
果蝇基因剂量补偿的实现机制*
2021-12-22 10:12:00张露丹王瑞雪
生物学通报 2021年3期
张露丹 王瑞雪 孙 林
(北京师范大学生命科学学院 北京 100875)
基因剂量补偿是指染色体虽有剂量变化,但其基因的表达量与正常二倍体的表达量是近似相等的。该现象最早在果蝇白眼等位基因中被发现,在XY 型性染色体性别决定生物中,X 染色体基因剂量补偿已被广泛研究。在果蝇中,雄性个体通过2 倍上调转录以平衡单条染色体对转录丰度的影响。近年来,有关果蝇基因剂量补偿的研究已有极大进展,但其分子机制一直存在争议,研究者主要持有2 种观点:平衡模型和激活模型。
1 平衡模型
平衡模型的主要内容是:雄性个体X 染色体剂量不一致所导致的反式剂量效应往往会使基因组表达量上调约2 倍。同时,MSL 复合体会将组蛋白修饰物定位于X 染色体上,以避免常染色体上发生基因剂量补偿,同时防止X 染色体上的基因过 表达[1]。
1.1 反式剂量效应 在遗传学研究过程中,研究者发现,单个染色体拷贝数的改变比整个染色体倍性变化对个体表型的影响更大,并据此提出了基因平衡假说(gene balance hypothesis),以解释该现象的分子基础[2]。果蝇的非整倍体样本表现出对于酶活性和蛋白质、RNA 水平表达的调节,而不是倍性的平衡变化,这种调节通常与不同染色体的剂量呈正相关或负相关。由于基因剂量改变而引起的基因表达量的变化称为基因剂量效应,包括正向剂量效应和反式剂量效应2 种影响[3](图1)。基因组内的基因剂量补偿正是通过这2 种基因剂量效应相互作用实现的。正向剂量效应指的是基因表达量与基因剂量变化呈现正相关;而反式剂量效应指的是基因表达量与基因(某染色体片段或完整的染色体)剂量变化呈现负相关。反式剂量效应通过调控转录因子和级联效应相关的基因,在非整倍体全基因组内产生广泛影响。这使得拷贝数变化的染色体基因能通过2 种剂量效应维持与正常拷贝数中一致的表达水平,而其他染色体上的基因表达水平出现与非整倍体数量变化相反的变化趋势。当染色体数目从二倍体中的2 条增加至三体个体中的3 条,基因正向剂量效应会随着染色体剂量的增加变为150%,而与此同时,产生的反式剂量效应会作用于全基因组内,并将变化染色体片段上的基因及其他正常染色体基因表达量均下调为67%[4]。数量发生变化的染色体上的基因同时受到正向剂量效应和反式剂量效应共同调控,最终可维持与正常二倍体一致的表达水平,即实现剂量补偿现象。但其他染色体的基因表达量在未发生正向剂量效应的变化的同时,仅受到反式剂量效应的调控,因此,它们的表达水平下降至67%(图1)。
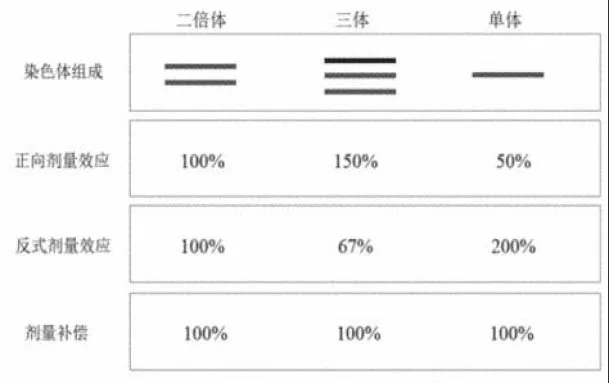
图1 三体和单体中的剂量补偿
1.2 MSL 复合体 雄性特异性致死(male-specific lethal,MSL)复合体是富集在雄性果蝇X 染色体周围的一种蛋白质复合体,其组成成分包括雄性特异性致死蛋白1、2、3 (MSL1、MSL2、MSL3),组蛋白乙酰化酶(MOF),RNA/DNA 解旋酶(MLE)和磷酸激酶(JIL-1)6 种蛋白质亚基及2 种非编码RNA(roX1 和roX2)[5]。其中MOF 负责乙酰化组蛋白H4 的16 位的赖氨酸残基(H4K16Ac),使组蛋白乙酰化修饰在雄性X 染色体上富集。由于MSL 复合物可精准结合在X 染色体的特定区域,组蛋白修饰因子MOF 和JIL1 激酶远离常染色体,从而消除了在常染色体上可能发生的反式剂量效应[5]。此过程是对正常雄性中发生的基因组失衡的一种反应,它将确保X 染色体表达量得到补偿,而常染色体不会过度表达。但这并不意味着MOF 富集导致的高乙酰化引起了单个X 染色体的上调。性别致死基因(sxl)的表达决定了MSL 的性别相关性[5]。在雌性中,sxl早期产物对晚期转录物进行“自我”剪接,继续表达,并发生级联剪接决定性别分化。SXL 蛋白与MSL2 蛋白的mRNA结合,并阻断其在雌性中的表达。雄性中SXL 蛋白的缺失使得msl2基因得以表达,而MSL2 蛋白正是复合体组装必不可少的组分。
研究者通过敲除雌性果蝇X 染色体上MSL2和SXL 的结合位点,使MSL 复合物可组装并定位到雌性的X 染色体上。结果表明,MOF 在X 染色体上富集,H4K16Ac 组蛋白修饰水平随之增加,但X 染色体连锁的大部分基因表达量并未显著增加[5]。相反,许多常染色体基因的表达量是显著下调的。实际上,MSL 复合体负责进行适度的剂量补偿,并发挥一定的抑制作用以防止基因过表达。其机制可能是随着X 染色体上组蛋白修饰物的增加,其染色质结构发生变化,导致染色质重塑[1]。异染色质蛋白1(HP1)在雄性X 染色体上富集,但其与剂量补偿效应的相关性尚待探索[6]。此外,遗传筛选表明MSL 复合物并不是限制剂量补偿的唯一因素,例如,Overcompensating male(ocm)被认为是MSL 复合体的限制因子,其雄性缺失突变体死亡是X 连锁基因过度表达的结果[7]。这表明了ocm在剂量补偿中起约束基因过表达的作用,并且研究者发现ocm和MSL 复合物可相互抑制,这进一步支持了该假设,但ocm调节MSL 复合物活性的机制尚不清楚。
2 激活模型
基于雄性果蝇基因表达水平上调至2 倍以补偿两性间基因表达这一结果,一些研究者提出了果蝇基因剂量补偿的另一种实现机制——激活模型。在用于解释果蝇X 染色体超活化的过程中,MSL 复合体扮演着关键角色。在激活模型中,MSL复合体可特异性地结合于雄性X 染色体上的多个高亲和力位点(也称为染色质进入位点CES,chromatin entry site)[8],携带自身的组蛋白修饰子从CES 顺式扩散到侧翼序列,对相关位点的组蛋白进行修饰以改变染色质结构,例如,将MOF 蛋白富集至X 染色体,促进X 染色体连锁基因的转录起始,JIL-1 会导致组蛋白H3 第10 位丝氨酸残基磷酸化,进而实现雄性X 连锁基因表达的2 倍上调[9]。
2.1 MSL 复合体介导的剂量补偿机制 MSL 复合体结合会导致H4K16Ac 的增加,从而使染色质更加松散。然而,这种调节在剂量补偿中的作用尚不清楚。随着技术的不断进步,研究者对MSL 复合体介导的剂量补偿机制存在几种不同的猜想。
“转录伸长模型”认为,H4K16Ac 修饰在剂量补偿中的潜在作用是减少剂量补偿基因转录过程中的RNA 聚合酶Ⅱ的抗扭强度[10]。但由于缺乏雌、雄之间的比较,很多研究者对此猜想提出了批评,这导致了启动子近端RNA 聚合酶Ⅱ富集模型的出现。最初,在雄性X 连锁基因启动子处的RNA 聚合酶富集约2.2 倍,从而验证了该模型[11]。而之后的研究发现,分析流程中的计算错误导致X 连锁基因之间的RNA 聚合酶富集数据夸大,这就消除了仅基于启动模型的可能性[12]。在进一步使用核苷酸水平的RNA 聚合酶Ⅱ作图后,研究者提出了“跳跃开始和获得”模型。H4K16Ac 的发生提高了释放RNA 聚合酶Ⅱ的速率并降低了早期转录终止的风险,使得RNA 聚合酶Ⅱ在基因中过度积累,从而导致转录产物的增加[13]。
2.2 激活模型存在的问题 由非整倍体所引发的基因表达变化不仅局限于发生剂量改变的染色体连锁基因,而是在全基因组具有广泛性。激活模型虽能解释两性间X 染色体中的剂量补偿效应,但不能解释X 染色体与常染色体之间的基因表达失衡现象。研究者检测了含有3 条染色体的超雌果蝇的基因表达情况,发现超雌体果蝇中不含有MSL 复合体,但在X 连锁基因中同样发现了剂量补偿现象,且常染色体的基因表达水平出现了不同程度的下调[14]。此外,激活模型无法解释雄性染色体相关基因的性别二态性的倾向,对于雄性果蝇X 连锁基因如何精确实现2 倍上调的机制,该模型也没有给出准确的描述。
3 总结与展望
随着基因剂量补偿研究的不断深入,激活模型已无法解释一些相关现象,例如,X 染色体三体超雌果蝇中的基因剂量补偿及雄性常染色体表达中的性别二态性[1]。此外,多个实验室已报道了几种不同的非整倍体常染色体剂量补偿现象,在一定程度上表明,基因剂量补偿现象随染色体剂量的改变而固有发生,这与平衡模型中的反式剂量效应相符合。因此,平衡模型更能科学合理地解释基因剂量补偿的分子调控机制。
但平衡模型中仍存在尚未解决的问题,例如,当MSL 复合物被解离时,X 染色体相关基因的剂量补偿现象是否会消失。相关研究表明,基因的剂量补偿现象并未消失,但是常染色体上的基因表达水平呈现上调[1]。因此,常染色体的基因表达水平就无法作为标准值检测X 染色体基因表达量,并且常染色体的变化有可能会改变调节机制的化学计量学[3],有关基因剂量补偿丧失与否需要重新考量。
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
大自然探索(2023年11期)2023-03-01 09:04:36
学苑创造·A版(2022年3期)2022-03-29 23:32:16
中国生殖健康(2020年5期)2021-01-18 03:00:06
学苑创造·A版(2019年6期)2019-07-11 01:07:39
中国生殖健康(2018年5期)2018-11-06 07:15:56
食品与机械(2018年5期)2018-07-16 01:34:00
中国民族医药杂志(2016年5期)2016-05-09 07:43:50
原子与分子物理学报(2015年1期)2015-11-24 12:49:26
食品科学(2013年15期)2013-03-11 18:25:51
