呈现随着干旱胁迫增加而上调,复水后下调。
3 讨论
油茶是我国南方山地或丘陵地区主要的经济树种之一。虽然油茶属于中等耐旱植物,但是干旱胁迫仍是其生长发育过程中常遇到的逆境胁迫。研究发现,南方地区夏季持续干旱会对油茶生长产生严重影响,进而造成油茶林大面积减产,油茶果的果径、果高、果形指数和体积等降低,花径的生长量较自然状况减少[5]。NAC 家族的NAM和CUC基因具有高度保守的结构域,能调控顶端分生组织的形成和分化,且启动子区富含逆境响应元件,在植物应答干旱胁迫中具有重要作用[29-30]。本研究从油茶全长转录组数据中鉴定出67 个NAC 转录因子。油茶为六倍体树种,其NAC基因数目明显低于拟南芥(117)[31]、水稻(151)[32]、大豆(152)[33]、茶树(45)[23]等二倍体物种。这可能是在三代转录组测序过程中由于受到基因文库构建或测序仪数据读取不确定性的影响,导致相对基因组测序来说存在基因数目不全等问题。因此油茶NAC家族成员的完整性需要依赖全基因组测序数据来进一步确定和完善。
根据油茶、拟南芥和茶树NAC 家族共建的系统发育树,油茶67 个NAC基因家族的蛋白序列可分为17个亚族,除AtNAC001亚组中不包含油茶CoNAC基因,其它每个亚族都同时含有拟南芥和茶树NAC基因。其中ANAC2 和ONAC003 亚组成员较多,暗示ANAC2 和ONAC003 亚组对油茶具有更加重要的作用。一般而言,位于同一系统发育进化枝小组上的蛋白,若序列的相似性较高,则它们可能具有极为类似的蛋白功能[34]。因此,可以利用NAC 蛋白间的进化保守性,来探讨和预测油茶NAC 蛋白的抗旱能力。已有研究表明,ATAF、AtNAC3、ONAC003、NAM 亚组在非生物胁迫中具有重要作用[23,35]。拟南芥水胁迫处理抑制ATAF1的表达,脱水和ABA 处理可以诱导拟南芥ATAF 亚组的一些干旱相关基因表达,从而影响拟南芥抵抗干旱胁迫[12,36]。在本研究中,系统发育结果分析显示,油茶CoNAC60和CoNAC61分布于ATAF亚类,与拟南芥AtNAC102、AtNAC81、AtNAC32、AtNAC2聚为一类,并且该基因在干旱胁迫后表达量上调,表明其可能与ATAF亚类基因具有相似的功能。拟南芥AtNAC3亚类若干AtNAC19、55、72基因受水分影响诱导表达,且过表达该基因可提高植株抗旱性[37]。Fujita等[38]对拟南芥进行脱水处理,分离出一个编码NAC 蛋白AtNAC3 亚组的AtNAC072基因,在干旱、高盐胁迫诱导下高表达。CoNAC56分布于AtNAC3亚组,与拟南芥AtNAC072、AtNAC019和AtNAC055有较近的同源关系。水稻ONAC003的表达受干旱诱导,其过表达株系在高温、干旱抗性增强,表明ONAC003调控水稻非生物胁迫过程[39]。油茶CoNAC28基因归于ONAC003亚组,在干旱胁迫下的表达模式相似,具有相似的亲缘关系,表明它们在干旱胁迫下具有相似的功能。陈琼琼等[40]采用实时定量qRT-PCR 表达分析显示,10个NAM基因在不同组织中均特异表达,大部分NAM 基因在盐和干旱胁迫下表达上调。CoNAC51和CoNAC52分布于NAM亚组,它们可能具有极为类似的蛋白功能在干旱胁迫时都表现为上调。在干旱胁迫下,油茶中的大部分ATAF、AtNAC3、ONAC003、NAM 亚组成员上调表达,暗示这些NAC 蛋白极有可能在油茶抗干旱胁迫中发挥重要作用。
此外,基于RNA-Seq 和qPCR 分析,本研究验证了大部分候选CoNAC基因在油茶干旱胁迫时的表达模式与转录组数据结果较为相似,整体呈现为在干旱下基因表达上调,复水后下调。此外,本研究发现,候选CoNAC51和CoNAC52基因在长林18 和长林53 表达趋势在干旱和复水后存在差异。课题组前期对江西主推油茶品种在干旱胁迫下的生理和形态数据结果显示,长林53 耐旱性较强,而长林18 耐旱性相对较弱。这与冯士令[41]对长林系列的9 个油茶品种幼苗的抗旱性综合评价结果相似。笔者的研究结果表明,在长林18 中CoNAC52在干旱胁迫和复水后表现为先下调后上调,而CoNAC51基因的表达量呈上升趋势,它们在长林53中的表达模式均呈现随着干旱胁迫上调,复水后下调。而这2个CoNAC基因的所在亚组,在其他物种中已证实在对干旱胁迫起到正调控作用[42]。由此说明,在长林18中CoNAC52基因的表达调控对水分敏感程度较低,表现为下调表达,在长林53中对水分较为敏感,基因表达上调。笔者的研究结果从一定程度上说明由于不同耐旱油茶品种的CoNAC基因表达对水分的敏感程度不同,应对干旱胁迫的响应机制存在差异。
4 结论
本研究的结果丰富了油茶NAC 家族转录因子的基本信息,对其蛋白理化特征、保守基序和基因结构等进行了分析。多数CoNAC基因保守结构域位于蛋白N 端,与其他植物中NAC 的结构特征相符。CoNAC基因启动子区具有多种胁迫响应元件,系统进化树将67 个CoNAC划分为17 个亚类,同一亚组在基因结构方面具有相似性且同一或不同亚类CoNAC基因在干旱胁迫处理后呈现差异表达。基于公共数据库中油茶干旱RNA-Seq 数据,筛选油茶干旱胁迫相关的42 个NAC基因的TPM 值,显示不同NAC基因在干旱胁迫下具有不同的表达程度。qPCR 结果显示,6 个油茶NAC基因在干旱胁迫下呈现大致相似的表达趋势,而CoNAC52和CoNAC51基因表达存在品种差异,说明其基因表达在不同油茶品种中对水分的敏感程度不同,不同油茶品种对干旱胁迫的响应机制存在差异。本研究为更好的探讨油茶耐旱胁迫响应机制提供一定的参考,同时也能为油茶进一步抗旱品种选育和遗传改良提供良好的基因资源。
猜你喜欢
中国银幕(2022年4期)2022-04-07
国际放射医学核医学杂志(2020年4期)2020-07-27
中国中医基础医学杂志(2020年1期)2020-03-03
心电与循环(2020年1期)2020-02-27
中国医学影像技术(2019年10期)2019-10-24
浙江林业科技(2019年3期)2019-09-18
中南林业科技大学学报(2017年9期)2017-12-11
上海农业学报(2017年3期)2017-04-10
中国全科医学(2015年26期)2015-02-23
