
根系功能性状对干瘠立地适应的种间差异
——以北京石质山地主要观赏树种为例
2021-12-25 01:20:16韦柳端朱济友李夏榕孙广鹏张新娜徐程扬
生态学报 2021年23期
韦柳端,朱济友,李夏榕,孙广鹏,张新娜,徐程扬,*
1 北京林业大学城市林业研究中心, 北京 100083 2 干旱半干旱地区森林培育及生态系统研究国家林草局重点实验室, 北京 100083
干旱、瘠薄是困难立地的重要特征,也是限制林木生长的主导因子。北京浅山区以低山丘陵地貌为主,森林植被以人工林为主。近些年来,由于气候变化等原因,北京市山地森林树木频发生长衰退、甚至个体死亡现象。北京周边地区绿地建设和观赏树种培育对北京市的生态环境改善和美化有着积极的作用,但由于该区域降水量少、土层浅、土壤保水较差、水土流失严重,造成了干瘠的立地环境,对林木的生长极为不利。养分和水分常常同时限制干瘠环境下植物的生长[1],根系是树木生长最基本、最重要的结构,树木根系形态可反映树木对生境的适应策略和土壤的固定、保持作用[2]。因此,通过测量干瘠环境中树木根功能性状(Root functional traits, RFT),进一步研究树木根系对干瘠立地的响应以及树木对干瘠立地土壤资源获得对策,一直是近年来研究的热点。
根的形态特征和空间分布是影响水分、养分吸收的重要因素[3],根直接与土壤接触,更易对土壤环境做出反应[4]。在植物生长阶段环境因子对根系功能性状产生重要影响,处于不同环境资源的植物根系外部形态和内部生理机能会发生变化,导致根系同一性状的特征有所差异,从而表现出不同的生长方式和适应策略[5- 6]。根系的大小、形态结构以及分布范围等直接决定着植物对土壤资源的吸收和利用能力[7- 8],同时决定了植物对不利环境的适应,成为植物生长的关键控制因素之一。它会随着环境胁迫的改变而进行调整,以此获取最大的资源量[9]。根系构型是植物根系对环境响应和适应的具体表现,干瘠环境中生长的植物通常在幼苗期根系就开始快速地向下延伸,以获取土壤深层的水分,这对于植物的生长极为重要[10- 12]。
目前关于根系构型的研究主要位于西北干旱地区[13-15],以干旱胁迫中植物根系的形态变化为主,对养分胁迫下植物根系的适应对策甚少,而且大多数都以田间模拟试验为主,难以还原真实环境下植物根系形态特征的反应,在真实环境下不同树种根系性状对不同干瘠的反应方面报道极少。本研究主要探讨:(1) 在不同干瘠环境中树木根系生长特性、抗(耐)干瘠程度在不同树种间是否存在显著差异;(2) 不同树种对干瘠的响应方式和强度是否一致。本文以北京石质山地常见的3种观赏树种(黄栌(C.coggygria)、山桃(Amygdalusdavidiana)、栓皮栎(Quercusvariabilis))为研究对象,分析其根系连接长度、分支强度、组织密度、比根长、比根表面积、根系密度,从根系形态的变化揭示不同树种对干瘠的适应对策,为干瘠立地植被恢复适树适地提供理论依据和研究思路。
1 材料与方法
1.1 试验材料
试验区设置在西山国家森林公园(39°58′N,116°11′E),位于北京西郊小西山,为石质山区,属低山区。黄栌(C.coggygria)是漆树科黄栌属,是中国重要的观赏红叶树种,耐干瘠和碱性土壤,根系发达,萌蘖性强;山桃(Amygdalusdavidiana)属李亚科桃属,耐干瘠,对土壤适应性强;栓皮栎(Quercusvariabilis)是壳斗科栎属,抗旱、耐瘠薄,适应性强,在酸性、中性及钙质土壤均能生长,具有深根性。本文以林龄(中龄林)和林分密度相对一致的黄栌、栓皮栎、山桃人工林作为研究对象,林分内近期没有进行过抚育采伐,避免根系生长受到人为干扰。
1.2 样品采集及处理
1.2.1采样方法
于2019年7—8月份进行野外取样。在林分内设置一个20 m×20 m的临时样地,共获得128个样地数据,其中黄栌有57个、山桃36个、栓皮栎35个。每个样地内按照“S”字型选取5株平均木,从树冠外缘往里范围30 cm×50 cm内取细根,并在两林木之间挖一个30 cm×30 cm的土坑,将土坑内的根系全取;在调查样地沿对角线等距三点取0—10cm土层环刀土样,并沿土壤垂直剖面取0—30cm混合土样,各取3份。将根系样品置自封袋、土壤样品置信封中,带回实验室处理。
1.2.2干瘠程度划分
在实验室采用环刀控水法测量田间持水量,称重法测量石砾含量,根据调查样地田间持水量和石砾含量的大小对干瘠程度划分,如表1所示:

表1 调查样地干旱程度和瘠薄程度划分
1.2.3林木根系性状的测定与计算
根系清洗干净后,用根系扫描仪Epson Perfection V800对根系扫描,扫描时将根系放入透明托盘,摆放时尽量避免根系互相重叠和缠绕。利用图像分析软件Winrhizo Pro 2004a测定细根表面积(cm2)、体积(cm3)、根长(cm)等[16],根系连接长度(Root link length, RL)利用游标卡尺测量,然后将样本装于牛皮纸信封内置于烘箱内70 ℃烘干至恒重,获得细根干重。相关性状的计算方法如下:
分支强度(Branching intensity, BI): 分叉数/根系长度
(1)
根组织密度(Root tissue density, RTD: 根体积/干重
(2)
比根长(Specific root length, SRL): 根长/干重
(3)
比根表面积(Specific root area, SRA): 根系表面积/干重
(4)
根表面积密度(Root area density, RAD): 根表面积/土壤体积
(5)
根长密度(Root length density, RLD): 根长/土壤体积
(6)
1.3 数据处理
利用Microsoft Office Excel 2013统计数据并绘制图表。利用SPSS分析软件对不同干旱、瘠薄土壤条件中根系形态指标进行差异性检验(显著性水平α =0.05),双因素方差分析(Two-way ANOVA)研究干旱、瘠薄以及二者的交互作用对树木根系功能性状的影响,结合LSD、Duncan对根系性状进行多重比较(P≤ 0.05)。
2 结果与分析
2.1 土壤干旱、瘠薄及其交互作用对树木根系功能性状的影响
由表2可知,干旱、瘠薄及其交互作用对3种树种的根系功能性状均影响显著,尤其是对山桃、栓皮栎的根系功能性状具有极显著影响。干旱对黄栌RL、RAD具有显著影响,对SRL、RLD具有极显著影响。瘠薄对其SRL、SRA影响显著,对RAD、RLD具有极显著影响,二者的交互作用对黄栌RAD、RLD具有极显著影响。干旱、瘠薄及其交互作用对山桃BI、SRL、SRA、RTD、RAD、RLD均具有极显著影响,干旱、瘠薄的交互作用对山桃RL影响不显著,但干旱、瘠薄两因素对山桃RL影响均极显著。干旱对栓皮栎RL、BI、SRA、RTD、RAD、RLD影响极显著,瘠薄对其RL、SRL、SRA、RAD、RLD影响极显著,二者的交互作用对SRL、SRA显著影响,对RLD、RL、BI影响极显著。
根据表2中F值,干旱、瘠薄及其二者的交互作用对3种树木根功能性状的影响程度各不相同,干旱、瘠薄对大部分根系性状的作用均强于二者的交互作用。从黄栌的分析结果可知,干旱、瘠薄以及二者的交互作用对黄栌根功能性状的影响程度相差不大,但干旱、瘠薄对黄栌SRL的影响远大于二者的交互作用。对于山桃,干旱和瘠薄对RL、BI、SRL、SRA、RAD、RLD的影响远大于二者的交互作用,其中干旱对RL、SRL的影响程度大于瘠薄的作用,瘠薄对SRA和RLD的影响程度大于干旱,干旱、瘠薄及二者的交互作用对山桃RTD的影响程度相差不大。瘠薄对栓皮栎RL、SRL、SRA、RAD、RLD的影响大于干旱和交互作用,干旱对RTD的影响大于瘠薄和交互作用,二者的交互作用对BI的影响大于干旱、并远大于瘠薄作用,干旱和瘠薄对栓皮栎RL、SRA、RAD的影响远大于两者的交互作用。

表2 干旱、瘠薄及其交互作用对树木根功能性状影响的方差分析
2.2 树木根系功能性状在不同干瘠环境中的变化
2.2.1连接长度
不同根功能性状和不同树种随土壤干旱、瘠薄的加剧,表现出不同的变化,相同性状在不同树种中具有显著差异(图1、图2)。从轻度干旱到极度干旱,3种树种的RL均逐渐减小,黄栌减少不显著;山桃RL显著增大后又显著减小,中度干旱比轻度干旱增加7.2% (P<0.01),而重度干旱、极度干旱分别比中度干旱减小26.9% (P<0.01)、41.7% (P<0.01);栓皮栎RL随干旱增强显著减少后极显著增大,在极度干旱极显著减小达到最小值。在土壤逐渐瘠薄的条件下黄栌RL在中度瘠薄减小后几乎保持不变,变化不显著;山桃和栓皮栎RL均在极度瘠薄时显著减小。
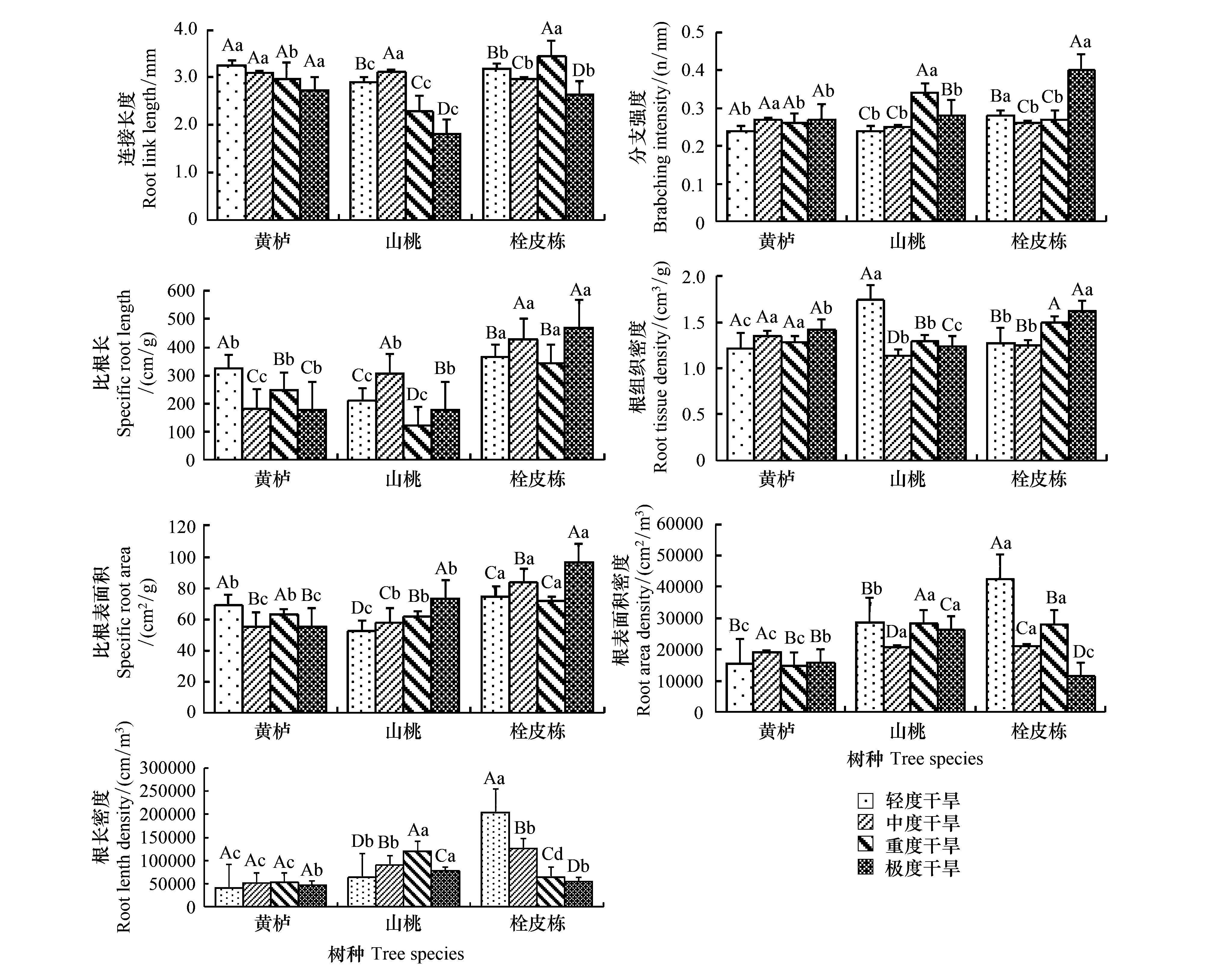
图1 不同树种根系功能性状对干旱胁迫的响应Fig.1 Response of root functional traits of different tree species to drought stress大写字母表示同一树种不同干旱水平之间的差异显著性,小写字母则表示不同树种间在同一干旱水平上的差异显著性

图2 不同树种根系功能性状对瘠薄胁迫的响应Fig.2 Response of root functional traits of different tree species to barren stress
2.2.2分支强度
BI与RL的变化呈现出相反的趋势,随着干旱的严重加剧,BI变大,在不同的瘠薄条件下,变化不明显。黄栌的BI在不同干旱、瘠薄环境中变化不显著,其变化规律一致,从轻度到中度BI增强,随后减弱后又增强,在中度干旱达到最大值;山桃随着干旱的加剧,其BI显著增大,当在极度干旱减少,在重度干旱时BI最大。山桃BI在瘠薄环境中变化不一致,但中度、极度瘠薄BI均比轻度瘠薄大,分别增加17.9% (P<0.01)、7.4% (P<0.05);栓皮栎BI随着干旱、瘠薄的加剧整体变化趋势均为随之增强,在极度干旱和瘠薄中达到最大值,分别比轻度增加了42.9% (P<0.01)、25.6% (P<0.01)。
2.2.3比根长
SRL在轻度、极度干旱出现最大值或最小值,在土壤中度或重度干旱条件下SRL显著增大,随瘠薄的加剧,总的变化趋势是随之减小,但也会呈现出相反的变化趋势。黄栌和山桃SRL在随着干旱、瘠薄的加剧,表现出一致的规律,即从轻度到极度,其SRL随即减小,但是在不同瘠薄情况下的变化规律较明显。在不同干旱下黄栌和山桃SRL的最大值分别出现在轻度、中度干旱。相反,栓皮栎SRL随着干旱、瘠薄的加剧而随之增大。栓皮栎在重度、极度干旱SRL分别比轻度干旱增加17.6% (P>0.05)、28.8% (P<0.05),重度干旱减少5.5% (P>0.05);栓皮栎在中度、重度、极度瘠薄SRL分别比轻度瘠薄增加54.3% (P<0.01)、10.8% (P>0.05)、45.5% (P<0.01),其中在中度瘠薄栓皮栎SRL最大。
2.2.4根组织密度
黄栌、栓皮栎RTD都随着干瘠的加剧而减小,而山桃随着干瘠的加剧而增加。黄栌在不同干瘠中RTD的变化都不显著,但分别在重度干旱、极度瘠薄有所下降;山桃在中度、重度、极度干旱比轻度分别减少34.5% (P<0.01)、25.9% (P<0.01)、28.7% (P<0.01),中度瘠薄比轻度瘠薄减少22.5% (P<0.01),重度、极度瘠薄比重度分别增加20.9% (P<0.01)、44.1% (P<0.01);栓皮栎RTD在中度干瘠环境下呈现微小的减小后增加,但是在极度瘠薄下降,在极度干旱和重度瘠薄RTD最大。
2.2.5比根表面积
黄栌随干瘠的加剧,SRA显著减小,而山桃、栓皮栎SRA随着干瘠的加剧呈现出相反的变化。黄栌SRA在中度干旱比轻度减少19.7% (P<0.05),随后增加14.1% (P>0.05),在极度干旱下SRA减少到与中度干旱一致。黄栌SRA随瘠薄的加剧而逐渐减少,中度、重度、极度瘠薄分别比轻度减少15.8% (P>0.05)、21.1% (P<0.05)和28.3% (P<0.01);山桃SRA从轻度干旱到极度干旱呈现出极显著增加的变化,在中度瘠薄环境中山桃SRA极显著减小,随后在重度、极度瘠薄均呈现出极显著增大的变化;栓皮栎SRA在中度干瘠极显著增大,重度干瘠下极显著减小,几乎与轻度的数值相同,在极度干瘠极显著增大。
2.2.6根系密度
黄栌在不同干瘠环境中RAD和RLD变化都不显著,在随着瘠薄的加剧有增大的趋势,均在重度瘠薄中达到最大值,在极度瘠薄呈现出不显著的减小;随着干旱的加剧,山桃RAD、RLD分别呈现出极显著减小、极显著增加的变化,在中度干旱RAD出现最小值,但在极度干旱RLD极显著减小;栓皮栎在随着干旱的加强,RLD极显著减小,中度、重度、极度干旱分别比轻度减少38.4% (P<0.01)、68.5% (P<0.01)、72.9% (P<0.01)。RAD在中度干旱极显著下降,重度干旱极显著增大而极度干旱极显著减小;栓皮栎RAD、RLD均在中度瘠薄中出现最大值,并且从中度瘠薄开始呈现极显著减小的变化。
3 讨论
土壤水分和养分是影响植物生长发育的重要非生物因素之一[17],根系是最先感知土壤水分、养分变化的器官,是连接土壤与植物地上部分之间物质能量交换的重要桥梁,其形态结构决定了植物对土壤资源的获取能力[18]。根系的变化是对环境的适应,目的是为了有效吸收土壤中的水分和养分[19],提高在不利环境中的生存能力,但植物根系性状对干瘠胁迫的适应因树种不同而异。
干旱、瘠薄胁迫下的根系生长并不是单一的被促进或者被抑制,而是与胁迫程度、树种有关[18,20]。连接长度是表征根系在地下空间的分布范围,同时也反映了根系在土壤中的拓展能力[21- 24];根系分支强度是对土壤资源的就地利用能力的指标,可以表示根系分叉数的密集程度[25]。随着土壤环境水分降低,黄栌、山桃和栓皮栎的根连接长度变小,分支强度变大,说明在干旱胁迫下3种树种通过增加侧根数量,形成较多的分叉数[26],减少植株间资源传输距离,保证个体间的资源连接和共享[27],提高土壤水分资源就地利用率。但是山桃、栓皮栎分别在中度、重度干旱其根连接长度显著增大,在极度干旱分支强度减小,增大连接长度减小分支强度扩大根系在土壤的分布范围,可以扩大根系的营养空间[28],随着土壤干旱的加剧根连接长度又显著减小、分支强度显著增大,严重干旱时,植物并没有增加根系深度来获取水分,而是采取小根系来增强竞争力[29],可以在根系扩展范围内对土壤水分原位利用。在瘠薄胁迫中,山桃、栓皮栎根系的根连接长度显著增大,而均在极度瘠薄显著减小,分支强度也呈现逐渐增大的趋势,但是在中度瘠薄中变化不明显,而山桃在重度瘠薄根系分支强度极显著增大,栓皮栎在极度瘠薄极显著增大。根连接长度的增加避免了根系的交叠重复,分支强度增大增加了根系分叉数,有助于根系在瘠薄土壤中有效寻找更大范围养分的同时利用侧根对土壤养分原位利用,增加根连接长度是根系对土壤贫瘠环境的一种适应策略[21,30]。但随着土壤有效养分极度瘠薄,连接长度显著减小、分支强度显著增大,山桃、栓皮栎逐渐发育出相对细密但向周围拓展能力较低的根系结构,这种分枝有利于提高植物对原位养分的利用效率[26]。黄栌在轻度瘠薄的根连接长度最大、分支强度最小,中度瘠薄下根连接长度减小、分支强度增大,随着瘠薄的加剧变化均不大,但是黄栌的平均根连接长度在3个树种中是最大的,说明黄栌在瘠薄胁迫中根系在土壤中延伸范围较广,但在轻度瘠薄的扩展范围最大,而在瘠薄加剧后扩展根系的同时增加了侧根分叉数,提高了对原位土壤养分的利用。
比根长是细根形态与生理功能的一个重要指标,其高低与土壤资源有效性密切相关[28],可以综合反映植物吸收资源的能力与生态适应性[31];比根表面积的值表示与土壤接触机会高低、可以反映植物根系的就地资源利用能力;根组织密度与抗逆相关,代表物质贮存能力、对环境的耐受能力强与弱,是细根生长和吸收能力的重要指标[25]。随着干瘠程度增强,黄栌比根长、比根表面积显著减小,而根组织密度显著增大,说明黄栌在干瘠、瘠薄胁迫下细根较少。干旱环境下,根系生长在浅土区受到抑制,且抑制作用随着降雨的减少而加强[32],而低养分环境有利于延长细根寿命[33- 36],干瘠使黄栌细根生产降低,根系周转相对较慢,减少了细根周转对碳的消耗[26,37]。但是黄栌在干旱、瘠薄胁迫下根组织密度增大,根系物质贮存较好、吸收水分养分能力较强,有效弥补了细根较少的不足。随着干瘠胁迫的加剧,山桃比根长显著减小、比根表面积显著增大,根组织密度显著增大,说明干旱胁迫抑制山桃的根系活性[38]、限制根系生长[39],在土壤水分和养分缺乏环境中根系的根组织密度增大,导致比根长减小[10,40-41]。同时山桃具有较大的根面积有利于最大限度地吸收水分、养分,从而保存较多的稀缺营养,导致根组织密度增大,有利于山桃在严酷的生境中生存[42]。因为根系吸收机制是在比较肥沃的土层中尽可能投入较多的碳水化合物、尽可能多地吸收养分和水分[43],所以山桃根组织密度在轻度干旱、轻度瘠薄时较大。栓皮栎比根长和比表面积随土壤干旱、瘠薄胁迫增加而增加,表明土壤水分、养分胁迫诱导栓皮栎 产生更多数量、活力较强的新生细根,从而使比表面积、比根长增加[44]。这一现象说明,在土壤水分、养分胁迫下,栓皮栎并不是被动忍受逆境胁迫,而是主动调节其根系生长以提高对水分和养分的吸收能力,减缓逆境伤害。栓皮栎根组织密度也随着干瘠加剧而显著增加,表明栓皮栎在干瘠胁迫下根系生长和吸收能力也越来越强,有利于提高物质贮存能力、对环境的耐受能力。而且栓皮栎的比根长、比根表面积都比较大,意味着栓皮栎对养分和水分等资源有着很强的竞争力,这就使植物进行快速生长有了可能[42]。
根长密度表示单位体积土壤中根系总长度,反映根系数量的多少[45],对根系吸水和营养物质影响较大[46]。根表面积密度是单位土壤体积的总根表面积,是研究水分吸收或养分吸收的重要参数之一[47]。黄栌根长密度、根表面积密度在不同干瘠环境中变化较小,而且黄栌的根长密度、根表面积密度较小,虽然在干瘠立地上黄栌细根不多,但是粗根或老根则可以更好地保持植物取得的土壤水分和养分[48],以提高黄栌耐干瘠能力。山桃根表面密度不同干瘠环境中变化也较小,说明干瘠、瘠薄对山桃根表面积密度的影响不大,山桃利用较大根表面积贮存营养物质,而根长密度随干瘠的加剧显著增大,说明干旱胁迫都刺激了山桃根系的生长,在干旱的环境条件下,根系为了寻找更多水源,增加根长密度[47],但是在极度干旱时,山桃根长密度显著减小,说明当土壤干旱超过一定的阈值后,树木根系将逐步丧失其活力和功能[38]。栓皮栎根长密度、根表面积密度随干瘠加剧而减小,但在中度瘠薄中显著增大并达到最大值,说明栓皮栎在中度瘠薄土壤环境中,为获得更多的土壤养分而采取主动适应的生态对策,投入大量的光合产物至根系以刺激根系生长[49],通过增加根系与土壤的接触面积而获取更多的土壤养分资源。但是,细根周转对干旱胁迫的响应较为敏感[50],林木常因产生的细根量大、根系周转速度较快而付出根系损失较大的代价[51],所以为了减少细根周转对碳的消耗,栓皮栎减小了对细根的生长。栓皮栎的根长密度、根表面积密度在干瘠环境中的均值较大,说明栓皮栎的细根数量比黄栌、山桃多,吸收土壤水分、养分能力强,在干瘠环境中,栓皮栎是速生树种,而黄栌、山桃是慢生树种。总之,植物对干瘠环境的变化都有一定的响应能力及适应对策,且在漫长的进化过程中已经形成了一种自我调节机制,能够通过自身根系形态变化来适应不断变化的环境,但不同树种对干瘠环境变化的适应策略不同,具有明显差异[52]。
4 结论
(1) 干旱、瘠薄同时存在,且均对树木根功能性状产生影响。3种树种根功能性状在不同干瘠等级下发生显著变化,说明在干瘠环境下植物主动调节根系构型适应干瘠立地。此外,而且不同树种同一性状的变化也各不相同,植物根系在适应极端干旱环境中,通过改变根系形态和根系构型,表现出显著的差异。
(2) 在干瘠胁迫下,黄栌、山桃、栓皮栎均通过增加侧根分叉数、增大根组织密度和扩展根系在土壤中的分布范围来提高抗干瘠能力。黄栌比根长、比根表面积在干瘠中显著减小,根系萌蘖性强;山桃比根长显著减小、比根表面积显著增大;栓皮栎的比根长、比根表面积在干瘠胁迫中比较大,意味着栓皮栎对养分和水分等资源有着很强的竞争力。
(3) 植物对干瘠环境的变化都有一定的响应能力及适应对策,且在漫长的进化过程中已经形成了一种自我调节机制,能够通过自身的形态变化来适应不断变化的环境。不同物种之间对干瘠环境变化的适应途径不同,通过权衡碳分配、碳消耗采取了相应的生态适应策略,不同树种间具有明显的差异。
猜你喜欢
动漫界·幼教365(小班)(2021年6期)2021-07-21 15:42:20
小学阅读指南·低年级版(2020年9期)2020-10-12 02:43:08
南方农业·下旬(2020年1期)2020-06-01 07:47:32
中学生百科·悦青春(2020年6期)2020-01-06 07:51:01
北京广播电视报(2019年19期)2019-07-12 13:24:47
现代园艺(2019年24期)2019-01-07 01:19:36
现代农业科技(2009年21期)2018-09-03 00:00:00
北方文学(2017年2期)2017-08-04 05:31:43
北方文学·中旬(2017年1期)2017-03-15 15:52:17
农村百事通(2017年18期)2017-02-08 11:23:54
