
Calycosin attenuates severe acute pancreatitis-associated acute lung injury by curtailing high mobility group box 1 - induced inflammation
2021-12-06 08:52:40ChangJuZhuWanGuangYangDeJianLiYaoDongSongSanYangChenQiaoFangWangYanNaLiuYanZhangBoChengZhongWeiWuZongChaoCui
World Journal of Gastroenterology 2021年44期
Chang-Ju Zhu, Wan-Guang Yang, De-Jian Li, Yao-Dong Song, San-Yang Chen, Qiao-Fang Wang, Yan-Na Liu,Yan Zhang, Bo Cheng, Zhong-Wei Wu, Zong-Chao Cui
Abstract
Key Words: Severe acute pancreatitis; Acute lung injury; Calycosin; Mouse model; Highmobility group box 1 ; Nuclear factor-kappa B
INTRODUCTION
Acute pancreatitis (AP) is an inflammatory disease with wide clinical variation,resulting in an approximately 35 % mortality when progressing to severe AP (SAP)[1 ].Acute lung injury (ALI) is the most common cause of death in patients with severe AP(SAP), occurring in 10 %-25 % of SAP cases and responsible for up to 60 % of APassociated deaths[2 ]. Inflammation and pro-inflammatory cytokines play a key role in the development of SAP; therefore, inhibition of inflammation and the release of inflammatory factors are thought to be potential approaches for the therapy of SAPALI.
High mobility group box 1 (HMGB1 ), a highly conserved DNA binding nuclear protein, plays a vital role in the pathogenesis of inflammatory diseases such as pancreatitis[3 ]. Secreted HMGB1 released from necrotic acinar cells has been shown to aggravate the pancreatic inflammatory process[3 ,4 ]. Secreted HMGB1 exhibits cytokine-like properties that induces both the local and systemic inflammatory cascade that ultimately leads to multi-organ dysfunction[5 ,6 ]. HGMB1 has been shown to activate pro-inflammatory nuclear factor-κ-B (NF-κB) signalingviainteraction with multiple cell-surface receptors including Toll-like receptor (TLR) 2 , TLR4 or TLR9 and receptor for advanced glycation end products (RAGE)[7 ]. The activation of NF-κB upregulates the gene expression of pro-inflammatory cytokines, chemokines and adhesion molecules which further aggravates the inflammatory response[4 ]. HMGB1 has also been shown to serve as a chemo-attractant recruiting neutrophils to site of inflammation and prevents neutrophil apoptosis which exacerbates tissue damage[8 -10 ]. Blockade of HMGB1 by administration of anti-HMGB1 neutralizing antibodies was shown to inhibit the recruitment and accumulation of neutrophils in the lung[11 ,12 ]. Thus, HMGB1 is a potential target for the treatment of ALI that is commonly found in SAP.
Calycosin (Cal) is one of the bioactive constituents extracted from the Chinese medicinal herbRadix Astragali, one of the five herbs of the Wutou Decoction, a classic herbal formula concocted by ancient Chinese medical doctor, Zhongjing Zhang,widely used for the treatment of rheumatoid arthritis[13 ]. Cal is a phytoestrogen isoflavone that has been shown to exhibit various biological effects including potent anti-inflammatory properties[14 ], as well as anti-cancer[15 ], neuroprotective[16 ], anti-Parkinson activity[17 ].
However, no studies have assessed the potential use of Cal for the expression of HMGB1 in the treatment of ALI in SAP. Hence, this study aims to address this question by exploring the effects of Cal administration on the expression of HMGB1 both in LPS induced ALIin vitroand an L-arginine induced ALI model in mice with SAP.
MATERIALS AND METHODS
Chemicals and reagents
L-arginine (L-arg: purity > 98 %, endotoxin-free) and the BCA Protein Assay Kit were purchased from Beijing Solarbio Science and Technology Co., Ltd. (Beijing, China).Calycosin (Cal: C16 H12 O5 , purity > 98 %;) was from Chengdu Biopurify Phytochemicals Ltd. (Chengdu, China). Enzyme-linked immunosorbent assay (ELISA) kits for interleukin (IL)-6 , HMGB1 , IL-1 β and MPO were obtained from Wuhan Cloud-Clone Corp. (Wuhan, China). ELISA kits for tumor necrosis factor (TNF)-α and CXCX-1 were procured from Proteintech Group (Rosemont, IL, United States). The amylase ELISA kit was bought from Shanghai BlueGene Biotech Co., Ltd. (Shanghai, China). Primary antibodies against NF-κB p65 (p65 ), phosphorylated NF-κB p65 (p-p65 ), and GAPDH were purchased from Cell Signaling Technology Inc. (Danvers, MA, United States).Primary antibody against lymphocyte antigen 6 complex locus G6 D (Ly6 G) was obtained from Abcam (Cambridge, United Kingdom). Primary antibody against HMGB1 and Fluorescent secondary antibody were produced by Proteintech Group.Horseradish peroxidase (HRP)-conjugated secondary antibodies and Hypersensitive WB Chemiluminescent Substrate Reagent were from Beyotime Biotechnology (Jiangsu,China).
Animals
Twenty-four male C57 BL/6 N mice (weight: 18 -22 g, age: 8 –10 wk) were purchased from Charles River Company (Beijing, China). The mice were housed in a specific pathogen-free facility with a dark/light cycle of 12 /12 h in ambient temperature of 22 ± 2 ℃ and humidity of 50 % ± 10 %. Mice were fed standard rodent chow and clean waterad libitum. All animal experiments were conducted in accordance with relevant guidelines and regulations and approved by the Animal Ethics Committee of The National Drug Clinical Trial Institution of The First Affiliated Hospital of Zhengzhou University (Ethic Review Number: 2019 -KY-140 ). All mice received humane care and the study was conducted following the ARRIVE guidelines.
Murine model of L-arginine-induced severe acute pancreatitis
L-arg was dissolved in normal saline and then sterilized by filtration (pH approximately equal to 7 .0 ). Mice were randomly divided into four groups (n = 6 in each group): Control (Saline), L-arg (4 g/kg + Saline), L-arg + Low-dose Cal (L, 25 mg/kg bodyweight), and L-arg + High-dose Cal (H, 50 mg/kg bodyweight). The Cal treatment groups received prophylactic Cal treatment (25 or 50 mg/kg)viaintraperitoneal injection 1 h before the first injection of L-arg. The Control and L-arg groups received an intraperitoneal injection of normal saline before L-arg injection. After the 1 h prophylactic treatment, the mice received intraperitoneal injections of either normal saline (Control group) or 4 g/kg of L-arg every hour for 2 h to induce severe AP as previously described by Dawraet al[18 ]. Blood samples were drawn from the retroorbital venous plexus under general anesthesia using sodium pentobarbital. Blood samples were centrifuged at 3000 rpm for 15 min at 4 °C and plasma serum stored at -80 °C for downstream biochemical analyses of serum amylase and cytokine levels.Mice were then sacrificed, and the pancreas and lung tissues from each mouse were quickly removed. The pancreas and left lung were dissected in two with one half being fixed in 4 % paraformaldehyde (PFA) for histopathological assessment and the other half snap-frozen in liquid nitrogen and stored at -80 ℃ for biochemical analysis.
Measurement of wet-to-dry weight ratio in lung tissue
Lung tissue wet-to-dry weight (W/D) ratio was employed to determine the extent of pulmonary edema following L-arg administration. The right lung was excised and surface water was removed by blotting with filter paper. The lung weight was immediately measured on a standard electronic laboratory scale and recorded as the wet weight (W). The lung was then dried in an oven at 60 °C for 48 -72 h and reweighed as dry weight. The W/D ratio was calculated based on the following formula: W/D =(wet weight − dry weight)/dry weight.
Histopathological assessments
Fixed pancreatic and lung tissues were embedded in paraffin blocks and 4 μm thin sequential sections were prepared. Tissue sections were stained with hematoxylin and eosin as per our standard laboratory protocol. Stained sections were visualized and imaged under an optical light microscope (CX31 , Olympus Optical Co., Ltd., Japan),and histopathological changes were assessed by three experienced pathologists who were blind to the experimental procedure. Pancreatic tissue damage was graded using a modified Schmidt Scoring System[19 ] as normal to severe (scale of 0 -4 ) based on the degree of inflammatory cell infiltration, vacuolization and acinar cell necrosis.Similarly, lung tissue sections were assessed for alveolar thickening and inflammatory cell infiltration with scoring system ranging from 0 -3 [20 ].
Immunohistochemical evaluation of Ly6 G expression
For immunohistochemical staining, lung tissue sections were deparaffinized and rehydrated in graded ethanol. Sections were immersed in 3 % hydrogen peroxide (in methanol) for 20 min to block endogenous peroxidase activity, and then boiled in 0 .1 %citrate buffer for antigen retrieval, followed by incubation in 3 % BSA serum (in PBS)for 30 min at room temperature to block non-specific immuno-reactivity. Tissue sections were incubated overnight at 4 °C with anti-Ly6 G antibody (1 :500 dilution in 3 % BSA-PBS) to stain neutrophils. Following incubation with HRP-conjugated secondary antibody color development was achieved by incubating sections with diaminobenzidine color development reagent and visualized under an optical light microscope. Five non-overlapping high-power fields (×100 magnification) for each section were captured. The integrated optical density (IOD) of positive expression for Ly6 G in lung tissue sections were measured from gray-scale images using Image J software (Image J 1 .52 , National Institute of Health) following calibration of hue (0 –25 ), saturation (0 -255 ), and intensity (0 -255 ) levels in the area of interest. The AOD(relative expression) was determined as IOD/positive area.
Serum enzymes and cytokine quantification using ELISA
Blood samples collected were subjected to ELISA analysis to determine serum levels of amylase, TNF-α, IL-6 , IL-1 β, CXCL-1 and HMGB1 in accordance with the corresponding manufacturer’s protocol. Serum amylase level was expressed as U/L and serum TNF-α, IL-6 , IL-1 β, CXCL-1 and HMGB1 Levels were expressed as pg/mL.
MPO activity in lung tissues
To assess lung MPO activity, 20 mg of frozen lung tissue were homogenized in homogenization buffer (0 .5 % hexadecyl trimethylammonium bromide, 5 mmol/L EDTA, and 50 mmol/L potassium phosphate buffer; pH 6 .2 ) on ice. The homogenate was then centrifuged at 12000 × g for 15 min at 4 °C and the resulting supernatant was retained. MPO activity was determined in accordance with the corresponding manufacturer’s protocol, MPO levels were expressed as pg/mg.
In vitro model of LPS-induced ALI in A5 49 cells
The lung adenocarcinoma A549 cell line was purchased from ATCC (VA, United States) and maintained in RPMI-1640 medium containing 10 % fetal bovine serum and 100 IU/mL streptomycin and 100 IU/mL streptomycin. Anin vitrocellular model of ALI was established by treating A549 cells with 1 µg/mL lipopolysaccharide (LPS,Sigma, United States) for 24 h[21 ]. To test the effects of Cal, cells were pretreated with various concentrations of Cal (1 µM, 5 µM, 10 µM, and 20 µM) for 1 h before LPS stimulation. After 24 h of stimulation, cells were either fixed for immunofluorescence assessment, or harvested for protein extraction for downstream western blot analyses.
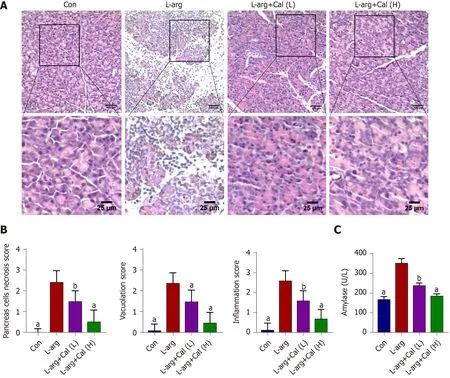
Figure 1 Effects of calycosin on L-arginine induced severe acute pancreatitis. Mice were pretreated with calycosin [calycosin (Cal): 25 and 50 mg/kg]for 1 h, L-arginine (L-arg: 4 g/kg) was intraperitoneally (i.p.) injected every hour for two consecutive hours. A: The histological assessment of pancreatic tissue damage in the control (Con), L-arg and Cal group (L: 25 mg/kg; H: 50 mg/kg); B: Pancreatic histological scores; C: Serum amylase and lipase levels. Data represent mean ± SD values. aP < 0 .001 vs L-arg group; bP < 0 .01 vs L-arg group. Con: Control group; Cal: Calycosin group; L-arg: L-arginine group.
CCK-8
The CCK-8 assay was used to assess the effects of Cal on A549 cell viability. Cells were seeded onto 96 -well plates at a density of 1 ×104 cells/well and then treated with various concentrations of Cal (1 , 5 , 10 and 20 µM) for 24 h. DMSO (0 .1 %) was added to the 0 µM group. After treatment, the cells were incubated with 10 μl of CCK-8 reagent for 1 h (Bosterbio, United States) and then the absorbance at 450 nm was measured using a Microplate Reader.
RT-qPCR
Total RNA was extracted using TRIzol reagent. Two microgram of total RNA was reversed transcribed into cDNA using Hiscript II Q RT SuperMix for qPCR in accordance with the manufacturer’s instructions (Nanjing, China). Real-time quantitative PCR was carried out using the SYBR Green qPCR Master Mix, containing template cDNA and specific primers for TNF-α, IL-6 , IL-1 β, CXCL-1 , or HMGB1 . PCR reactions were carried out on an ABI Prism Real-time PCR System (Applied Biosystems, Foster City, CA, United States). GAPDH was used as an internal housekeeping control. The primers used are listed in Supplementary Table 1 .
Immunofluorescence staining
The expression of HMGB1 and NF-κB (p-p65 ) in A549 cells and lung tissue was evaluated by immunofluorescence. Briefly, cells was washed twice with PBS, fixed with 4 % paraformaldehyde (PFA) for 30 min, and then permeabilized with 0 .5 % Triton X-100 in TBS-Tween 20 (TBST) for 5 min. For lung tissues, sections were dewaxed,hydrated, and treated with EDTA-containing antigen retrieval buffer (pH 8 .0 ) in a microwave oven, and then blocked with 5 % BSA for 1 h. Samples (cells or tissue sections) were then incubated with the primary antibodies (HMGB1 , 1 :100 ; or NF-κB p-p65 , 1 :200 ) in TBST overnight at 4 °C. Samples were washed three times with TBST and then incubated with Cy3 –conjugated Affinipure Goat Anti-Rabbit IgG (H + L)antibody (1 :100 , Proteintech) for 1 h. After washing three times with TBST, the samples were incubated with DAPI to stain the nuclei and then mounted in anti-fade mounting medium for assessment by fluorescence microscopy.

Figure 2 The effects of calycosin on histopathological findings of lung tissue in mice. H&E sections were examined by light microscopy. A:Representative pathological images of the lung tissue; B: Lung injury scores in alveolar thickness; C: Inflammatory infiltrate; D: The lung wet/dry weight ratios are shown. Data represent mean ± SD values. aP < 0 .001 vs L-arg group; bP < 0 .01 vs L-arg group. Con: Control group; Cal: Calycosin group; L-arg: L-arginine group.
Western blot analysis
Frozen cells or lung tissues were lysed and homogenized in RIPA lysis buffer containing protease inhibitors and 1 mmol/L phenylmethanesulfonylfluoride (PMSF)on ice. Lysates were then mixed with SDS loading buffer and denatured by heating at 100 °C for 10 min. Each protein sample was separated by SDS-PAGE electrophoresis(10 % gel), and then transferred onto polyvinylidene difluoride membranes.Membranes were blocked with 5 % (w/v) skim milk in TBST for 1 h at room temperature and then incubated with primary antibodies overnight at 4 °C. The following primary antibodies and dilutions were used: anti-p65 (1 :1000 ), anti-p-p65 (1 :1000 ), anti-HMGB1 (1 :500 ) and anti-GAPDH (1 :1000 ). Following extensive washing with TBST the membranes were incubated with the HRP-conjugated secondary antibody (1 :1000 in 1 % (w/v) skim milk in TBST) for 1 h at room temperature. Proteinantibody immunoreactivity was detected by Hypersensitive Chemiluminescent Reagent and imaged on a LI-COR Odyssey Imaging System (LI-COR, Lincoln, NE,United States). Densitometry analysis was performed using the associated software and the band intensity of target proteins was normalized to GAPDH signals.
Molecular docking

Figure 3 Effect of calycosin on tumor necrosis factor α, interleukin-6 , interleukin-1 β, chemokine ligand 1 and high mobility group box 1 in serum and mRNA levels. A: TNF-α; B: IL-6 ; C: IL-1 β; D: CXCL-1 ; E: HMGB1 in serum levels were determined by ELISA kits. F: TNF-α; G: IL-6 ; H: IL-1 β; I:CXCL-1 ; J: HMGB1 in mRNA levels. Data represent mean ± SD values. aP < 0 .001 vs L-arg group; bP < 0 .01 vs L-arg group; cP < 0 .05 vs L-arg group. TNF-α: Tumor necrosis factor α; IL-6 : Interleukin-6 ; IL-1 β: Interleukin-1 β; HMGB1 : High mobility group box 1 ; CXCL-1 : Chemokine ligand 1 ; L-arg: L-arginine; Con: Control group;Cal: Calycosin group; L-arg: L-arginine group.
Molecular docking analysis of the interaction(s) between Cal and HMGB1 was carried out using the open-source Autodock Vina v1 .1 .2 software (Scripps Research, CA,United States). Two-dimensional (2 D) coordinates of Cal were retrieved through the PubChem website (https://pubchem.ncbi.nlm.nih.gov). The three-dimensional structure of HMGB1 A-box (PDB ID: 2 RTU) was retrieved from the RCSB Protein Data Bank. Optimized binding conformations were generated using criteria such as energy minimization and cluster size. To increase the accuracy of the binding conformations generated, the value of exhaustiveness was set to 1 . Finally, the superposition of Cal and HMGB1 was carried out. Receptor proteins and ligand molecules were converted into PDBQT formats.
Statistical analysis
All bar graphs presented in this study are expressed as mean ± SD values of at least three independent experiments. Differences between 4 groups were compared by oneway analysis of variance using GraphPad Prism 8 .0 .2 (GraphPad Software Inc., San Diego, CA, United States). Statistical significance was set at aPvalue < 0 .05 unless otherwise stated.

Figure 4 The effect of calycosin on the immunological staining of lung slices for Ly6 G. A: Micrographs of lung section stained with immunological staining of lung slices for Ly6 G (brown); B: Average optical density for Ly6 G was determined; C: Levels of myeloperoxidase in per milligram lung tissue were determined by enzyme-linked immunosorbent assay kits. Data represent mean ± SD values. aP < 0 .001 vs L-arg group; bP < 0 .01 vs L-arg group; cP < 0 .05 vs L-arg group. Con: Control group; Cal: Calycosin group; L-arg: L-arginine group.
RESULTS
Cal treatment protects against ALI in mice with SAP
As shown in Figure 1 A and B, the pancreas from mice treated with L-arg exhibited classical histological signs of SAP including significant tissue damage, acinar cell vacuolization and necrosis, as well as abundant inflammatory cell infiltration when compared with control mice (P< 0 .001 ). On the other hand, mice that were administered prophylactic Cal throughout the experimental period showed a dosedependent reduction in pancreatic tissue damage (Figure 1 A). Elevated serum amylase is a key indicator of pancreatic acinar cell injury in AP. Amylase activity was markedly elevated (more than 2 -fold) at 72 h following the induction of SAP in the L-arg group when compared to the control group (Figure 1 C; P < 0 .001 ). Consistent with histological findings, Cal administration dose-dependently decreased serum amylase levels when compared to the L-arg SAP group (P< 0 .001 ). Taken together, these results indicated that Cal administration can protect mice against L-arg-induced SAP.
SAP is often accompanied by ALI and contributes to the majority of AP-associated death[22 ,23 ]. We examined whether secondary ALI was similarly induced in our SAP mouse model following L-arg treatment. As shown in Figure 2 A-C, lung tissue from control mice showed normal pulmonary architecture, while the lung tissue in the Larg SAP group exhibited significant lung edema, alveolar wall thickening, local hemorrhage, and inflammatory cell infiltration (P< 0 .001 ). In contrast, the aforementioned histopathological observations in the Cal treatment groups exhibited noticeable improvement particularly when mice were treated with 50 mg/kg Cal (high-dose).Alveolar capillary permeability or pulmonary edema was assessed using the W/D ratio. Consistent with secondary ALI, L-arg SAP mice exhibited a markedly elevated W/D ratio compared with controls (Figure 2 D; P < 0 .001 ), whereas Cal treatment significantly reduced pulmonary edema (lower W/D ratio) in a dose-dependent manner. Together, our results showed that Cal treatment can further alleviate secondary ALI in mice with SAP.

Figure 5 Effects of calycosin on the expression of high mobility group box 1 in lung tissues. A: Western blot detection of high mobility group box 1 (HMGB1 ) expression in lung tissue; B: Quantitative analysis of HGMB1 in lung tissue; C: Immunofluorescence staining of HMGB1 in lung tissue. Data represent mean ± SD values. aP < 0 .001 vs L-arg group; bP < 0 .01 vs L-arg group; cP < 0 .05 , vs L-arg group. L-arg: L-arginine; HMGB1 : High mobility group box 1 ; Con: Control group; Cal: Calycosin group.
Cal inhibits pro-inflammatory cytokine expression and secretion
The expression and release of pro-inflammatory cytokines and mediators are critical effectors that exacerbate local pancreatic tissue damage and mediate systematic inflammation during SAP[24 ]. Thus, serum levels of pro-inflammatory cytokines and chemokines such as TNF-α, IL-6 , IL-1 β, CXCL-1 and HMGB1 were determined using ELISA and their mRNA levels in lung tissue were also determined. Consistent with histological observations, serum levels of TNF-α, IL-6 , IL-1 β, CXCL-1 and HMGB1 were markedly elevated in the L-arg SAP group when compared with controls(Figure 3 A-E, respectively; P < 0 .001 ) as well as their mRNA expression levels(Figure 3 F-J). In contrast, Cal treatment dose-dependently reduced the secretion and serum concentration of these pro-inflammatory cytokines and chemokine when compared with the L-arg SAP group.
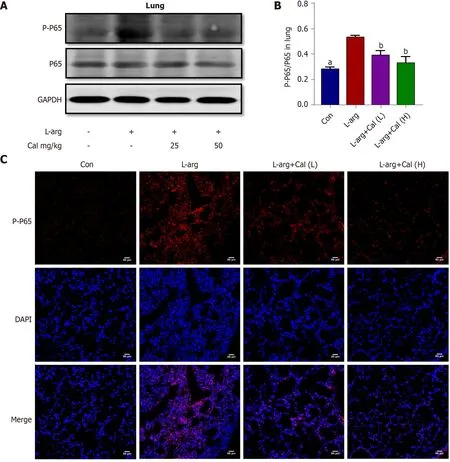
Figure 6 Effects of calycosin on the expression of phosphorylated nuclear factor-kappa B-p65 expression in lung tissues. A: Western blot detection of phosphorylated nuclear factor-kappa B-p65 (NF-κB-p65 ) expression in lung tissue; B: Quantitative analysis of phosphorylated NF-κB-p65 in lung tissue;C: Immunofluorescence staining of phosphorylated NF-κB-p65 in lung tissue. Data represent mean ± SD values. aP < 0 .001 vs L-arg group; bP < 0 .01 vs L-arg group.p-p65 : Phosphorylated nuclear factor-kappa B-p65 ; Con: Control group; Cal: Calycosin group; L-arg: L-arginine group.
Cal prevented neutrophil infiltration in the lung
Previous studies have shown that the accumulation of a large number of infiltrating neutrophils in the lungs is a pathophysiological feature of ALI[7 ,8 ]. We assessed whether this corresponded with reduced neutrophil infiltration in the lungs. To this end, lung tissue sections were stained for Ly6 G, a specific marker that separates neutrophils from other inflammatory cells such as leukocytes[25 ]. As shown in Figure 4 A and B (P < 0 .001 ), a significantly greater number of Ly6 G-positive cells was observed in L-arg SAP mice than in control mice. This elevation in Ly6 G positivity in the L-arg SAP group was also correlated with marked increases in lung MPO activity(Figure 4 C; P < 0 .001 ) further attesting to significant neutrophil infiltration in the lungs following L-arg-induced SAP. On the other hand, Cal treatment dose-dependently reduced neutrophil infiltration as demonstrated by reduced Ly6 G positivity in lung tissues (Figure 4 A and B; P < 0 .05 and P < 0 .01 ) and diminished lung MPO activity(Figure 4 C; P < 0 .001 ). These results further strengthened the protective effects of Cal against secondary ALI associated with AP.
Cal suppressed the activation of HMGB1 and the NF-κB signaling pathway in vivo

Figure 7 Effects of calycosin on the cell viability of A549 cells. A: Cell vitality was detected by the CCK8 assay after calycosin Cal (1 µM, 5 µM, 10 µM,and 20 µM) treatment; B: Cal pretreatment markedly increased cell vitality induced by lipopolysaccharide (LPS, 1 µg/mL). Data represent mean ± SD values. aP <0 .001 vs LPS group; cP < 0 .05 vs L-arg group. Con: Control group; Cal: Calycosin group; LPS: Lipopolysaccharide group.
Previous studies have shown that many of the pro-inflammatory effects of extracellular HMGB1 is driven by activation of the NF-κB signaling pathway[3 -4 ]. To examine whether NF-κB signaling is involved in mediating the inflammatory effects of HMGB1 in ALI in vivo, we examined HMGB1 and NF-κB expression and signaling activation using immunofluorescence and western blot analyses, respectively. Our results showed that the expression of HMGB1 (Figure 5 A-C) and p-p65 (activated form of NF-κB p65 subunit) (Figure 6 A-C) was significantly elevated (P < 0 .001 ) in lung tissues following L-arg induced SAP. Treatment with Cal resulted in a dose-dependent decrease in HMGB1 and p-p65 expression in the lungs. These results suggest that the inhibition of HMGB1 , NF-κB signaling activation and pro-inflammatory cytokine secretion is in part associated with the protective effect of Cal against ALI in mice with SAP.
Cal treatment improved LPS-induced A5 49 cells viability and inflammatory response
To further define the molecular mechanism of the protective effects of Cal against ALI,anin vitromodel of ALI was established by stimulating the lung adenocarcinoma cellline A549 with LPS in the absence or presence of Cal. As shown in Figure 7 A,treatment with the indicated concentrations of Cal for 24 h did not exert detrimental effects on A549 cell viability (Figure 7 A). On the other hand, treatment with LPS (1 μg/mL) markedly reduced A549 cell viability as compared with untreated controls,while treatment with Cal resulted in a dose-dependent enhancement of cell viability especially at 20 μM (Figure 7 B). Based on these data, 20 μM of Cal was used for downstream investigations.
Cal suppressed the activation of HMGB1 and the NF-κB signaling pathway in vitro
The effects of Cal on HMGB1 and NF-κB p65 expression and activation were examined using the LPS-induced ALI cellular model. Immunofluorescence and western blot analyses demonstrated that LPS stimulation markedly induced HMGB1 and p-p65 expression (Figure 8 and Figure 9 ; P < 0 .001 ) and this was markedly attenuated following treatment with Cal.
The theoretical binding mode of the Cal and HMGB1 pathway
The interaction between HMGB1 A-box and Cal was assessed using molecular docking software. The HMGB1 structure consists of the A-box and B-box, two DNA binding domains and a negatively charged acidic C-terminal tail. The B-box fragment has a pro-inflammatory effect, while the A-box fragment is known to be antagonistic to inflammatory responses, when the A-box binds to the TLR4 receptor but is not able to trigger TLR4 dimer formation due to a lack of critical interactions with the TLR4 [26 ].As shown in Figure 10 , amino acid residues Arg73 , and Gly14 of HMGB1 interacts with Cal to form hydrogen bonds. Amino acid residues Glu77 , Lys11 , Lys10 , Asp8 ,Arg8 , Arg13 , and Pro12 of the HMGB1 A-box form hydrophobic interactions with Cal.These molecular docking results suggest that Cal exhibits distinct binding affinities to the HMGB1 A-box that inhibited its downstream signaling pathways.
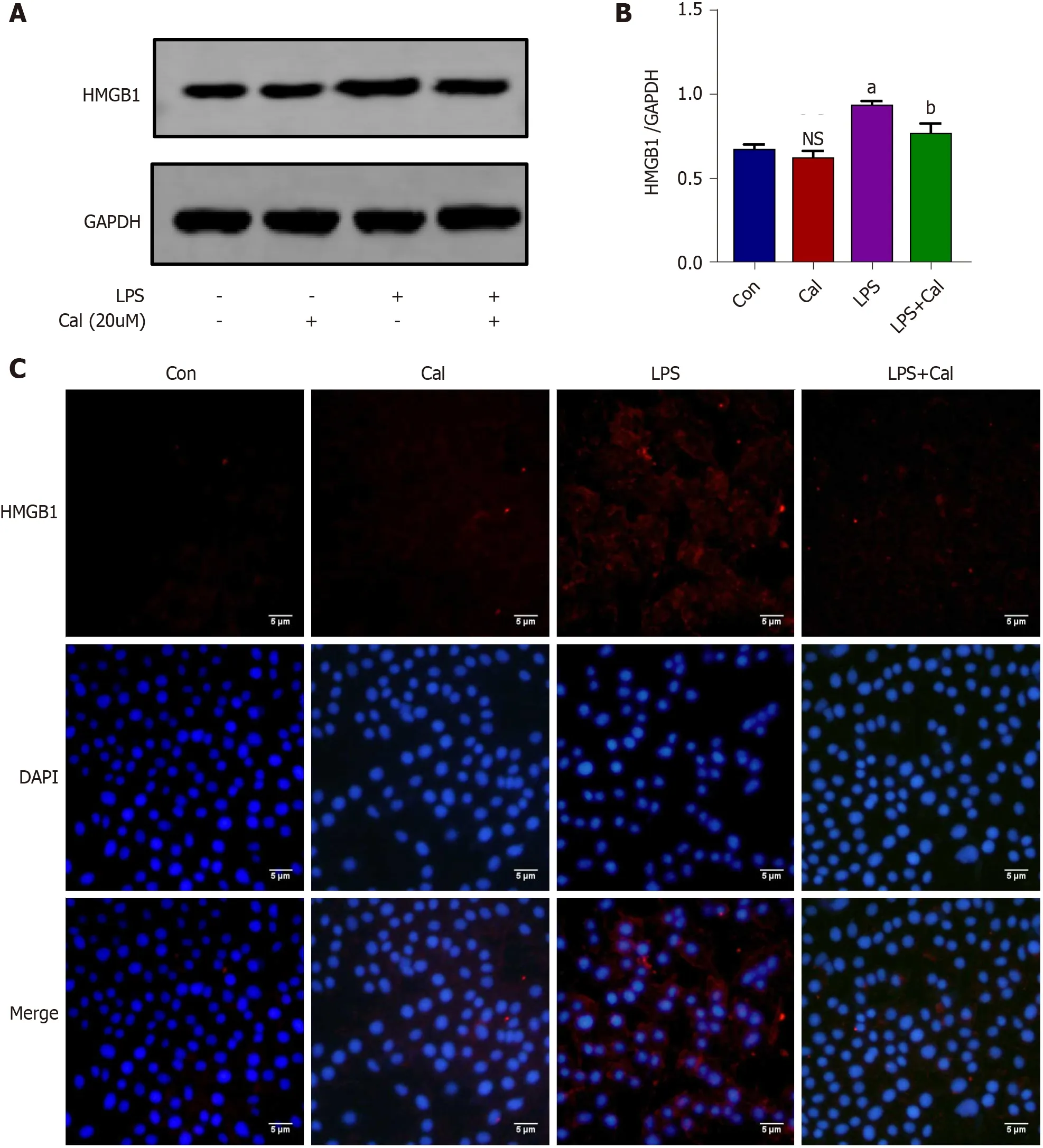
Figure 8 Effects of calycosin on the expression of high mobility group box 1 in vitro model of LPS-induced ALI using A549 cells. A:Western blot detection of high mobility group box 1 (HMGB1 ) expression in A549 cells; B: Quantitative analysis of HMGB1 in A549 cells; C: Immunofluorescence staining of HMGB1 in A549 cells. Control group (untreated), Calycosin group (Cal, 20 µM), LPS group and LPS + Cal group were treated with 1 µg/mL LPS for 24 h in the absence or presence of Cal (20 µM) pretreatment for 1 h. Data represent mean ± SD values. aP < 0 .001 vs control group; bP < 0 .01 vs LPS group. ns: No significance vs Con group. HMGB1 : High mobility group box 1 ; Con: Control group; Cal: Calycosin group; LPS: Lipopolysaccharide group.
DISCUSSION
SAP is one of the most common causes of acute abdominal pain and often manifests with many complications, resulting in high mortality[1 ]. The pathogenesis of AP is multifactorial and involves a multi-step process. In the early stages of the disease,intra-pancreatic activation of pancreatic enzymes such as trypsin, leads to autodigestion of acinar cells and initiates the production and release of various pro-inflammatory mediators (or cytokine cascade). The elevation in pancreatic pro-inflammatory cytokine levels induces pancreatic oxidative stress and increases vascular permeability, leading to pancreatic edema and acinar cell necrosis, which augments pancreatic inflammation[27 ,28 ]. At this stage, inflammatory cell infiltration and macrophage activation results in the further release of systemic cytokines and inflammatory mediators, leading to systemic inflammatory response syndrome. ALI is a common and severe complication associated with SAP[29 ]. Thus, identifying effective therapies that can effectively treat local and systemic tissue damage remains an unmet medical challenge.
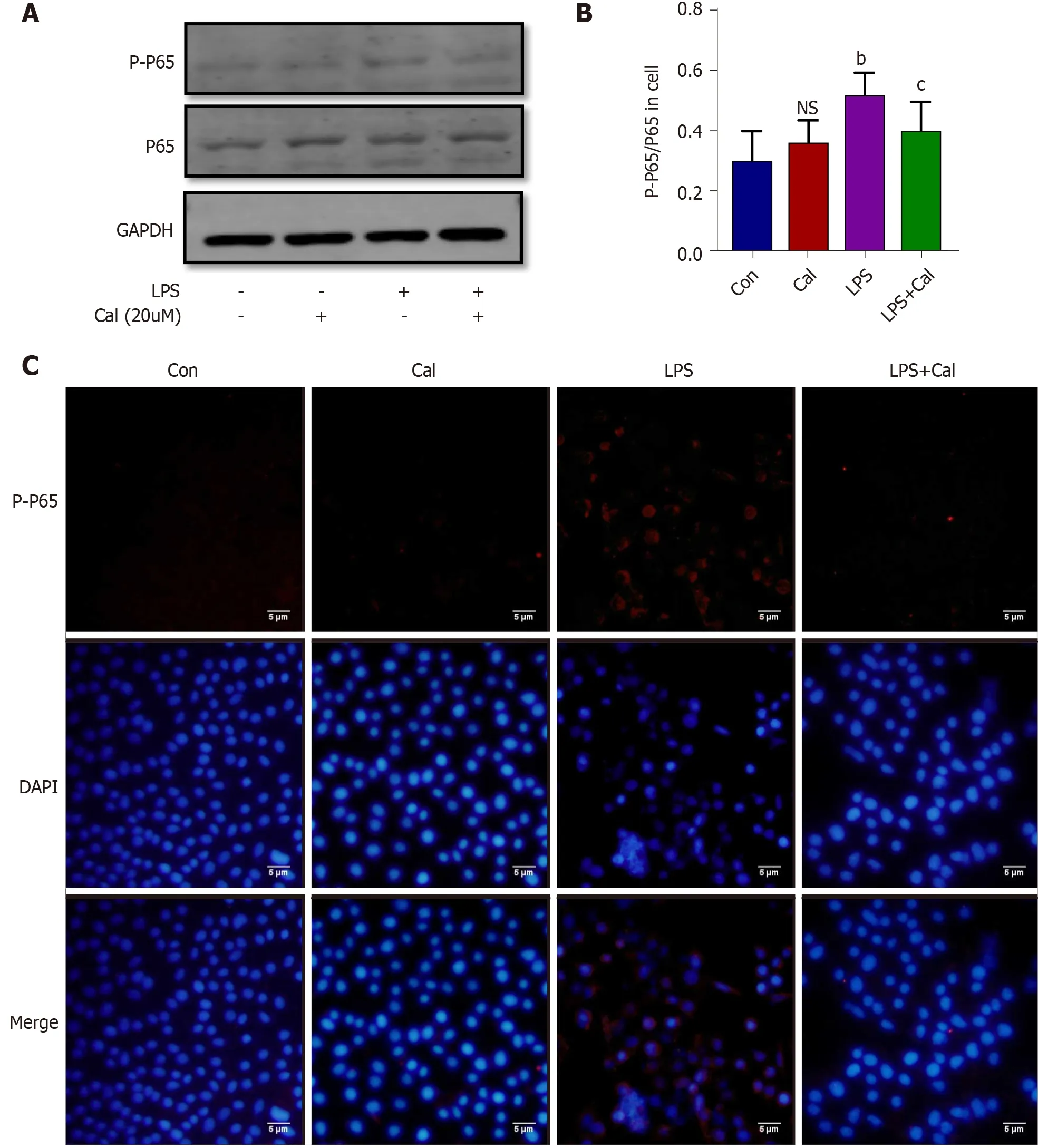
Figure 9 Effects of calycosin on the expression of phosphorylated nuclear factor-kappa B-p65 expression in the in vitro model of LPSinduced ALI using A549 cells. A: Western blot detection of phosphorylated nuclear factor-kappa B-p65 (NF-κB-p65 ) expression in A549 cells; B: Quantitative analysis of phosphorylated NF-κB-p65 in A549 cells; C: Immunofluorescence staining of phosphorylated NF-κB-p65 in A549 cells. Data represent mean ± SD values;bP < 0 .01 vs Con group; cP < 0 .05 vs LPS group. Ns: No significance vs Con group. p-p65 : Phosphorylated nuclear factor-kappa B-p65 ; Con: Control group; Cal:Calycosin group; LPS: Lipopolysaccharide group.
ALI is the most common extra-pancreatic complication leading to death in SAP patients, and there is no consensus on the most effective treatment[3 ]. Therefore, we aimed to explore the effect of Cal on ALI in SAP. We showed that Cal extracted from the Chinese medicinal herbRadix astragalieffectively protected mice against L-arginduced ALI in SAP. L-arg-induced SAP is a well-established model of pancreatitis that reflects the pathological changes seen in humans[18 ]. We found that mice prophylactically treated with Cal exhibited a marked decrease in serum amylase and tissue MPO activity, pronounced reductions in pancreatic and pulmonary lung tissue damage, and significantly diminished pro-inflammatory cytokine production in part due to inhibition of the HMGB1 /NF-κB signaling axis. The molecular docking analysis results suggest that Cal directly binds HMGB1viahydrogen bonds and hydrophobic interactionsviamultiple residues on HMGB1 . However, the implication of these residues on HMGB1 activity and function remains to be elucidated.
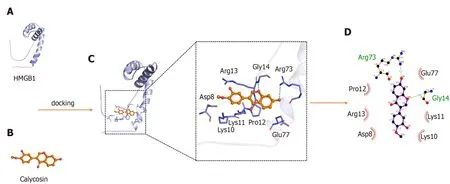
Figure 10 Theoretical binding mode of calycosin and high mobility group box 1 A-box. A: The 3 D structure of high mobility group box 1 (HMGB1 )A-box; B: The 3 D structure of calycosin (Cal); C: 3 D docking mode between Cal and HMGB1 A-box simulated by Discovery Studio and the amino acid of the active site; D: Two-dimensional schematic interaction diagram between Cal and HMGB1 A-box, the color of the amino acid residue is drawn by interaction. HMGB1 : High mobility group box 1 ; Cal: Calycosin.
We first assessed the effect of Cal on the histopathological injury in pancreas and lung tissue. The results revealed severe pancreatic histoarchitectural changes,including pancreatic edema and vacuolization, acinar cell necrosis, and inflammatory cell infiltration. The incidence of lung inflammation, including histopathological changes such as marked alveolar wall thickening, edema (demonstrated by increased lung wet/dry ratio), and pronounced inflammatory cell infiltration particularly by neutrophils (elevated lung MPO levels) was observed in SAP mice. In contrast, mice prophylactically treated with Cal showed dose-dependent amelioration of the severity of ALI in L-arg-induced SAP, as demonstrated by significant reductions in the histopathological manifestations and serum indices of SAP and ALI.
We then investigated the protective effect of Cal on the expression of inflammatory factors and neutrophil infiltration. In fact, the severity of local and systemic organ damage is dependent on the level of pro-inflammatory cytokine production[30 ]. Proinflammatory cytokines TNF-α, IL-1 β and IL-6 , play critical roles in the development and progression of SAP and perpetrators of systemic inflammatory response and organ damage, including ALI[31 ,32 ]. Blockade of serum TNF-α, IL-1 β and IL-6 has been shown to attenuate the disease process[33 ,34 ]. Consistent with previous findings and our histopathological observations, serum levels of TNF-α, IL-6 , and IL-1 β were elevated in SAP mice when compared with controls as well as the mRNA levels in lung tissue, while Cal treatment reversed the change. In particular, the infiltration of inflammatory cells, mainly neutrophils, is a hallmark of tissue injury in SAP. Previous studies have demonstrated that HMGB1 and chemokine (CXC motif) ligand 1 (CXCL-1 ) play a role in the recruitment of neutrophils to the lungs, leading to tissue damage[8 ,35 ]. Functional inhibition of HMGB1 or CXC chemokines was shown to ameliorate tissue damage[11 ,12 ,36 ]. Similarly, we showed that Cal treatment suppressed the serum levels and gene expression of HMGB1 and CXCL-1 levels in the lung, with an associated reduction in neutrophil infiltration and MPO expression in lung tissue. This result may have partially contributed to the inhibitory effects of Cal on inflammatory factors and neutrophil infiltration.
Mechanistically, the HMGB1 -dependent activation of NF-κB has been implicated in the development of ALI[37 ]. Studies have shown that the suppression of HMGB1 expression using siRNA can inhibit NF-κB activation, reduce inflammatory reactions,and protect mice against developing ALI in SAP[38 ,39 ]. Consistent with these reports,we demonstrated that Cal treatment dose-dependently inhibited the expression of HMGB1 and NF-κB signaling activation bothin vivoandin vitro. In addition, studies have illustrated that HMGB1 mediates pancreatic pain by targeting RAGE and the CXCL12 /CXCR4 signaling axis in mice with AP[40 ], therefore, pain may be relieved in animals after Cal treatment.
This study provides an experimental basis for the clinical application of Cal, which may be a candidate for the treatment of SAP-ALI patients in the future. However,there are limitations to the present study. For example, Cal inhibited the HMGB1 /NFκB signaling pathwayin vivoandin vitroand validated the interaction through molecular docking. Therefore, the specific interaction between Cal and HMGB1 requires further study.

Figure 11 Calycosin attenuates acute lung injury in L-arginine induced severe acute pancreatitis by curtailing high mobility group box 1 -induced inflammation.
CONCLUSION
In summary, our data clearly demonstrated that Cal exhibits protective and beneficial effects against ALI in SAP by averting local and systemic neutrophil infiltration and the inflammatory response in partviathe suppression of HMGB1 -NF-κB signaling activation. (Figure 11 ).
ARTICLE HIGHLIGHTS


Research conclusions
Cal protects mice against L-arg-induced SAP and associated ALI by attenuating local and systemic neutrophil infiltration and the inflammatory responseviainhibition of HGMB1 and the NF-κB signaling pathway.
Research perspectives
Cal may be used as a potential medicine in SAP-ALI therapy.
 World Journal of Gastroenterology2021年44期
World Journal of Gastroenterology2021年44期
- World Journal of Gastroenterology的其它文章
- Proton pump inhibitors and colorectal cancer: A systematic review
- Autosomal recessive 333 base pair interleukin 10 receptor alpha subunit deletion in very early-onset inflammatory bowel disease
- Treatment of Helicobacter pylori infection in the presence of penicillin allergy
- Challenges in the diagnosis of intestinal neuronal dysplasia type B: A look beyond the number of ganglion cells
- COVID-19 : Effect on gastroenterology and hepatology service provision and training: Lessons learnt and planning for the future
- Role of early transjugular intrahepatic portosystemic stent-shunt in acute variceal bleeding: An update of the evidence and future directions