
氮肥基追比例对春小麦茎鞘非结构碳水化合物及产量的影响
2022-01-08 07:06王海琪蒋桂英黄艺华王荣荣马艳梅晏世杰
麦类作物学报 2021年12期
王海琪,蒋桂英,黄艺华,王荣荣,马艳梅,晏世杰
(石河子大学农学院,新疆石河子 832000)
小麦(TriticumaestivumL.)是新疆最重要的粮食作物,2020年全疆小麦种植面积达到1.62×106hm2,约占粮食作物总播种面积的 48.18%[1],小麦的稳产高产对新疆粮食安全具有重要意义。氮素是影响作物生长发育的关键营养元素,其施入量、施入时期及追施比例直接影响着小麦同化物的积累与转运[2]。适宜的氮肥运筹因环境而异,长江中下游流域麦区施氮量一般为220~250 kg·hm-2,基追比例6∶4有利于同化物的积累与转运及高产[2-4];华北平原麦区适宜的氮肥施用量一般为270 kg·hm-2左右,基施 肥∶拔节肥∶孕穗肥6∶3∶1有利于花后维持较高的物质生产能力和高产[5]。适宜的氮肥基追比例可实现氮素的高效利用和小麦高产。
茎鞘是谷类作物重要的物质贮藏和转运器官[6],其对光合物质的积累、分解、运转能力影响小麦籽粒产量[7]。小麦花前、花后贮藏在茎鞘的光合同化物大多为非结构性碳水化合物(non-structural carbohydrate,NSC),约占茎鞘干重的40%以上[8],对籽粒干物质贡献率在适宜条件下为 11%~47%,在逆境下可到达29%~100%[9]。适宜的施氮方式有利于更多的贮藏物质由茎鞘向籽粒中转运,而氮肥过多或过少则不利于其积累和转运[10]。长江中下游麦区的氮肥施用量为240 kg·hm-2、基追比例为5∶5时,茎鞘贮藏物质在花前、花后积累总量最大[11]。不同小麦品种的花前干物质的转运能力存在显著的差异,高产型小麦品种的花前干物质积累量及其运转率较低产型高[12]。贮藏在茎鞘中的NSC主要为果聚糖和蔗糖[13]。果聚糖占NSC含量的60%~85%[14],反映了茎鞘贮藏NSC的能力[15],并作为信号物质参与小麦抗逆代谢调控[15-16]。小麦植株中果聚糖的转运和代谢是复杂的生理过程,其变化受自身生长节律和外部因素的双重调控[17]。侯有良等[18]认为,悉尼地区的高蛋白质基因型小麦品种在增施氮肥后(0~200 kg·hm-2)可显著降低植株内果聚糖的累积与运转速度。已有研究证明,果聚糖代谢主要受蔗糖果糖基转移酶(sucrose fructosyltransferase,SST)和果聚糖外水解酶(fructan exohydrolase,FEH)调控[17,19]。当茎鞘干重降低时,其中的果聚糖经SST酶催化合成[20],由FEH酶分解转运,最终形成籽粒产量[21]。姜 东等[22]在华北麦区发现,在施氮量为210~330 kg·hm-2范围内,增加施氮量可增加小麦茎鞘中果聚糖含量,提高SST酶活性,抑制FEH活性,但不利于成熟期小麦茎鞘中果聚糖的分解和运输。可见,氮肥量及施用比例对小麦茎鞘中果聚糖代谢以及籽粒产量有很大的影响。
实现水、肥、药一体化施用与田间智能管理、降低人工与投入成本的滴灌技术,已成为新疆小麦生产的主要栽培技术。迄今,滴灌条件下氮肥基追比例对小麦茎鞘中贮藏物质的积累与再转运动态规律鲜有报道,对贮藏物质及其相关酶的影响及其与粒重的关系尚不清楚。为此,本试验在前人研究的基础上,选取新疆当地强筋和中筋小麦品种为材料,研究茎鞘中NSC、果聚糖的代谢特征,分析花后关键生育阶段茎鞘中NSC和果聚糖积累和再转化动态规律,以期了解茎中NSC和果聚糖向籽粒转运的氮素响应机制,为新疆麦区滴灌小麦氮肥的高效利用和高产栽培提供科学 依据。
1 材料及方法
1.1 试验区概况
试验于2020年4月-7月在新疆石河子市石河子大学农学院实验站(85°59′ E,44°18′ N)进行。石河子多年平均气温为7.5~8.2 ℃,多年平均降水量208 mm,平均蒸发量1 660 mm,属于典型的大陆性气候,2020年温度略高于多年平均温度,属于常规年型,具体见图1。供试土壤类型为灌溉灰漠土,其中0~20 cm土层养分基本状况为: 有机质28.4 g·kg-1,全氮1.3 g·kg-1,碱解氮71.3 mg·kg-1,速效磷15.2 mg·kg-1,速效钾159 mg·kg-1。

图1 小麦生育期间日平均温度和降雨量
1.2 试验设计
试验采用裂区设计,小麦品种为主区,供试品种为中筋型品种新春6号(XC6,蛋白质含量 13.5%)和强筋型品种新春37号(XC37,蛋白质含量 16.3%);氮肥基追比为副区,设置5种氮肥处理,具体见表1。各处理重复3次,小区种植面积为12 m2(3 m×4 m),各个小区间埋置100 cm深度的防渗膜,防止肥料外移。生育期间施氮处理施用纯氮250 kg·hm-2。除不施肥处理外,其余处理的各小区在播前将120 kg·hm-2的P2O5(磷酸二铵)作基肥施入,再根据各处理的基追比例,用尿素对基施和追施的施氮量进行调整。全生育期共灌水9次,总灌溉量为6 000 m3·hm-2,各个时期的灌溉量通过水表精确控制。小麦于2020年3月28日播种,播量为345 kg·hm-2,采用宽窄行、“一管4行”方式种植,行间距为 12.5+20+12.5+15 cm(如图2),滴灌带(管径16 mm,滴头间距30 cm,流量2.6 L·h-1)放置在20 cm的宽行,其他田间管理同当地大田生产。于2020年7月6日收获。

图2 滴灌带布置示意图

表1 各处理间氮肥基追肥施用量
1.3 测定指标与方法
1.3.1 茎鞘干物质测定及相关指标计算
开花期挂牌标记同一日开花的小麦。于开花期和成熟期选取10株有代表性的小麦单茎,置于105 ℃烘箱内杀青30 min,于70 ℃下烘干至恒重。将茎鞘分为穗下节(穗下第一个节间及其基部节),倒二节(穗下第二个节间及其基部节)和其余节,分别测量每节重量。开花前茎鞘同化物的转运量、转运效率及其对籽粒产量贡献率的计算公式参照Bonnett[23]方法。
1.3.2 NSC含量的测定及相关指标计算
于花后0 d(开花期)、7、14、21、28和35 d(成熟期),取同一日开花的5株小麦单茎,将茎鞘按穗下节、倒二节和其余节分样,参照杨建昌[24]及马召朋[25]的方法测定NSC含量并计算NSC花前、花后转运效率及其对粒重贡献率。
NSC积累量=NSC含量×干物质量;
花前NSC转运效率=(开花期NSC积累量-成熟期NSC积累量)/开花期积累NSC量×100%;
花后NSC转运效率=(最大NSC积累量-开花期NSC积累量)/最大NSC积累量×100%;
花前NSC对籽粒粒重贡献率=(开花期NSC积累量-成熟期NSC积累量)/(1 000×主穗粒重)×100%;
花后NSC对籽粒粒重贡献率=(最大NSC积累量-开花期NSC积累量)/(1 000×主穗粒重)×100%。
1.3.3 果聚糖相关酶活性及含量的测定及计算
在花后0 d(开花期)、7、14、21、28和35 d(成熟期)取标记小麦10株,将茎鞘按其余节、倒二节和穗下节分样,其中5株用于测定果聚糖含量;另外5株小麦鲜样置入液氮,随后于-80 ℃保存,用于测定果聚糖代谢关键酶活性。
蔗糖果糖基转移酶(SST)和果聚糖外水解酶(FEH)活性测定参照Verspreet[14]方法;果聚糖含量测定参考刘燕琼等[26]方法。果聚糖花前、花后转运效率及其对籽粒产量贡献率计算参照杨德龙等[27]的方法。
1.3.4 产量的测定
于小麦成熟期随机选取1 m2调查小麦穗数,从中随机选取20个植株,测定穗粒数和千粒重。小区人工收割,籽粒晒干、称重后计算籽粒产量。重复3次。
1.4 数据分析
使用Microsoft Excel 2016进行数据处理和图表绘制,利用SPSS 22.0进行统计分析,运用Duncan及LSD方法进行显著性检验。
2 结果与分析
2.1 氮肥基追比对小麦不同节位的果聚糖代谢和粒重影响
2.1.1 对果聚糖关键酶活性的影响
由表2可知,不同氮肥基追比处理下,两个小麦品种植株不同节位的SST活性均随花后天数增加均呈先上升后降低的趋势,花后21 d最大,花后35 d(成熟期)最小;除成熟期外,不同处理下SST活性其他时间均表现为N37>N46>N55>N28>N00。与对照(N00)相比,其余节SST活性在基追比处理N37时差异显著(P< 0.05),与其他处理差异不显著。各处理SST活性均表现出随节位降低而升高的趋势,即其余节>倒二节>穗下节,即其余节活性 (2.04~14.31 mg·g-1DW)最高,分别比倒二节、穗下节高出38.01%,101.33%。两品种SST活性变化趋势一致,但整体表现为XC37高于XC6。
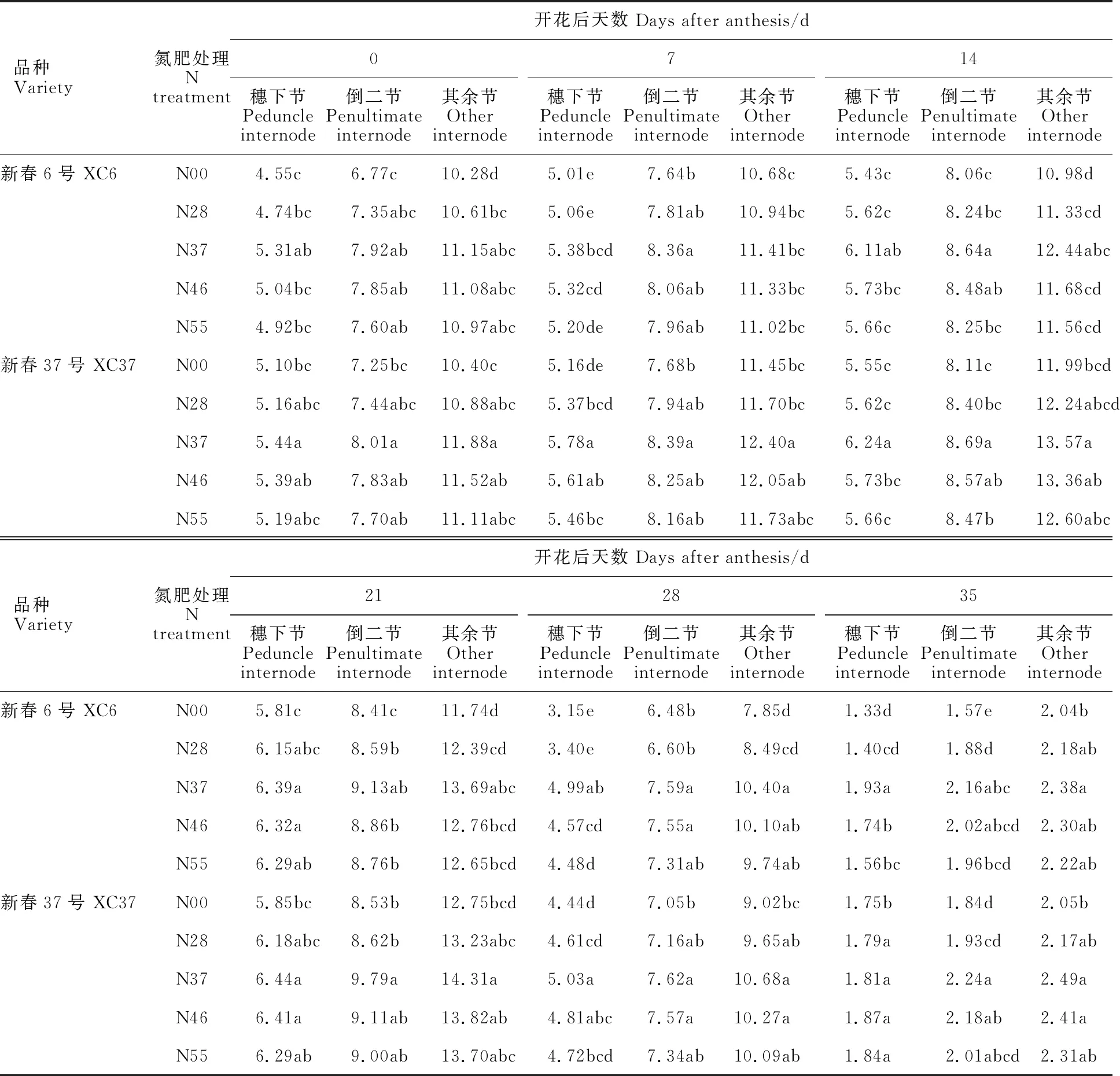
表2 氮肥基追比例对春小麦穗下节、倒二节和其余节蔗糖果糖基转移酶(SST)活性的影响
不同氮肥基追比下,小麦植株不同节位的FEH活性随花后天数增加呈“降-升-降”的变化,花后14 d最低,28 d最高,35 d成熟期快速下降(表3),各处理间均表现为N37>N46>N55>N28>N00。除成熟期外,各节位FEH活性在基追比为3∶7时,与对照(N00)差异显著(P< 0.05),与其余处理差异不显著。FEH活性随小麦节位的下降而升高的趋势(除花后28 d),表现为其余节活性(71.32~85.30 mg·g-1DW)最高,分别比倒二节、穗下节高出3.28%、19.26%。两品种FEH酶活性变化趋势一致,表现为XC37高于XC6。
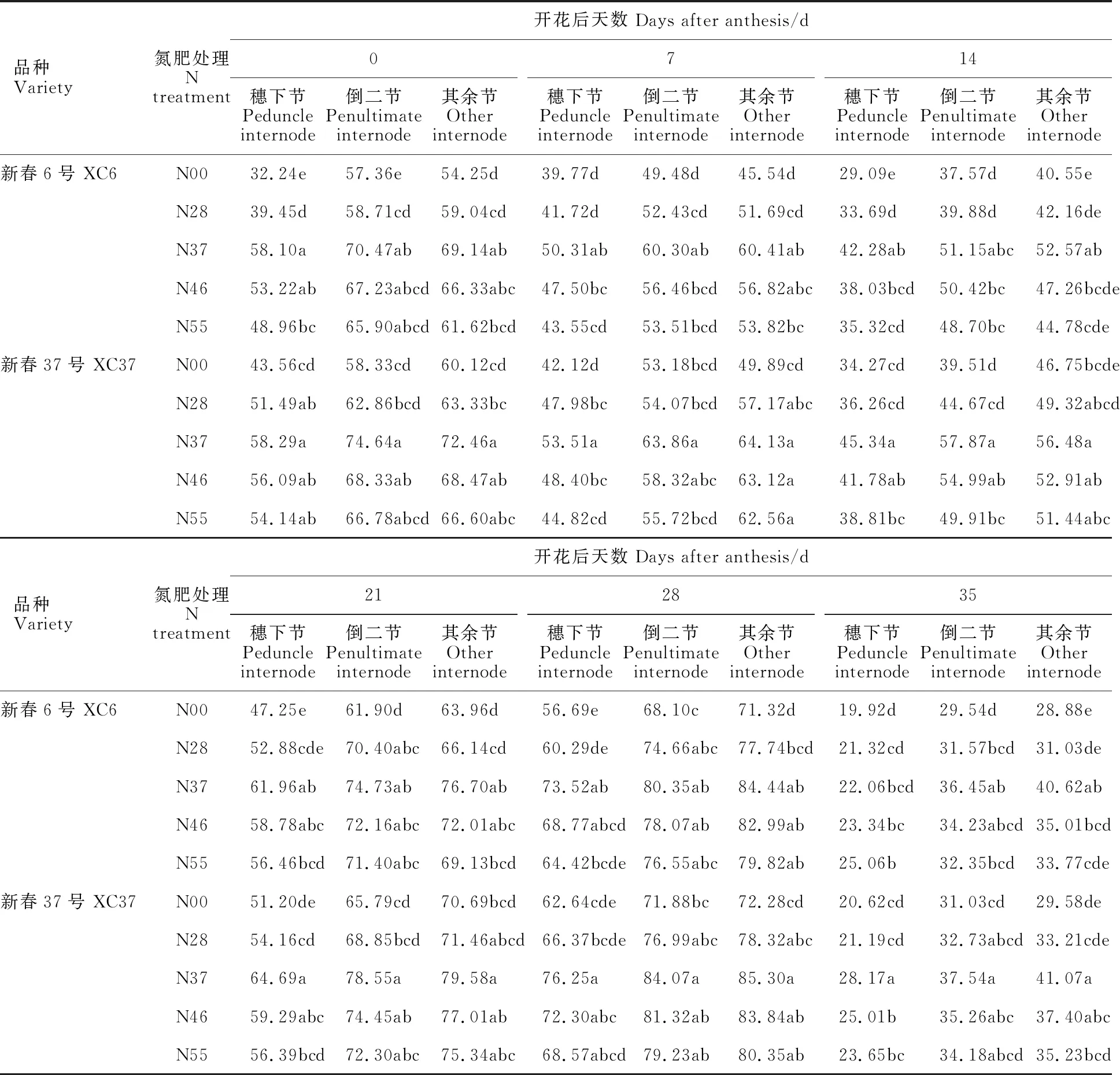
表3 氮肥基追比例对春小麦穗下节、倒二节和其余节果聚糖外水解酶(FEH)活性的影响
2.1.2 果聚糖含量的变化
由图3可知,在不同氮肥基追比处理下,小麦茎鞘不同节位的果聚糖含量随花后天数增加呈先升后降的趋势,花后21 d最高,35 d最低,表现为N37>N46>N55>N28>N00,且各节位果聚糖含量均以N37处理显著高于其他处理(P< 0.05),分别比N00、N28、N46、N55处理高 68.07%、 35.62%、17.58%、25.32%。花后21 d时其余节果聚糖含量最高(76.84~116.51 mg·g-1DW),分别比倒二节和穗下节高 11.48%~16.76%和22.76%~40.35%。两品种果聚糖含量变化趋势一致,表现为XC37大于XC6。

图3 氮肥基追比例对滴灌春小麦穗下节(A,D)、倒二节(B,E)和其余节(C,F)果聚糖含量的影响
2.1.3 果聚糖转运对粒重的贡献
由表4可知,就果聚糖转运而言,品种对被测茎节的花前转运效率无显著影响,对穗下节和其余节的花后转运效率及花前和花后转运对粒重的贡献率均有显著效应,氮肥基追比对被测指标均有显著效应(其余节的花后转运效率除外)。各氮肥基追比中,除XC6的倒二节花前转运对粒重贡献率外,N37处理下不同节位的花前和花后果聚糖转运效率及其对粒重的贡献率与对照(N00)相比均存在显著性差异(P<0.05),其中,两个品种的倒二节和其余节的花前转运效率和穗下节的花后转运效率均显著低于对照,其他被测指标均显著高于对照。不同节位间果聚糖转运效率略有差异,其余节果聚糖转运效率(41.71%~78.62%)和贡献率 (4.27%~43.42%)高于穗下节 (33.31%~ 65.57%,1.54%~17.57%)和倒二节(33.33%~ 71.94%,3.18%~28.7%)。两品种花后果聚糖转运效率及其对粒重贡献率均高于花前,XC37转运效率及对穗粒重贡献率整体高于XC6。

表4 氮肥基追比例对滴灌春小麦各节花前和花后果聚糖转运效率、贡献率的影响
2.2 NSC转运对粒重的影响
由表5可以看出,就NSC而言,品种对倒二节花前和花后转运效率影响不显著,对其他被测指标均有显著效应(P<0.05);氮肥基追比对被测指标均有显著影响。各氮肥基追比中,不同节的花前、花后NSC转运效率及其对粒重贡献率均在N37处理下达到最大值,且均显著高于相应N00处理;花前、花后贡献率表现出N37>N46>N55>N28>N00趋势。NSC转运效率达到 50.42%~70.27%,对粒重贡献率为2.77%~53.94%。不同节位花前和花后NSC转运效率略有差异,其余节NSC转运效率(63.54%~ 70.27%)和贡献率 (8.34%~53.94%)较高,穗下节(50.42%~ 66.89%,3.61%~25.17%)次之,倒二节 (61.44%~67.80%,2.77%~ 19.04%)较低。两品种不同节位NSC花前转运效率及其对粒重贡献率普遍低于花后;XC37的转运效率、对粒重贡献率基本高于XC6。

表5 氮肥基追比例对小麦各节花前和花后NSC转运效率、贡献率的影响
2.3 茎鞘干物质的转运特点
由表6可知,品种和氮肥基追比对被测指标均有显著影响,二者对被测指标无显著互作效应。各氮肥基追比处理下,小麦茎鞘不同节间的干物质转运量略有差异,N37处理与N00处理相比显著提高了两个品种倒二节和其余节的干物质转运量(P< 0.05)。小麦茎鞘不同节的干物质转运量对籽粒产量贡献率随茎节下降呈增加趋势,即其余节>倒二节>穗下节;与N00处理相比,N37处理提高了倒二节和其余节干物质转运效率、对籽粒产量贡献率,降低了穗下节干物质转运效率、对籽粒产量贡献率。两品种干物质转运效率、对籽粒产量贡献率均以XC37高于XC6。

表6 小麦茎鞘干物质的转运率及其贡献率
2.4 氮肥基追比对春小麦籽粒产量的影响
从表7可以看出,产量受氮肥基追比及其与品种互作的影响显著,两个品种产量及构成均随氮肥基追比例的增加呈先增后降的趋势,表现为N37>N46>N55>N28>N00,其中N37处理与其他处理差异显著(P<0.05);XC37在N37处理下的千粒重、穗粒数、穗数及产量分别比N00处理高19.89%、46.77%、34.69%、53.59%;XC6在N37处理下分别比N00高20.45%、38.21%、 32.48%、52.68%。两品种相比,XC37产量及构成高于XC6。说明适宜的氮肥基追比例(3∶7)能够增加小麦的千粒重、穗粒数、穗数和产量。

表7 滴灌春小麦产量及产量构成因素的变化
3 讨 论
3.1 氮肥基追比例对滴灌春小麦茎鞘果聚糖及其关键酶活性的影响
果聚糖可占小麦茎鞘干重的40%以上,是小麦茎鞘中关键性的储藏型碳水化合物[28],最高可达茎鞘NSC的85%[9]。果聚糖代谢受自身和外界因素的双重调控[29],同时影响籽粒重量[14,27]。SST可调控小麦体内果聚糖的合成,FEH则调控其降解[1]。姜 东等[17,19,22]研究认为,在花后 15~20 d小麦茎鞘中果聚糖的积累量可达到最大,且此时相关酶活性较高,花后10 d FEH酶活性开始升高,25~30 d达到分解高峰。本研究中,果聚糖含量及SST活性均在花后21 d达最大值,而FEH活性在 28 d达到最大,与姜 东等[22]研究结果基本一致。前期茎鞘中FEH活性较低,分解能力较弱,利于开花前期果聚糖在茎鞘中积累,开花后期FEH活性逐渐升高,导致籽粒发育后期果聚糖含量迅速降低。
适宜的施氮量不仅可以促进小麦茎鞘中果聚糖积累,还可促进其向籽粒的运转;而过高的氮则抑制果聚糖累积和运转,最终造成减产[10,30]。本研究发现,不同氮肥基追比下小麦茎鞘中的果聚糖含量转运效率变动于33.31%~77.39%,对粒重的贡献率在1.54%~43.42%之间,小麦各茎节都积累较多的果聚糖,以其余节和倒二节最多,不同茎节花后转运对粒重贡献率普遍高于花前。杨德龙[27]发现,在正常灌溉处理下果聚糖花后转运效率普遍高于花前,这与本试验结果一致。各氮肥基追比中以N37处理的果聚糖含量及SST、FEH活性最高,且均显著高于N00处理。说明适宜的氮肥基追比例(3∶7)促进了各节中果聚糖含量的合成和积累以及达到峰值后的降解,从而促进了果聚糖运转。
3.2 氮肥基追比例对滴灌春小麦茎鞘NSC转运的影响
小麦茎鞘中NSC对产量形成尤为重要,通常对粒重贡献率可达20%以上[29]。籽粒产量与茎鞘暂贮物质的积累与运转密切相关[31]。马尚宇等[32]认为,茎鞘中贮存的NSC花前转运对粒重贡献率可达到3%~30%,花后转运对粒重贡献率为10%~25%。本研究发现,不同氮肥基追下的NSC花前、花后转运对粒重贡献率可达 2.77%~13.28%;7.89%~ 53.94%,高于上述研究。可能因小麦茎鞘中NSC含量的高低受环境因素调控[33]。马召朋[25]认为,干旱促进了小麦茎鞘花前NSC转运以及对粒重贡献率;Bahrani等[34]在地中海地区发现,适宜的氮肥施用量(80 kg·hm-2)可促进小麦茎鞘中的NSC转运效率的提高,而过量氮肥(160 kg·hm-2)则会降低NSC转运效率。也有人认为,在长江中下游麦区施氮量为240 kg·hm-2下,茎鞘暂贮物在基追肥比例由2∶8上升为5∶5时可使花前、花后转运效率提高14.64%和 18.55%[11]。本研究发现,施氮量为250 kg·hm-2的条件下,基追比为3∶7时,可获得较高的NSC转运效率及其对粒重贡献率。当基追肥比例过大时,不利于其对粒重的贡献。可见,适宜的氮肥基追比例能够显著提高NSC转运以及对粒重贡献率。两品种小麦对不同氮肥基追比例的响应也有所不同,XC37其余节、倒二节的NSC花前转运效率增幅( 5.44%, 8.36%)高于XC6(4.47%, 7.86%);XC37对粒重的贡献率(3.14%~ 53.94%)的普遍高于XC6(2.77%~42.67%)。两品种小麦花前转运效率、贡献率普遍低于花后。
3.3 氮肥基追比例对滴灌春小麦茎鞘干物质及产量的影响
茎鞘是同化物临时贮藏和转运的器官,其物质转运能力和积累量与小麦籽粒产量关系密切[7]。增施氮肥可促进茎鞘中的同化物向籽粒中运输分配[35]。王月福等[36]发现,在适宜施氮量(180 kg·hm-2)内,增加施氮量可促进花前贮藏物质向籽粒运转;但氮肥过量则会导致茎鞘干物质花后转运效率下降9.14%~15.46%,这可能是与品种特性、土壤肥力不同有关。张向前等[2]认为,长江中下游小麦在施氮量一定情况下,以基追比为6∶4时产量及干物质积累量最高。姜丽娜等[37]则发现,豫中小麦在氮肥施用量为270 kg·hm-2时,基追比例为3∶7,可促进小麦产量的提高。本试验结果认为,小麦不同茎鞘贮藏物质转运效率对氮肥基追比例的反应有所不同,氮肥基追比例3∶7处理可显著提高两品种小麦倒二节和其余节的干物质转运量,降低穗下节干物质转运量,获得最大产量。说明同一小麦品种不同部位节距离穗部越远,茎鞘中贮藏干物质的转运量增幅越大;转运量越大对粒重的影响也越大。XC37倒二节和其余节干物质对籽粒产量贡献率高于XC6,这主要与小麦品种有关。
猜你喜欢
中国果业信息(2022年1期)2022-11-28
作物学报(2022年1期)2022-11-05
麦类作物学报(2022年6期)2022-08-09
农业工程学报(2022年7期)2022-07-09
军事运筹与系统工程(2020年2期)2020-11-16
现代农业研究(2019年3期)2019-06-05
吉林农业(2017年7期)2017-07-12
领导文萃(2017年11期)2017-06-12
农民致富之友(2017年9期)2017-05-21
