
自然覆雪对5个不同秋眠级紫花苜蓿越冬率及抗寒性的影响
2021-12-23 09:10:06苏力合张凡凡王旭哲贺婷婷马春晖
新疆农业科学 2021年11期
苏力合,张凡凡,王旭哲,宋 磊,俞 雪,贺婷婷,马春晖
(石河子大学动物科技学院,新疆石河子 832003)
0 引 言
【研究意义】紫花苜蓿(MedicagosativaL.)是多年生豆科牧草,其产草量高、适口性好、再生能力强[1]。目前我国优质苜蓿的产区集中在东北、西北地区,种植面积约占全国89.8%[2]。新疆是我国主要苜蓿种植区和草食畜牧业主产区,新疆地区冬季寒冷漫长,种植紫花苜蓿面临越冬问题,冬季气候,土壤环境,品种耐寒性等[3]。苜蓿引种需要根据其特有的秋眠性(Fall dormancy)特征合理进行栽培生产,以满足不同生态区域的需要[4]。【前人研究进展】抗冻性是受多基因控制的复杂性状[5],且不同地区的田间环境条件多变,传统的育种方法在抗冻性方面只取得了适度的改善[6-7]。【本研究切入点】除了苜蓿自身遗传的抗寒性之外,田间管理措施也会影响苜蓿的越冬率及抗寒性,越冬前的田间灌水、留茬高度、覆盖麦草和积雪等在不同程度上都可提高苜蓿的越冬率[8-10]。研究新疆石河子地区不同秋眠级紫花苜蓿在自然覆雪下越冬率及抗寒生理指标的表现。【拟解决的关键问题】选择秋眠型(1、3级)、半秋眠型(5级)及非秋眠型(7、9级)紫花苜蓿,从秋季至翌年春季无积雪和自然降雪条件下,分析苜蓿根系可溶性糖(WSC)、可溶性蛋白(SP)、游离脯氨酸(Pro)、丙二醛(MDA)、超氧化物歧化酶(SOD)和过氧化物酶(POD)含量以及越冬率。为紫花苜蓿品种合理选择和高效生产提供理论支撑。
1 材料与方法
1.1 材 料
试验地位于新疆石河子大学农试验场牧草试验站(N 44°20′,E88°30′,海拔450.8 m),地处新疆北部,地势平坦;属于温带大陆性气候,冬季长而严寒,夏季短而炎热。年均气温2~15℃,无霜期为168~171 d,年降水量为125.0~207.7 mm,年日照时数为2 721~2 818 h。试验田前茬作物为棉花。表1
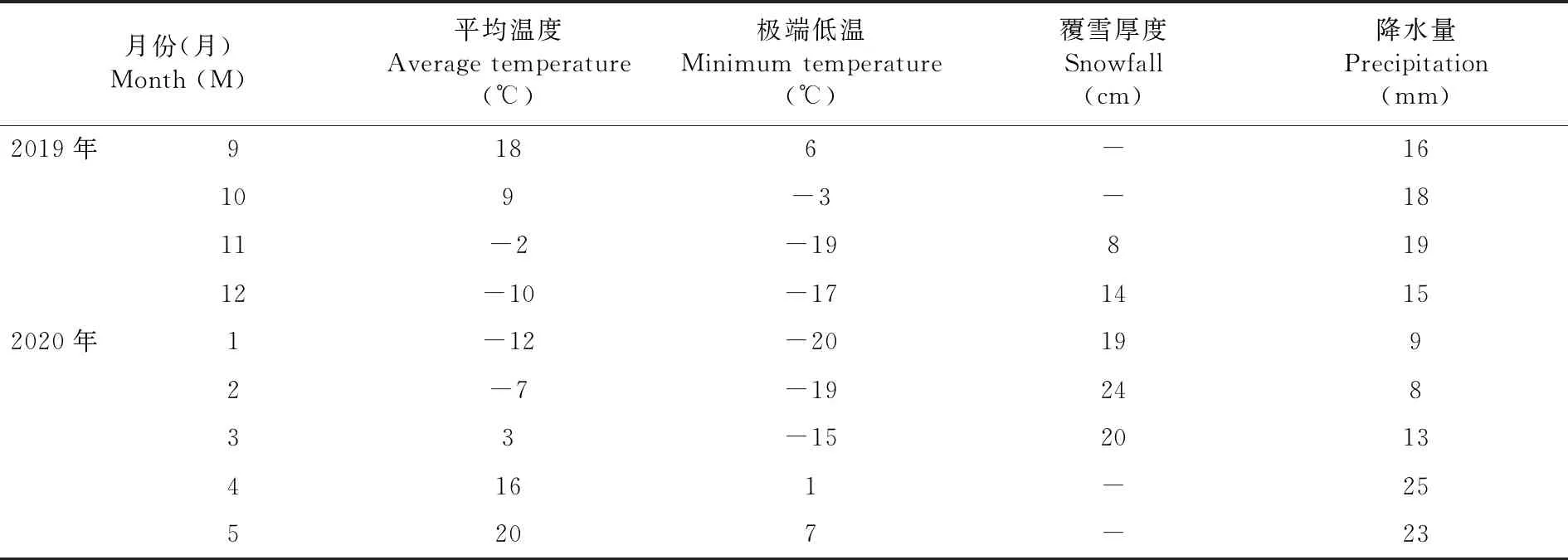
表1 试验年(2019年9月~2020年5月)的天气状况
1.2 方 法
1.2.1 试验设计
供试紫花苜蓿品种共5个。播种时间为2018年4月,设计小区面积50 m2(10 m×5 m),每个品种共3个小区,相邻2小区间隔0.5 m的保护行。种植方式采用穴播,播种深度1~2 cm,行距80 cm,株距60 cm。采取滴灌方式灌溉,以水井为主要水源。表2

表2 紫花苜蓿品种及来源
设计自然覆雪与不覆雪处理。每个品种选取小区面积10 m2(5 m×2 m),每次降雪后及时将雪打扫干净维持不覆雪状态。于2019年9月至2020年5月观测冬季苜蓿情况。
于2019年11月3日至2020年3月30日,采用多点式土壤温度记录仪(i500-E8T,玉环智拓仪器科技有限公司)测定覆雪与不覆雪下不同土层深度(1和20 cm)和苜蓿冠层处(高于地面5 cm处)的温度,土壤温度记录仪测量时间间隔设置为10 min,每个土层放置1个温度探头。
1.2.2 测定指标
分别于2019年9月8日、10月26日、11月24日、12月29日和2020年1月12日、5月12日从试验田随机挖取供试材料10 cm长的根及根茎,清洗干净放入液氮罐速冻带回实验室备测。
可溶性糖(water soluble carbon,WSC)采用蒽酮比色法;可溶性蛋白(soluble protein,SP)采用考马斯亮蓝G-250法;丙二醛(malondialdehyde,MDA)采用硫代巴比妥酸比色法;游离脯氨酸(proline,Pro)采用酸性茚三酮法;超氧化物歧化酶(superoxide dismutase,SOD)活性采用NBT光化学还原法;过氧化物酶(peroxidase,POD)活性采用愈创木酚法。
于2019年9月最后一茬刈割后在种植各苜蓿品种的小区内选择观测每个紫花苜蓿品种样方内(1 m2)越冬前和返青后,在有无覆雪条件下的植株数,重复3 次,越冬率计算方法为,越冬率=存活总株数/植株总数×100%。
1.3 数据处理
用Ecxel 2010软件对原始数据进行初步整理,用SPSS21.0软件进行单因素方差分析,多重比较采用Ducan法。应用隶属函数法进行紫花苜蓿抗寒性的综合评价。其中,与抗寒性呈正相关的指标(SS、SP、Pro、SOD、POD和WSR)采用公式Fijk+=(Xijk-Xmin)/(Xmax-Xmin)计算,与抗寒性呈负相关的指标(MDA)采用公式Fijk-=1-Fijk+计算,其中Fijk为第i个品种第j个时间第k项指标的隶属度,Xijk为第i个品种第j个时间第k项指标测定值,Xmax和Xmin为所有参试材料中第k项指标的最大值和最小值。
采用隶属函数值法对5个不同秋眠级,6个采样时间的苜蓿根系SS、SP、MDA、Pro、SOD、POD含量和WSR7个指标进行抗寒性综合评价。综合评价分越高,紫花苜蓿抗寒性和越冬率越好,反之则差。
2 结果与分析
2.1 覆雪与不覆雪处理对紫花苜蓿根冠处、土壤表层(1~20 cm)温度的影响
研究表明,无积雪与自然降雪下紫花苜蓿根冠处及土壤表层每日平均温度差异很大。无积雪覆盖时,暴露在空气中的苜蓿根冠处温度上下波动幅度很大,其中在1月11日出现最低温度(-20.74℃),在3月10日出现最高温度(4.98℃),温度变化幅度为25.72℃,土壤表层20 cm处温度在-5~-10℃波动,其中在12月28日出现最低温度(-12.07℃),在3月10日出现最高温度(5.30℃),温度变化幅度为17.37℃。田间从2019年11月18日至2020年3月10日间有雪覆盖,苜蓿根冠处温度上下波动幅度较无积雪条件小,其中在1月10日出现最低温度(-7.89℃),在3月6日出现最高温度(3.67℃),温度变化幅度为11.56℃,远远小于无雪覆盖下的苜蓿根冠处及土壤表层温度,而土壤表层20 cm处温度在大部分测量期间都保持在-3℃以上,仅在1月10日出现最低温度(-4.20℃),在3月6日出现最高温度(3.44℃),温度变化幅度为7.64℃。图1

注:1A:无积雪覆盖;1B:自然降雪
2.2 覆雪与不覆雪下各秋眠级紫花苜蓿抗寒生理指标的变化
2.2.1 根系WSC含量变化
研究表明,各秋眠级紫花苜蓿根系WSC含量在覆雪下整体呈先升后降的趋势,不覆雪下除了WL903呈先升后降的趋势外,其他品种呈反复升降趋势。2019年10月冷驯化期,各秋眠级苜蓿WSC含量升高,显著高于9月未降温期(P<0.05)。11月田间形成有效积雪后,各品种WSC含量表现不同,无积雪覆盖下,除了WL903品种WSC含量继续上升外,肇东、中苜2号和渝苜1号WSC含量较10月显著下降,但仍高于9月含量(P<0.05);自然降雪下,各苜蓿品种WSC含量持续上升,除了中苜2号WSC含量较10月不显著外,其余各品种WSC含量显著高于10月(P<0.05)。12月及2020年1月田间稳定冻结期期间,各苜蓿品种在覆雪下的WSC含量均高于不覆雪苜蓿品种(除1月中苜2号外)。2020年5月,各苜蓿品种WSC含量显著低于冬季(P<0.05),且在覆雪下的苜蓿品种WSC含量仍高于不覆雪品种。表3

表3 紫花苜蓿根系可溶性糖含量变化
2.2.2 根系SP含量变化
研究表明,不同秋眠级紫花苜蓿根系SP的含量随气温的下降呈先升高后降低的趋势。2019年10月冷驯化期,各秋眠级苜蓿SP含量缓慢积累,肇东、中苜2号与渝苜1号的SP含量显著高于9月(P<0.05)。2019年11月随着寒冷加剧以及降雪,各秋眠级苜蓿SP含量显著增加并达到最大值(P<0.05),覆雪条件下Archer含量最高为18.72 mg/g,各秋眠级苜蓿SP含量在覆雪状态下整体高于不覆雪的苜蓿(除中苜2号外)。12月至2020年1月各秋眠级苜蓿SP含量开始降低,但仍高于未降温期,其中覆雪下的中苜2号下降幅度最小,各秋眠级苜蓿SP含量在覆雪状态下整体高于不覆雪的苜蓿。2020年5月,各品种SP含量降至最低值,仅无覆雪肇东和覆雪WL903与2019年9月含量差异显著(P<0.05)。表4
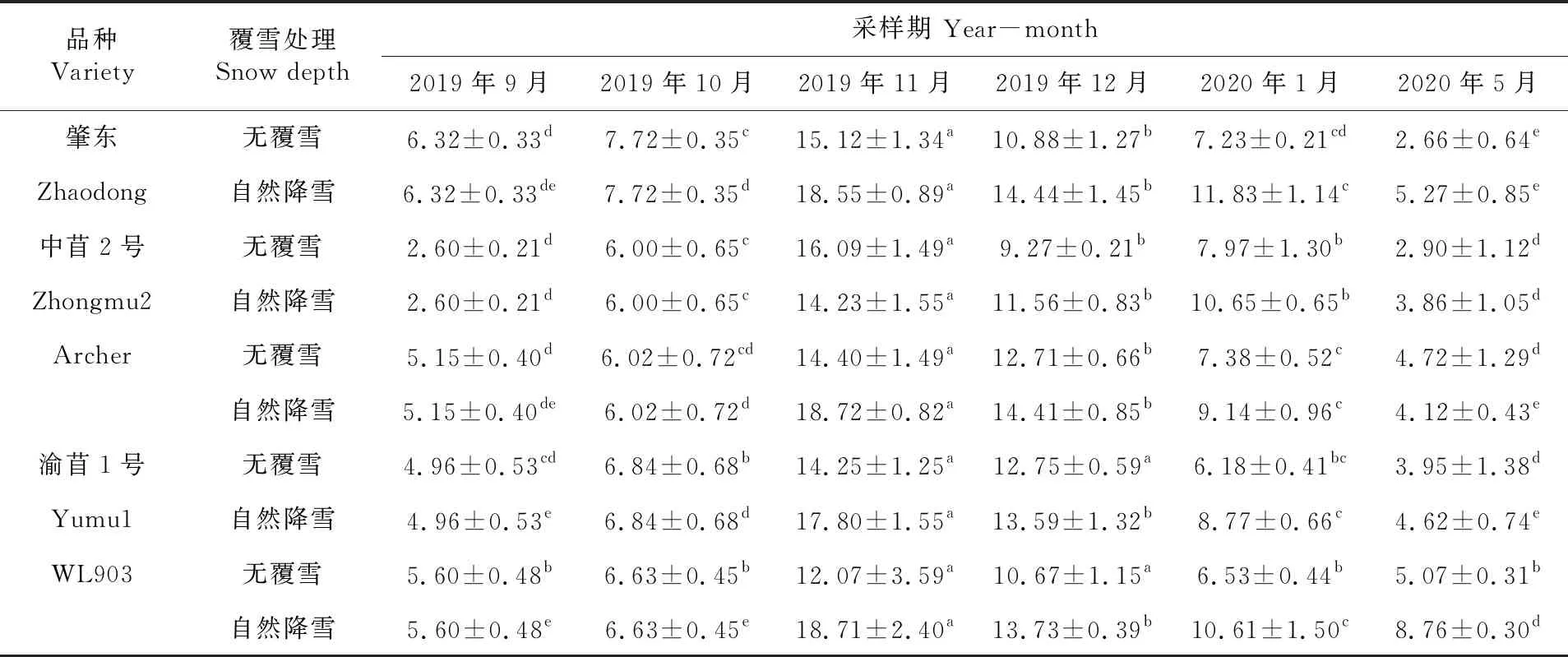
表4 紫花苜蓿根系可溶性蛋白含量变化
2.2.3 根系游离Pro含量变化
研究表明,各秋眠级苜蓿根系游离Pro含量整体呈波浪趋势,且在覆雪状态下各秋眠级苜蓿游离Pro含量整体高于不覆雪的苜蓿。2019年10月冷驯化期,各秋眠级苜蓿其游离Pro含量显著增加(P<0.05)。11月田间形成有效积雪后,各品种Pro含量表现不同,无积雪覆盖下,肇东、Archer和WL903显著上升(P<0.05),中苜2号较10月小幅度下降但差异不显著(P>0.05),渝苜1号较10月小幅度上升但差异不显著(P>0.05);覆雪条件下,各品种游离Pro含量均达到最大值,显著高于其他各时期(P<0.05),其中中苜2号含量最高为1 384.43 μg/g。2019年12月至2020年1月在无积雪覆盖下,肇东、Archer及WL903品种较2019年11月其Pro含量呈先降低后上升的趋势,中苜2号持续升高,渝苜1号先上升后降低,且各品种在不同月份含量差异显著(P<0.05);覆雪条件下,仅中苜2号持续降低显著低于2019年11月(P<0.05),其余各品种呈先降低后上升的趋势,其中WL903在2020年1月含量最高为1 192.00 μg/g。表5
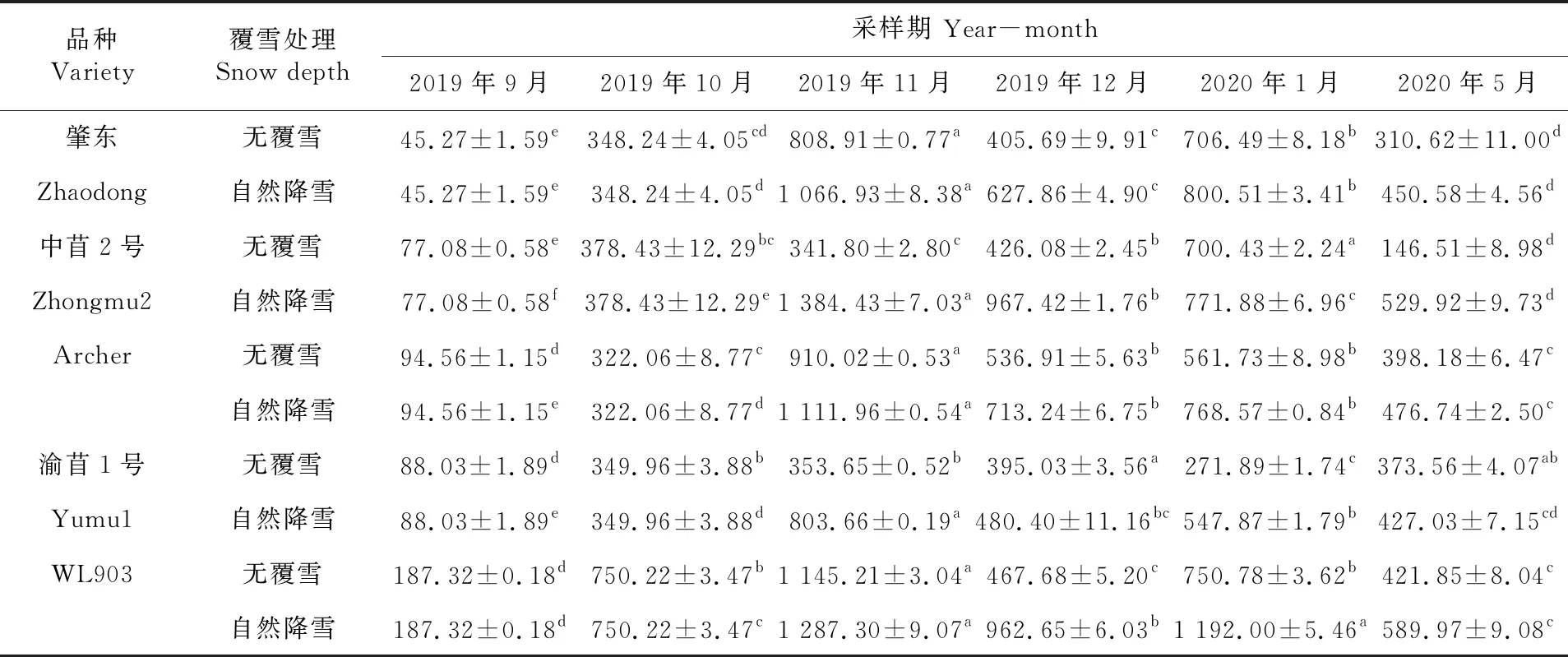
表5 紫花苜蓿根系游离脯氨酸含量变化
2.2.4 根系MDA含量变化
研究表明,各秋眠级苜蓿根系MDA含量整体呈先下降后上升再下降的趋势。2019年10月冷驯化期,各秋眠级苜蓿根系MDA含量缓慢降低,除WL903品种外其余品种MDA含量显著低于9月(P<0.05),但随11月冷胁迫期到来及降雪,各秋眠级苜蓿根系MDA含量变化各不相同,肇东MDA含量在无覆雪及覆雪下持续降低,中苜2号及Archer的MDA含量在无覆雪及覆雪下升高,渝苜1号和WL903的MDA含量在无覆雪下升高,覆雪下降低。12月各品种MDA含量均升至最大值,显著高于各时期(P<0.05),其中WL903在不覆雪和覆雪下根系MDA含量最高。2020年1月冷冻稳定期,各苜蓿经过抗寒胁迫后,其MDA含量开始降低,其中肇东在覆雪下含量最低,渝苜1号在不覆雪下含量最高。覆雪下各秋眠级苜蓿MDA含量均低于不覆雪苜蓿。表6
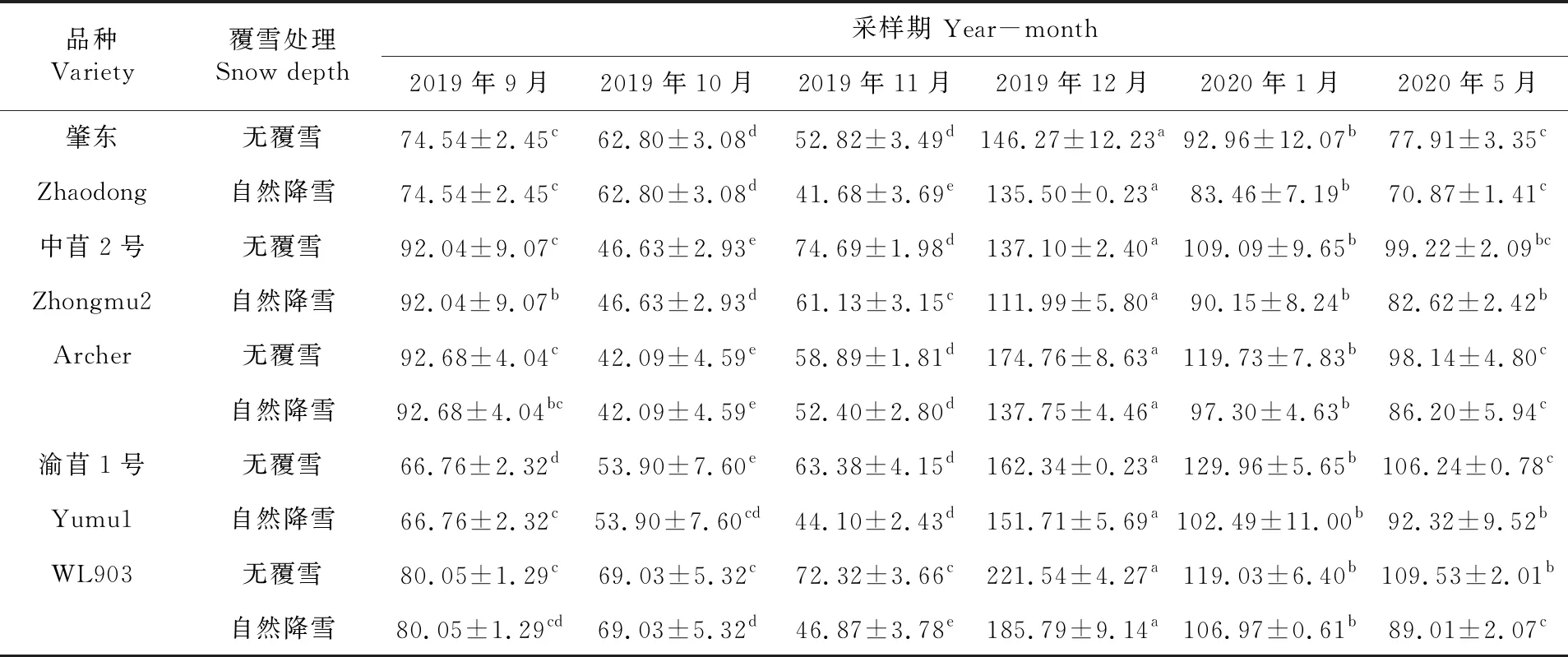
表6 紫花苜蓿根系丙二醛含量变化
2.2.5 根系SOD含量变化
研究表明,各秋眠级苜蓿根系SOD含量变化与游离Pro含量变化相似,表现为升高→降低→再升高→再降低,且在覆雪条件下各秋眠级苜蓿SOD含量整体高于不覆雪苜蓿(除11月WL903外)。2019年10月冷驯化期,紫花苜蓿感应低温SOD含量急剧上升,其中肇东SOD含量上升幅度最大。11~12月持续的冷胁迫以及降雪,各秋眠级苜蓿品种SOD含量开始降低,不覆雪条件下,肇东、中苜2号和WL903在11月SOD含量降至最低值,Archer和渝苜1号在12月降至最低值,显著低于其余时期(P<0.05);覆雪条件下,中苜2号、渝苜1号和WL903在11月降至最低值,肇东和Archer在12月降至最低值,显著低于其余时期(P<0.05)。2020年1月冷冻冻结期,各秋眠级苜蓿在经过长时间的抗寒锻炼后其根系SOD含量又开始上升,不覆雪条件下Archer品种SOD含量显著高于其余各时期(P<0.05);覆雪条件下,各秋眠级紫花苜蓿的SOD含量均在2020年1月达到最大值,显著高于其余各时期(P<0.05)。表7
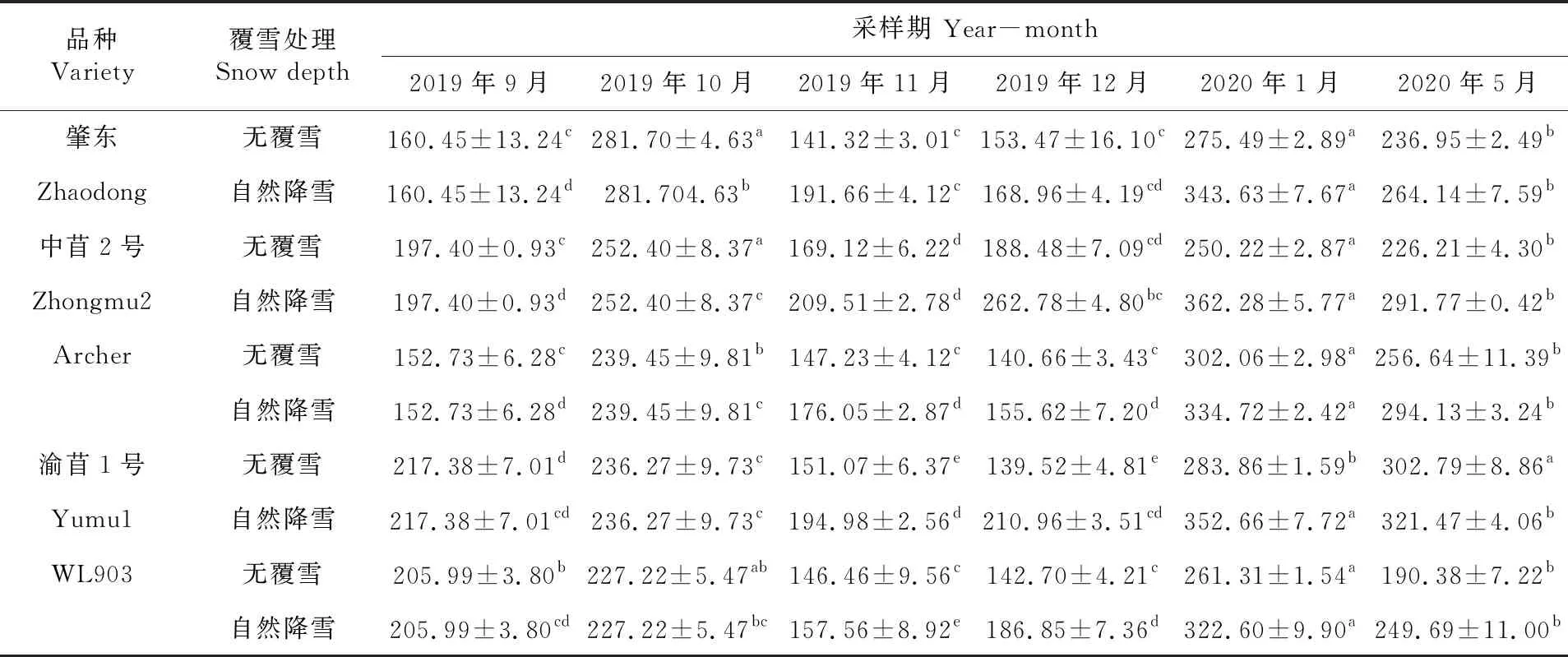
表7 紫花苜蓿根系超氧化物歧化酶含量变化
2.2.6 根系POD含量变化
研究表明,不同秋眠级紫花苜蓿根系POD含量随温度的降低呈先升高后降低的趋势。2019年10月冷驯化期,各秋眠级苜蓿POD含量逐渐积累。11月随着寒冷加剧,各秋眠级苜蓿POD含量在覆雪及不覆雪条件下都大幅度上升,显著高于10月(P<0.05),其中覆雪条件下的中苜2号上升幅度最大。12月各秋眠级紫花苜蓿的POD含量在覆雪及不覆雪条件下均达到最大值,显著高于其余时期(P<0.05),其中Archer在覆雪条件下含量最高为561.15[μ/(g·min)]。2020年1月开始各秋眠级苜蓿POD含量开始下降(不覆雪WL903下降幅度最大),但各品种仍显著高于未冷驯化期的含量(P<0.05),且各秋眠级苜蓿POD含量在覆雪状态下整体高于不覆雪的紫花苜蓿。表8
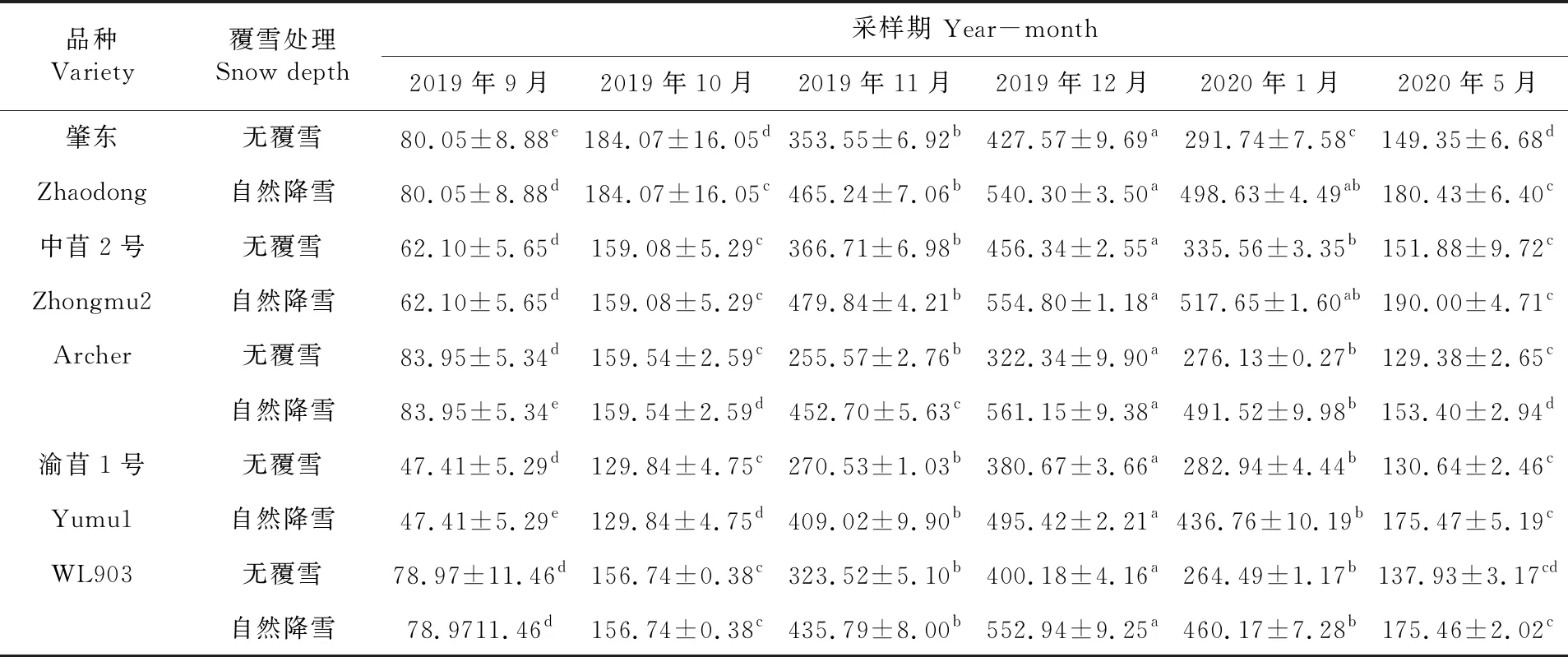
表8 紫花苜蓿根系过氧化物酶含量变化
2.3 覆雪与不覆雪条件对各秋眠级紫花苜蓿越冬率的影响
研究表明,各秋眠级苜蓿品种翌年均能返青。无积雪覆盖下,Archer、渝苜1号和WL903越冬率分别为77.31%、68.79%和66.99%,显著低于肇东和中苜2号(P<0.05)。自然降雪下,Archer、渝苜1号和WL903越冬率分别为93.33%、86.67%和73.33%,其越冬率较无覆雪显著提高(P<0.05)。覆雪下WL903越冬率显著低于其余品种(P<0.05)。图2
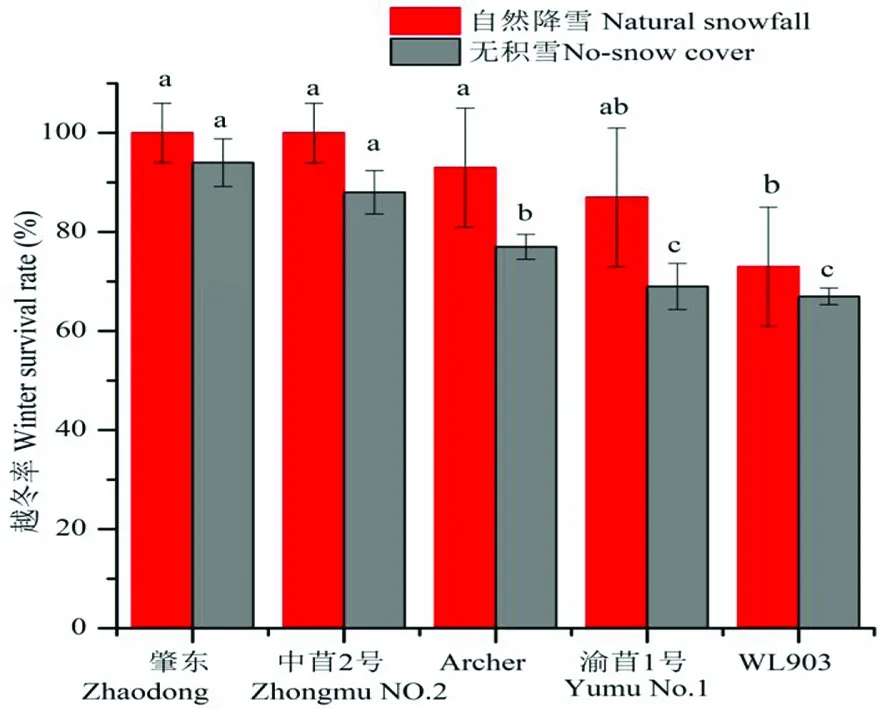
注:不同小写字母表示差异显著(P<0.05)
2.4 覆雪与不覆雪条件对各秋眠级紫花苜蓿抗寒性和越冬率的综合评价
研究表明,在无覆雪条件下,不同秋眠级紫花苜蓿抗寒性综合大小排序为肇东(1级)>中苜2号(3级)>Archer(5级)>WL903(9级)>渝苜1号(7级);自然降雪条件下:肇东(1级)>中苜2号(3级)>Archer(5级)>WL903(9级)>渝苜1号(7级)。表9
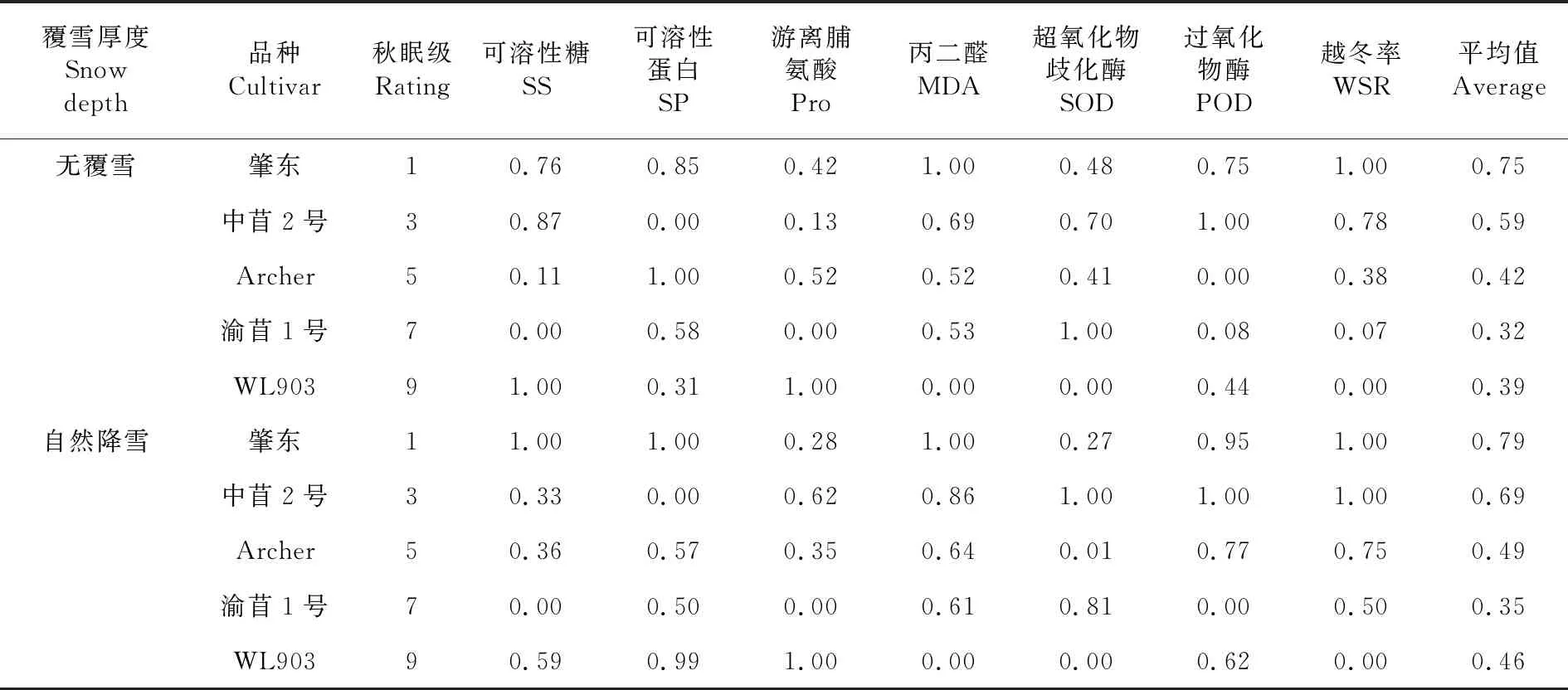
表9 覆雪与不覆雪条件下各秋眠级紫花苜蓿抗寒性和越冬率的隶属函数
3 讨 论
3.1 覆雪与不覆雪条件对各秋眠级紫花苜蓿根茎土层温度及越冬率的影响
无积雪覆盖时,苜蓿秋眠级和越冬率呈现负相关,高秋眠苜蓿越冬率较低主要由于秋季继续生长而未能在根系存储较多养分[11],冬季遭受冷冻胁迫耗尽根系养分,使得根系不能抵御冻害(极端气温-20.74℃)。在有积雪覆盖情况下,秋眠级5和7级苜蓿表现出较好的越冬率(与1、3级差异不显著),越冬率高的原因是雪的低导热性和高反射性能提高其覆盖下的地表温度[12],覆雪下相对恒定的根茎及土壤表层温度对紫花苜蓿造成较低的伤害,李如来[9]的研究结论相同。岳亚飞等[8]研究结果表明,秋眠级为10的品种在无积雪覆盖下翌年未能全部返青,而试验中秋眠级为9的品种在不覆雪下返青率为66.91%,造成差异的原因可能是其试验年天气温度(最低为-31℃)的不同及苜蓿自身遗传差异,试验年苜蓿根茎温度最低为-20.74℃,高于其苜蓿根茎温度,因而高秋眠级苜蓿部分能返青。
3.2 覆雪与不覆雪条件对各秋眠级紫花苜蓿抗寒性的影响
植物在遭受低温胁迫时,体内的渗透调节物质、膜透性以及酶的活性都会影响植物的抗寒性[13,14]。研究结果中,WSC、SP和Pro含量在2019年9月最低,10月冷驯化期显著上升(P<0.05),11和12月达到峰值,Castonguay等[6]认为,此时的苜蓿通常达到最大抗逆性,随后2020年1月开始下降,且覆雪下WSC、SP和Pro含量普遍高于不覆雪条件。冷驯化期秋眠型苜蓿(1、3级)的WSC含量最高,可能是由于秋眠型苜蓿生长停止的时间早,根、冠中早期积累的WSC含量也越高[15]。Cunningham等[16]研究认为WSC可作为衡量苜蓿抗寒性强弱的最有力指标,非耐寒和冬耐寒品种之间的最大耐寒性水平与苜蓿积累WSC的能力有关。除了苜蓿自身适应寒冷之外,冬季覆雪也能显著提高苜蓿的抗寒性,这点在WSC、SP和Pro含量覆雪下普遍比无积雪含量高可看出,原因是因为雪被使苜蓿根系大多数时间在相对恒定的温度下越冬,而无积雪覆盖时紫花苜蓿根冠处最低温度可达-20℃,持续低温使苜蓿根系发生冻害,随之渗透调节物质含量也降低,影响其抗冻性。试验中Pro含量在整个冬季及翌年春季都保持较高的水平,与申晓慧[17],张璐[18]等研究结果一致,其中WL903游离Pro含量在温度最低时期(1月)表现出与其他品种的差异性,其在不覆雪及覆雪下的含量远高于其余品种,表明苜蓿抗寒性受多重因素影响,不能单一的用秋眠级或单个指标衡量其抗寒性强弱[19]。 翌年春季天气回暖,WSC、SP和Pro含量与冬季相比明显下降,是由于WSC作为春季再生能源被越冬芽萌动消耗,此时苜蓿根系营养由库端变成源端[20]。
试验中各秋眠级苜蓿MDA含量呈先降低后上升再降低的趋势,这与陶雅[21],张仲鹃[22]等研究结果相一致。先降低是因为冷驯化引发苜蓿WSC、SP和Pro的大量积累,有效防止了活性氧对膜脂过氧化作用[17],另外SOD和POD活性在经过冷驯化后增强并协同作用清除细胞内的活性氧,起到保护细胞的作用[12],但随着气温持续降低,苜蓿根系渗透物质含量及酶活性降低,苜蓿抵抗寒冷减弱引发MDA含量升高。翌年1月紫花苜蓿根系酶的活性又开始增强,MDA含量开始降低,此间SOD和POD酶通过清除细胞内的活性氧自由基来降低膜脂过氧化程度。试验中高秋眠级苜蓿在覆雪时MDA含量下降,可能是积雪削弱了苜蓿的秋眠性[8],提高了高秋眠级苜蓿抗冻能力[23]。无积雪覆盖下的苜蓿MDA含量均高于覆雪下的苜蓿,覆雪可在一定程度上保护苜蓿根系免受冻害。
4 结 论
覆雪下各品种的平均值均高于无积雪下的苜蓿,覆雪能显著提高高秋眠级苜蓿的越冬率及抗寒性,秋眠型苜蓿的抗寒性较非秋眠型好。
相比于不覆雪下的紫花苜蓿根冠处最低温度(-20.74℃),覆雪可以提高紫花苜蓿根冠处(-7.89℃)及土壤表层20 cm的温度,保护紫花苜蓿根茎及根系使其安全越冬。紫花苜蓿越冬率表现为秋眠型>半秋眠型>非秋眠型,覆雪下半秋眠型(5级)以及非秋眠型(7级)苜蓿越冬率升高至85%以上,覆雪能显著提高紫花苜蓿的越冬率。根系保护物质含量随温度的降低而升高,覆雪下各秋眠级紫花苜蓿根系可溶性糖、可溶性蛋白和游离脯氨酸含量,以及超氧化物歧化酶和过氧化物酶活性高于不覆雪处理,丙二醛含量低于不覆雪处理。在有积雪覆盖下秋眠级1和3级的苜蓿抗寒性表现最好。
猜你喜欢
现代妇女(2024年4期)2024-04-29 06:59:03
福建文学(2019年12期)2019-08-06 14:59:46
黑龙江粮食(2018年8期)2018-10-18 10:09:24
炎黄地理(2017年10期)2018-01-31 02:15:05
少年文艺·开心阅读作文(2017年1期)2017-02-24 03:05:29
安徽农业科学(2016年32期)2016-12-14 09:39:08
安徽农业科学(2016年32期)2016-12-14 09:39:07
高原山地气象研究(2016年1期)2016-11-10 06:05:53
新疆农垦科技(2016年2期)2016-08-21 13:50:26
中国果菜(2016年9期)2016-03-01 01:28:40
