
宰后成熟对生鲜肉品质影响的研究进展
2021-12-02 09:20刘泽超张一敏毛衍伟董鹏程张文华杨啸吟梁荣蓉
食品科学 2021年21期
刘泽超,罗 欣,张一敏,毛衍伟,董鹏程,张文华,杨啸吟,*,梁荣蓉,*
(1.山东农业大学食品科学与工程学院,山东 泰安 271018;2.国家肉牛牦牛产业技术体系中卫试验站,宁夏 中卫 755000)
肉的品质指标主要包括肉色、多汁性、风味、嫩度和保水性,这些指标是决定消费者是否购买肉品的关键因素。为消费者提供性质稳定的高品质肉品一直是肉品行业追求的目标,而高品质肉制品生产技术也相应地成为了当前肉品工业生产迫切需要的技术[1]。
生鲜肉的成熟过程能显著提高肉的嫩度、风味和整体接受度,是肉类品质提升的重要手段。目前,主要的成熟方法有干法成熟和湿法成熟两种,这两种成熟方法在改善肉的适口性方面都有着积极的影响[2]。成熟早期的宰后生化变化及成熟过程中的酶系统、小热休克蛋白(small heat shock proteins,sHSPs)作用以及蛋白质修饰等都对肉的品质发挥着重要影响。另外,干、湿法成熟对肉品质的作用效果还取决于成熟条件和时间、肉的质量和成分(如肌肉类型、肌内脂肪和胴体质量)等因素,所以,为最大程度地提高肉品品质,研究人员一直在优化成熟方案,开发新型成熟技术。因此,关于肉品成熟技术的研究近年来始终是肉品领域的热点。
因此,本文系统地综述宰后早期的生化变化及其对肉品质的影响,并对成熟技术的成熟机制和目前尚存在的技术难题进行阐述;还详细介绍干湿法成熟的优化方案和近期涌现出来的一大批新型成熟技术,以及它们对肉品品质的改善效果,以期推动肉品成熟新技术的开发,提升肉品品质。
1 宰后生化变化对肉品质的影响
在成熟早期,伴随着肌肉向肉的转化,宰后肌肉发生了复杂的生理生化变化。其中磷酸肌酸的分解、糖酵解和氧化磷酸化进程等能量代谢过程、蛋白翻译后修饰(蛋白质巯基亚硝基化修饰和蛋白质磷酸化修饰)、宰后酶系统(钙蛋白酶系统、过氧化物还原酶(peroxiredoxins,Prdx)6、半胱天冬蛋白酶介导的细胞凋亡过程)和sHSPs等会影响宰后肌肉的pH值、肌原纤维的性质、肌肉收缩、细胞凋亡以及结构蛋白的降解等,进而对肉的嫩度、保水性、肉色及其稳定性等品质产生重要的影响[3]。
1.1 宰后能量代谢对肉品质的影响
动物屠宰放血后,肌肉组织供氧中断,肌肉细胞随即进入缺氧状态。为了维持此期间的细胞稳态,肌肉会发生一系列能量代谢进程,如磷酸肌酸的分解、糖酵解和氧化磷酸化进程等,其中以糖酵解途径尤为重要,会直接影响肉的品质(pH值、肌原纤维蛋白的性质和肉的嫩度等)[4]。宰后肌肉无氧状态下产生ATP的能力较低,腺嘌呤核苷三磷酸(adenosine triphosphate,ATP)的下降会引起肌球蛋白和肌动蛋白发生不可逆结合进而导致肌肉收缩,影响肉的嫩度和保水性。同时,宰后能量代谢过程中由pH值降低所引起的肉的酸化,以及累积产生的CO2、HCO-3、NH+4和乳酸等代谢副产物会影响肉的风味和质地[5]。而一些参与糖酵解和三羧酸循环的酶(丙酮酸激酶、磷酸葡萄糖变位酶-1、果糖-二磷酸醛缩酶、3-磷酸甘油醛脱氢酶、3-磷酸甘油脱氢酶、苹果酸脱氢酶、黄素还原酶等)可以生成琥珀酸或烟酰胺腺嘌呤二核苷酸(nicotinamide adenine dinucleotide,NADH)等,提高肌红蛋白的氧化稳定性,进而改善肉色及其稳定性。另外,Sergeeva等[6]研究表明胞浆pH值的下降可以在宰后早期阶段诱导细胞凋亡。
宰后肌肉的无氧糖酵解和ATP水解会使pH值发生显著的变化,pH值从7.2逐渐下降到5.6左右。宰后代谢的速率和程度会显著影响肉的品质。如肌肉的快速代谢,促使宰后pH值在1 h内迅速降低至极限pH值(pH 5.6左右),导致了苍白松软渗水(pale, soft, and exudative,PSE)肉的产生;而宰前应激导致的糖原缺失则会引起宰后代谢的过早终止,从而限制了pH值下降,产生黑干(dry, firm, and dark,DFD)肉。因此,宰后肌肉的异常代谢会影响宰后pH值的下降,进而对肉的颜色、嫩度和保水性产生不利影响[7]。
此外,AMP-激活蛋白激酶复合物(AMP-activated protein kinase,AMPK)作为一种丝氨酸/苏氨酸激酶,在能量代谢调节中发挥关键作用,它由一个α催化亚基和两个β和γ调节亚基组成[8]。AMPK在维持细胞稳态的过程中会受到Ca2+浓度、激素和升高的AMP/ATP等生化信号的影响。同时,AMPK是多种应激条件下细胞凋亡的重要调控因子,AMPK的激活可以调节下游信号分子促进细胞的凋亡[9]。Gao Yongfang等[10]研究发现,在宰后肌肉成熟过程中,CaCl2激活了AMPK,不仅可以通过增加糖酵解中乳酸脱氢酶和丙酮酸激酶的活性来提高糖酵解代谢水平,还可通过改变肌内环境诱导细胞凋亡,促进肉嫩度的改善,起到加速成熟的效果。
1.2 宰后酶系统对肉品质的影响
肉的成熟是一个复杂的过程,多种不同内源性蛋白酶都参与其中。这一过程从动物屠宰后就开始,酶对牛肉蛋白质的水解作用能够维持两周[11]。内源性蛋白水解系统对肌肉中肌原纤维蛋白细胞骨架的降解可以显著改善肉的食用品质,提高肉的嫩度、多汁性和风味等品质特性[12]。在众多内源性蛋白酶中,目前普遍认为钙蛋白酶系统在肉品品质方面起着至关重要的作用,也在研究中报道最多。然而,最新研究发现由半胱天冬蛋白酶介导的细胞凋亡过程、过氧化物还原酶家族以及蛋白质的翻译后修饰都会影响成熟肉品的品质。这些研究将进一步补充人们对宰后酶系统作用机制的认识。
1.2.1 钙蛋白酶系统对肉品质的影响
钙激活蛋白酶是一大类细胞内半胱氨酸蛋白酶,在骨骼肌的生理pH值下最活跃。钙蛋白酶系统包括μ-钙蛋白酶和m-钙蛋白酶、钙蛋白酶3和钙蛋白酶抑制蛋白四大成员[13]。现有证据表明μ-钙蛋白酶在蛋白质水解及肉类嫩化方面起着关键作用。μ-钙蛋白酶在Ca2+的作用下会发生自溶,激活后靶向作用于肌间线蛋白、肌联蛋白、伴肌动蛋白、肌钙蛋白T等骨架蛋白。而肌间线蛋白等主要结构蛋白的水解正是成熟过程中肉的保水性和嫩度得到改善的主要原因[14]。线粒体作为钙缓冲系统的一部分可以维持细胞内的钙稳态,是钙蛋白酶系统功能调控的重要细胞器。Dang等[15]研究表明线粒体可以通过调节细胞内的钙水平来提升μ-钙蛋白酶的活性,从而促进宰后蛋白质水解和肉的嫩化。此外,钙蛋白酶和其他蛋白酶(20S蛋白酶体)可能也会导致肌纤维骨架蛋白的降解[16],而肌纤维骨架蛋白的降解会使肌原纤维膨大,吸收肌原纤维外腔的水分,最终提高了肉的保水能力[17]。Qian Shuyi等[18]研究发现在宰后前4 d,钙蛋白酶通过调控肌肉中水分的分布和迁移,使得肌肉保持较高的持水力,而成熟后期肌肉的保水性则由肌间线蛋白酶、肌联蛋白酶和整合素蛋白酶控制。
据报道,m-钙蛋白酶的稳定性较μ-钙蛋白酶更高,但肌肉中的Ca2+浓度往往低于其激活浓度,因此,该酶无法有效促进肌原纤维蛋白的降解。钙蛋白酶3仅存在于骨骼肌中,且不受钙蛋白酶抑制蛋白的抑制作用,在蛋白水解及嫩度方面并没有发挥重要作用[13]。而钙蛋白抑制蛋白含有4 个抑制域,每个抑制域都能抑制钙蛋白酶的活性,影响宰后肌肉的蛋白水解和肉的嫩化进程[13]。
1.2.2 半胱天冬蛋白酶介导的细胞凋亡过程对肉品质的影响
宰后细胞凋亡过程主要是由半胱天冬蛋白酶家族介导产生凋亡信号,进而启动凋亡程序。半胱天冬酶系统是一类半胱氨酸天冬氨酸特异性蛋白水解酶,在动物宰后保持较高的活性,有助于肉的嫩化[19]。根据在细胞凋亡中的作用可以将半胱天冬酶分为引发半胱天冬酶(半胱天冬酶8、9、10、12)和效应半胱天冬酶(半胱天冬酶3、6、7)两大类[19]。效应半胱天冬酶可以被上游的启动子激活,一旦激活就会切割特定的底物(肌原纤维蛋白和细胞骨架蛋白),导致细胞凋亡。据报道,半胱天冬蛋白酶介导的细胞凋亡对宰后骨骼肌的蛋白水解和肉的嫩度都有积极影响[20]。
如图1所示,半胱天冬蛋白酶介导的细胞凋亡会受线粒体的调控,在细胞凋亡上游途径和半胱天冬蛋白酶途径以及其他下游凋亡途径中,线粒体位于整个凋亡通路的中心,是各个通路互相交汇的重要节点[21]。它释放的细胞色素c(cytochrome c,Cyt c)是整个凋亡通路的关键性分子。而Cyt c所介导的凋亡途径与Ca2+的浓度水平和线粒体膜间隙中B细胞淋巴瘤2(B cell lymphoma 2,Bcl-2)家族蛋白的调节控制有关[21]。最新研究发现,Bcl-2家族蛋白中促凋亡蛋白可加速Cyt c的释放,并与细胞凋亡因子结合形成凋亡复合体,聚集半胱天冬蛋白酶酶原9形成凋亡体,进而激活半胱天冬蛋白酶3、7,引发半胱天冬蛋白酶级联反应,导致细胞凋亡。同时,细胞凋亡蛋白抑制剂(inhibitor of apoptosis,IAPs)是半胱天冬酶3、7的抑制剂,一些促凋亡蛋白(Omi和Smac等)会抑制IAPs的产生,进而共同促进细胞凋亡反应的发生。此外,血液中残留的氧气也会使乳酸、琥珀酸等代谢物氧化产生活性氧自由基(reactive oxygen species,ROS),进而刺激线粒体膜上的通透性过滤孔(mitochondrial permeability transition pore,MPTP)释放Cyt c,激活下游的半胱天冬酶,导致细胞凋亡[22]。

图1 线粒体氧化磷酸化及凋亡通路途径[21-22]Fig. 1 Mitochondrial oxidative phosphorylation and apoptosis pathways[21-22]
然而,在线粒体固有凋亡通路的某些凋亡模型中,半胱天冬蛋白酶的广谱抑制剂并不能完全阻止细胞凋亡。这说明除了半胱天冬蛋白酶凋亡途径外,尚存在其他的细胞凋亡通路。所以,研究人员又提出了另一种线粒体凋亡通路——凋亡诱导因子(apoptosis-inducing factors,AIF)通路[22]。线粒体蛋白AIF是第一个不依赖于半胱天冬蛋白酶的凋亡因子,其作为特殊的胞外信号分子,可以触发MPTP释放AIF,导致细胞凋亡。Chen Cheng等[23]研究发现,在宰后成熟过程中,线粒体中AIF水平的变化与肉的嫩度和细胞凋亡密切相关,ROS诱导的氧化应激会提高线粒体膜的通透性,促进AIF的释放,从而导致AIF介导的细胞凋亡,进一步提高肉的嫩度。
因此,线粒体和线粒体中相关的蛋白酶在细胞凋亡中起着至关重要的作用。但是,最新研究发现,线粒体凋亡进程介导的肉类嫩化效果还与肌肉的肌纤维类型有关。Zhang Jiaying等[24]通过研究发现,与牛背最长肌相比,宰后初期牛腰大肌的线粒体凋亡发生速率较快,但并没有促进后续成熟过程中肌纤维蛋白的降解,这表明线粒体凋亡介导的嫩化过程可能存在肌肉类型特异性,关于这方面的内在机理还需进一步研究。
1.2.3 过氧化物还原酶6对肉品质的影响
Prdx6是一类非硒依赖型过氧化物酶家族,可以还原过氧化氢和烷基氢过氧化物。该家族共有6 个成员,过氧化物氧化还原酶1~5均含有两个半胱氨酸(Cys)残基,而Prdx6仅含有一个Cys残基。与其它成员不同,Prdx6具有非钙依赖型磷脂酶A2的活性[25],在控制哺乳动物细胞氧化还原状态和维持细胞膜的磷脂代谢中起关键作用,参与细胞内或细胞间的信号传导,促进细胞再生和增殖,并防止细胞凋亡[26]。
最近研究发现,Prdx6可能利用其过氧化物还原酶活性和磷脂酶A2活性参与对成熟肉品嫩度的调控。Jia等[27]通过对牛肉嫩度相关的蛋白质组进行研究发现Prdx6的表达量与嫩度呈正相关,而嫩度高的肉通常经历了较低的氧化应激,所以他们提出Prdx6可能是标记牛肉嫩度的潜在生物标签。另外,Prdx6具有谷胱甘肽过氧化物酶和非钙依赖型磷脂酶A2这两种功能酶的活性,Gagaoua等[28]发现Prdx6不仅改善了肉的嫩度,还与pH值的下降以及肉色稳定性等品质特征有关。
然而,目前大部分报道都集中在对Prdx6活性和生理作用的研究上,关于Prdx6对肉品质影响的研究则相对较少。动物宰后细胞的稳态被破坏,剧烈的环境变化会破坏机体的氧化还原体系和细胞膜结构,导致细胞内蛋白质和脂质氧化生成ROS并造成细胞损伤,最终影响肉的嫩度。而Prdx6具有的氧化还原酶活性可以清除骨骼肌细胞产生的ROS,进而调控一系列生物学进程来维持细胞内氧化还原稳态,提高肉的嫩度和肉色稳定性。Wu Wei等[29]研究发现Prdx6可作为肉品变色的生物标签,Prdx6的表达量与脂质氧化成显著的正相关,这可能是由于在贮藏后期ROS可以诱导肉品产生抗氧化蛋白,进而改善肉色。此外,Prdx6还可通过调控对过氧化脂质的还原来影响肉的嫩度。综上,Prdx6不仅可以清除宰后骨骼肌产生的过多ROS,还可以抑制ROS造成的脂质氧化,进而减缓蛋白质氧化和细胞程序性死亡进程。但目前还需更多研究来证实Prdx6与肉品质之间的关系。据报道,Prdx6或可成为预测、调控牛肉嫩度的一个新的关键因子[28]。
1.3 蛋白质翻译后修饰对肉品质的影响
蛋白质翻译后修饰在研究蛋白质多样性和复杂生物学调控方面发挥着重要作用,目前蛋白质翻译后修饰途径主要有磷酸化、乙酰化、亚硝酸化、羟基化、甲基化、泛素化等20多种常见的蛋白修饰[30]。蛋白质翻译后修饰在蛋白结构、功能、细胞信号和酶活性等方面都具有调节作用,赋予了改性蛋白新的功能和活性[31]。据报道,蛋白质翻译后修饰可以调节能量代谢、Ca2+释放、肌肉收缩和细胞凋亡等过程,从而影响肉的嫩度、保水性、肉色稳定性等品质指标变化,目前在研究蛋白质修饰影响肉品品质方面,以蛋白质亚硝基化和磷酸化修饰的研究报道最多,而其他宰后修饰对品质影响的研究也正成为该领域最新的研究热点之一。
1.3.1 蛋白质巯基亚硝基化修饰
一氧化氮(NO)作为一种重要的信号分子,具有多种控制肌肉代谢活动的功能,由骨骼肌细胞中高度表达的一氧化氮合成酶(nitric oxide synthase,NOS)持续催化产生[32]。蛋白质巯基亚硝基化是指由NO基团和蛋白质半胱氨酸的巯基共价结合形成S-亚硝基半胱氨酸残基(S-nitrosocysteine,SNO)的过程,是一种典型的氧化还原依赖的蛋白翻译后修饰,影响蛋白质在细胞内的构象、活性和功能,是发挥生物学作用的重要途径。蛋白质S-亚硝基化的程度取决于NOS的定位表达,且不同肌肉类型和不同动物宰后肌肉的NOS浓度和活性不同[31]。蛋白质S-亚硝基化可以参与调节蛋白质的活性、定位、稳定性及其相互作用,从而可能直接决定着翻译后的磷酸化、乙酰化、泛素化和基于胞嘧啶氧化还原等关键修饰。因此,人们推测蛋白质S-亚硝基化可能通过对成熟过程中的主要代谢过程(能量代谢、Ca2+释放、肌肉收缩和细胞凋亡等)的调节,在肉类品质变化方面发挥其潜在作用。
诸多研究已发现宰后蛋白质亚硝基化会通过多条途径影响肉的嫩度。Hou Qin等[33]研究发现,经NO供体处理的牛半膜肌的钙蛋白酶会加速自我降解,导致肌纤维蛋白和肌间线蛋白的降解量减少,而经NOS抑制剂处理的样品则加速这些蛋白的降解;因此,他们推测蛋白S-亚硝基化可能通过调节钙蛋白酶自溶和肌纤维蛋白的降解来影响肉的嫩度。相反,Liu Rui等[34]研究发现,成熟样品中许多蛋白质都具有较高的S-亚硝基化水平,这表明宰后肌肉中出现了S-亚硝基化而且该现象不断积累。同时,他们还发现S-亚硝基化可能作为肌肉细胞凋亡的调节因子,有利于成熟过程中肉嫩度的改善。另外,亚硝基化反应可以改变钙蛋白酶的活性基团,促进肌原纤维蛋白的降解,进而改善肉的嫩度。Wen Pengcheng等[35]研究发现μ-钙蛋白酶亚硝基化在一定程度上改善了肉的嫩度,这是由于μ-钙蛋白酶经亚硝基化后活性增加并抑制了其自身的降解,进而促进了肌纤维蛋白的降解。但是,此研究是以分离纯化的肌原纤维蛋白为底物,与肉内部复杂的真实环境存在区别,所以,μ-钙蛋白酶亚硝基化对肉嫩度的影响还需要做进一步的研究。也有学者认为,NO和蛋白质S-亚硝基化可以调节Ca2+信号调节,并对细胞凋亡过程中的Ca2+功能异常起重要作用。Cottrell等[36]发现,NO供体二乙胺可以提高羊肌浆网Ca2+摄取的最大速率,且降低了Ca2+摄取的敏感性,导致细胞质中的Ca2+浓度增加。肌浆中过多的Ca2+必然会加强肌动蛋白和肌球蛋白的结合,导致持续的肌肉收缩,不利于肉品嫩度的改善。所以,蛋白质S-亚硝基化对肉类嫩度的影响还需要进一步的研究。
此外,也有研究发现,糖原磷酸化酶、磷酸果糖激酶和丙酮酸激酶的过度亚硝基化修饰会抑制这些酶的活性,表明蛋白质的S-亚硝基化可以通过调节酶的活性来部分调控宰后糖酵解途径,减少PSE肉的产生[32]。
1.3.2 蛋白质磷酸化修饰
蛋白质磷酸化作为最重要的翻译后修饰之一,是指在蛋白激酶催化下将ATP或鸟嘌呤核苷三磷酸(guanosine triphosphate,GTP)上的磷酸基团转移到蛋白质氨基酸残基上的过程[37]。蛋白磷酸化对蛋白质功能及稳定性方面具有重要作用,据报道,宰后肌肉中的磷酸化蛋白与嫩度、肉色稳定性、pH值下降速率等有关。
首先,蛋白质磷酸化作为糖酵解代谢的关键调控因子,通过调节糖代谢酶的活性,影响宰后早期肌肉pH值的下降速率,进而影响肉的僵直进程和肉的品质变化。Chen Li等[37]研究发现,糖酵解的速率与磷酸果糖激酶、烯醇化酶和腺苷酸激酶同工酶的磷酸化水平呈负相关。Müller等[38]也发现糖原磷酸化酶在磷酸化酶激酶的作用下发生磷酸化修饰,转化为有活性的形式,促进了糖酵解,从而有利于肉的嫩化。同时,Chen Lijuan等[39]研究发现肌原纤维蛋白磷酸化可能通过糖酵解作用影响肉的僵直进程,最终影响肉的嫩度。此外,Li Meng等[40]研究发现,肌浆蛋白质磷酸化通过调节糖酵解的进程来提高pH值下降的速率和乳酸的累积,进而导致肉色稳定性降低,且较高的肌红蛋白磷酸化水平也不利于维持肉色的稳定。
肌原纤维蛋白磷酸化可能会通过抑制肌动球蛋白的解离来影响肌肉收缩,进而导致肉的嫩度降低。Cao Lichuang等[41]研究发现,肌球蛋白调控轻链(myosin regulatory light chain,MRLC)磷酸化与肌肉收缩有关,且MRLC磷酸化水平与肌动球蛋白解离度以及MRLC和肌球蛋白重链(myosin heavy chains,MHC)的降解呈负相关,宰后0.5~72.0 h内,MRLC磷酸化水平会随时间的延长而降低,MRLC-磷酸化可以促进肌动球蛋白的解离以及MRLC和MHC的降解来改善肉的嫩度。然而,值得注意的是,结构蛋白(肌动蛋白、肌钙蛋白-T和肌间线蛋白等)的磷酸化水平与肌原纤维的降解呈负相关,肌原纤维蛋白磷酸化可以提高肌原纤维蛋白的稳定性来降低钙蛋白酶对它的降解。Li Zheng等[42]的研究表明肌球蛋白重链、肌动蛋白、肌钙蛋白-T和肌间线蛋白等肌原纤维蛋白的磷酸化,会降低μ-钙蛋白酶降解肌纤维蛋白的敏感性,表明肌原纤维蛋白磷酸化不利于肉嫩度的改善。
1.4 小热休克蛋白对肉品质的影响
sHSPs是一种具有多种抗应激功能的伴侣蛋白,具有标记肉嫩度变化的潜力。迄今为止,哺乳动物中已鉴定出10 种sHSPs,但并非所有的sHSPs在哺乳动物中都普遍表达[43]。据报道,HSP20、HSP27和αβ-晶状体蛋白与肉的品质有关[43]。目前,动物宰后肌肉sHSPs表达和肉嫩度之间的关系尚存在两种相互矛盾的观点。Kim等[44]提出sHSPs浓度与牛肉的嫩度呈明显的负相关,这是因为sHSPs的伴侣功能可以维持蛋白质的完整性,不利于肌原纤维的降解。因此,HSP27和αβ-晶状体蛋白的下调有助于肌动蛋白和肌球蛋白的降解,从而产生嫩度较好的肉品[44]。相反,Morzel等[45]在金色阿基坦公牛背最长肌中却发现HSP27的表达与牛肉的嫩度呈正相关,这可能是由于HSP27可以阻止肌动蛋白的聚集,进而促进成熟过程中蛋白酶对肌动球蛋白的水解,他们发现HSP27在生鲜肉中的表达和HSP27片段在成熟后牛肉中的表达能解释91%的感官嫩度。事实上,一些研究已经推测sHSPs的宰后降解可能加速了细胞骨架蛋白的降解程度[46]。sHSPs降解后失去抗凋亡特性,可能会引起更多酶促蛋白水解的发生,提高了成熟嫩化的潜力,进而对肉的嫩度和多汁性产生正面影响。Ma Danyi等[46]研究发现,HSP27的降解和钙蛋白酶3的早期激活会导致牛腰背最长肌具有较高的嫩度和多汁性,且sHSPs的蛋白水解程度存在肌肉部位的特异性。
2 不同成熟方式对肉品质的影响
目前,肉类常用成熟方法主要有两种:干法成熟和湿法成熟。两种成熟方法均在成熟过程中显著改善了肉的嫩度和其他品质特性。近几十年来,随着长途跨国牛肉贸易的兴起,湿法成熟正逐步变得越来越流行。
2.1 干法成熟
干法成熟是将牛肉胴体、二分体、分割部位肉(未包装)在严格控制温度、风速和相对湿度的环境(冷藏室)中持续放置几周或几个月的成熟方法。对牛肉来说,干法成熟的最佳时间一般是14~21 d,长于湿法成熟(0~1 ℃下7~10 d)[2]。干法成熟后,肉的嫩度、风味和多汁性都得到了显著改善。
干法成熟对肉风味的改善尤为显著,长时间的成熟(超过28 d)会使肉品中与风味相关的挥发性化合物含量增加[47],且干法成熟的肉品在真空包装中贮藏19 d内风味都较稳定[48]。但成熟过程中往往会伴随着脂质氧化反应的发生,当牛肉的脂肪氧化后硫代巴比妥酸值超过2 mg/kg时,牛肉就会产生氧化味[49]。与湿法成熟相比,干法成熟14 d对产品适口性改善所带来的经济效益足以抵消干法成熟过程中的所出现的质量损失[2]。针对长期成熟过程中存在的脂质氧化和肌红蛋白氧化问题,研究人员正尝试通过饲喂VE营养强化剂来延缓成熟肉中的肌红蛋白氧化问题,进而提高产品肉色稳定性。Jose等[50]发现,添加3.5~4.0 mg/kg VE的羊肉在干法成熟条件下贮藏21 d后再进行托盘包装展示60 h,肉品表面仍具有令人满意的肉色,可以弥补成熟肉品在肉色稳定性方面的质量缺陷。
在嫩度改善方面,干法成熟至14~35 d时,牛肉的嫩度评分和剪切力虽然持续改善,但14 d之后的嫩度改善效果显著降低[51],因此干法成熟实际操作过程中应注意避免过长时间带来的牛肉干耗和企业耗能等经济损失。此外,干法成熟的肉品表面失水比较严重,肉品外观给人以汁液不足的感觉,事实上干法成熟期间肉品的多汁性得到改善[49]。DeGeer等[51]通过感官评定发现干法成熟的牛排的多汁性评分要高于湿法成熟,且干法成熟的牛排烹调后更加美味。
为进一步改善干法成熟的条件,提高肉的品质。最近,Kim等[52]首次尝试将干法成熟的处理条件(温度、相对湿度和风速)进行了优化处理,发现温度3 ℃、气流速率0.2 m/s、相对湿度49%条件下,成熟3 周的牛腰背最长肌适口性显著改善。这表明通过进一步优化干法成熟的条件,可实现肉类品质和企业生产效益的进一步提升。
2.2 湿法成熟
湿法成熟通常是将分割肉块真空包装后在特定条件下放置成熟,从而有利于肉品的加工、运输,并提高其产量,延长保质期。湿法成熟是目前肉品产业中最常用(95%的产品)的成熟方法[48]。与传统干法成熟相比,湿法成熟在一定程度上降低了肉的损耗,缩短了成熟时间,降低了生产成本和肉品价格,但许多消费者对湿法成熟肉品的喜好度偏低,因为湿法成熟的牛肉容易存在酸味和血腥味[51]。然而,一些学者却发现干法成熟和湿法成熟样品的风味差异较小或没有差异,且许多消费者对湿法成熟肉品的风味更加熟悉;而对于喜欢干法成熟肉品的消费者,他们可以接受干法成熟肉品较高的价格[53]。目前,为改善湿法成熟的不足,研究人员正尝试通过新型活性包装膜来替代传统的真空包装膜(图2)。Djenane等[54]就利用牛至精油活性真空包装膜进行湿法成熟,结果发现显著提高了牛肉的嫩度和产品的质量稳定性,并延长了货架期,这将是未来湿法成熟技术发展的一个新方向,可为肉品成熟提供更好的选择。
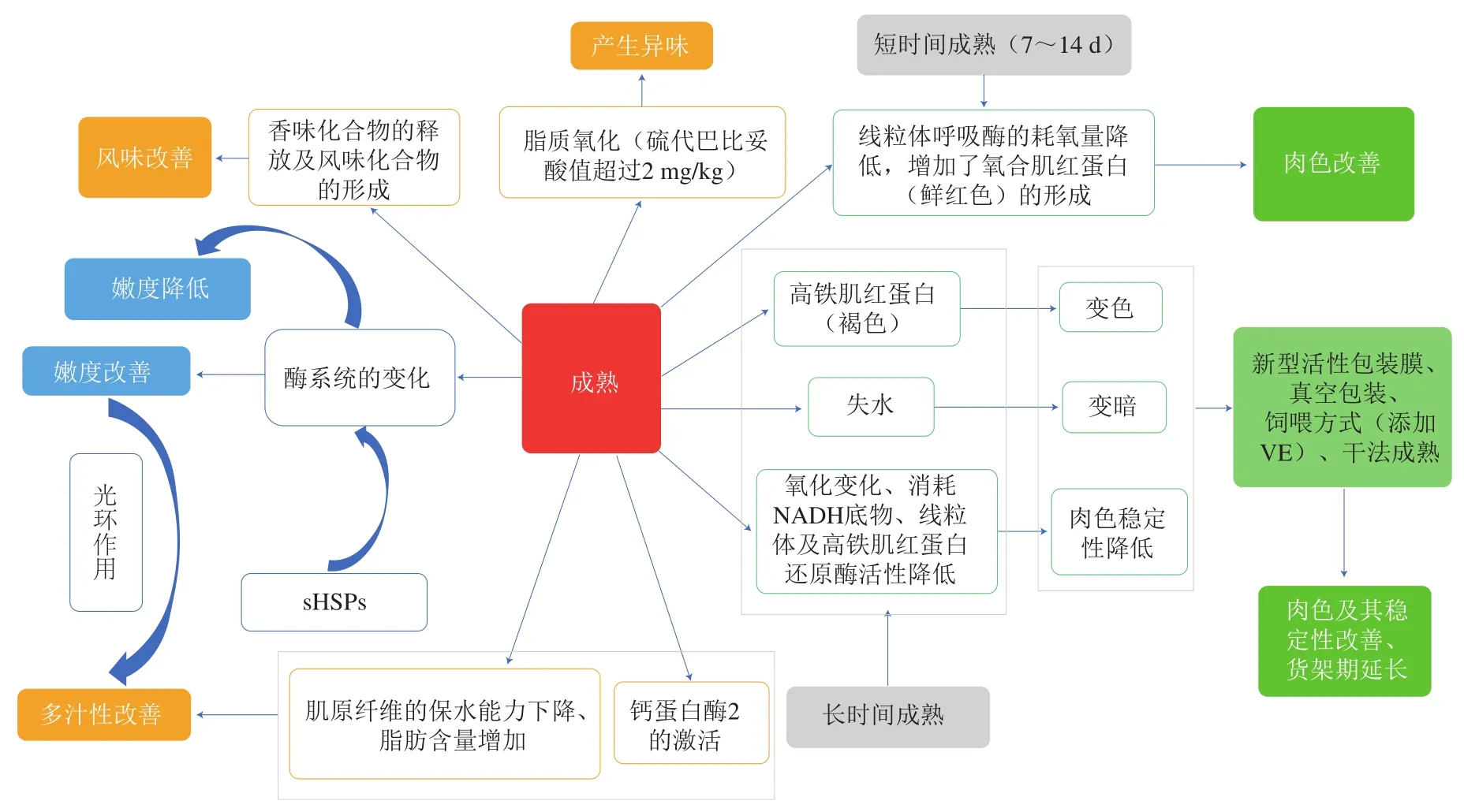
图2 成熟对肉类品质的影响[14,16,19,44,47,49-50,54]Fig. 2 Effect of aging on meat quality[14,16,19,44,47,49-50,54]
2.3 干湿法结合成熟技术
考虑到干法成熟成本高且耗时长,长时间的成熟对牛肉适口性的作用效果也会降低[2]。而湿法成熟技术虽然弥补了干法成熟成本上的缺陷,但湿法成熟的肉品有可能存在感官缺陷。因此,有必要对干、湿法成熟技术进行组合以弥补彼此的缺陷。干湿法成熟技术一般先将胴体进行干法成熟,然后再施加一段时间的湿法成熟处理。Kim等[55]研究发现,干湿法成熟(干法成熟10 d+湿法成熟7 d)可在保证牛肉感官品质(嫩度、多汁性、风味和整体喜爱度)和肉色与传统干法成熟肉差异不显著的前提下,显著提高成熟牛肉的保水性和嫩度。他们指出该方法缩短了干法成熟时间,节约了操作成本,从而使当地小型肉类加工企业受益。另外,Ha等[56]调查了不同成熟方法处理的澳洲牛肉在日本市场的受欢迎程度,发现与湿法成熟样品相比,干湿法成熟(湿法成熟21 d+干法成熟35 d)的牛排在嫩度和多汁性上与之并无显著差异,但在风味和总体评分上更高,该条件下成熟56 d后牛肉的L*值、b*值较低,而a*值较高,总体上更受日本消费者的喜爱。因此,干湿法结合成熟技术在提高肉品品质方面具有较好的应用前景,且可以在一定程度上缩短成熟时间,但还要进一步优化干湿法成熟技术的成熟时间,实现成熟肉品品质和经济效益的整体提升。
3 新型成熟技术
3.1 特殊包装干法成熟技术
在干法成熟过程中使用高透湿性包装袋(干法成熟包装袋)可以改进传统干法成熟无包装的状态,是一种新型的干法成熟方法。与传统的未包装干法成熟相比,使用干法成熟袋包装成熟的肉品不仅具有相同的感官品质,还能有效降低修整损失和微生物污染风险,降低了肉品对成熟环境的要求[48]。Ahnström等[57]研究发现传统干法成熟和新型袋装干法成熟在嫩度和多数感官性状方面并没有明显差异,这证明了该技术的有效性。而Lee等[58]的研究结果与大多数的报道不同,他们发现新型干法成熟包装袋并不能抑制微生物污染,并推测传统干法成熟使胴体表面脱水变硬,可能有效抑制了微生物的渗透繁殖,而这两种干法成熟方式在化学成分、腺苷酸、总游离氨基酸含量以及剪切力上均无显著差异。Lepper-Blilie等[59]也指出采用上述两种干法成熟方式的肉品在感官性状以及大肠杆菌和乳酸菌数量等方面差异均不显著,但干法成熟袋成熟牛肉的质量损失更低。因此,使用干法成熟袋既可以达到传统干法成熟对肉品品质的改善效果,又可降低肉品的干耗,是干法成熟较理想的改进技术。
3.2 差异化肌肉成熟技术
同一牛胴体的不同部位肉由于肌肉生化特性及肌纤维类型的不同,可能会导致牛肉成熟过程中发生不同程度的生化反应,最终造成嫩度、肉色和氧化稳定性的差异[60-61]。Nair等[61]发现不同成熟时间(0、7、14、21 d)对不同部位肌肉(背最长肌、腰大肌、半膜肌)的嫩化程度不同,成熟21 d内背最长肌的嫩度持续改善,而腰大肌只在成熟期前7 d内嫩度得到改善,对于半膜肌嫩度的改善则需要成熟14 d。Kim等[62]研究发现成熟期间不同肌肉类型之所以表现出不同的品质特性主要是由肌肉代谢类型的不同所致,他们确定了成熟过程中与不同肌肉的氧化稳定性有关的代谢物,如酰基肉碱、游离氨基酸、核苷酸和葡萄糖醛酸苷,可作为标记不同肌肉成熟差异程度的关键生物标签。因此,针对不同肌肉类型制定差异化成熟策略对牛肉产业的提升意义重大。而如何为不同部位肉品针对性设计科学合理的成熟时间及配套条件还有待进一步研究。
3.3 新型物理成熟技术
3.3.1 电离辐射技术
电离辐射作为一种有效控制肉品微生物的技术,能够很好地保证肉的新鲜度和营养品质。同时,电离辐射技术也属于一种物理成熟技术,一般流程是将屠宰后的牛肉切成牛排进行真空包装后,再进行不同辐射强度的照射,随即进行不同时间的成熟。但值得注意的是,目前电离辐射在肉嫩度、肉色等品质改善方面的研究结论不一致。Rowe等[63]发现经过6.4 kGy辐射后再成熟14 d会使生鲜牛肉中μ-钙蛋白酶活性下降,不利于肉的嫩化。相反,有研究者称电离辐射会破坏肌原纤维和溶解胶原分子,进而提高肉的嫩度,但辐射过程也会促进肉的脂质氧化、肉色劣变及异味的产生,产生负面影响[64]。同时,Kanatt等[65]研究发现,肉的嫩度与辐射强度呈正相关,且辐射后肉的a*值增加,2.5 kGy的辐射强度不仅改善了肉的嫩度,对其他感官特性也未产生不利影响,但此研究未对成熟后的辐射效果进行评估。Rodrigues等[66]最近研究了不同辐射强度(0、3、6、9 kGy)和不同成熟时间(1、14 d)对真空包装牛肉品质的影响。结果发现3 kGy和6 kGy辐射强度下牛肉的蛋白水解程度降低,剪切力较高,而高辐射强度(9 kGy)促进了胶原蛋白的溶解,降低了样品的剪切力,但9 kGy照射下样品的高铁肌红蛋白含量较高,出现明显的褐变。因此,作为肉类辅助成熟技术,还需要进一步探究不同辐射参数及其与成熟时间的组合对肉类的成熟效果,探索最优组合方案。
3.3.2 冷冻-冷藏结合的成熟技术
最新研究发现冷冻/解冻和湿法成熟技术相结合可以有效改善冷冻对肉品的负面影响,在抑制微生物的同时,显著提高肉品的嫩度,这可能是冰晶破坏了肌原纤维结构加之冷冻条件下降低了钙蛋白酶抑制剂活性的协同作用所致[64]。
冷冻-冷藏结合的成熟技术(freezing/chilling and aging,FCA)处理在成熟早期即可改善肉的嫩度。与常规湿法成熟相比,将屠宰后的牛肉立即于-20 ℃冷冻40 d后,再在4 ℃放置24 h解冻,随即转入(1.0±0.5)℃下成熟21 d,可在成熟第1周时显著改善安格斯牛肉的嫩度,使其具有常规成熟14 d时的嫩度,但这个处理对内洛牛肉的嫩度并没有改善效果,所以其改善效果因动物品种而异[67]。近期研究发现冷冻/冷藏前进行充分的成熟(aging prior to freezing and chilling,AFC)也可以产出品质更优良的冻肉,且冷冻/冷藏肉品的劣变程度会受到冷冻速率的影响[68]。Kim等[69]研究了成熟和冷冻顺序及冷冻速率对肉品质的影响,发现FCA处理的猪背最长肌的剪切力与AFC处理相同,甚至更低,但FCA的解冻损失较大,因此建议采用AFC结合快速冷冻技术来有效克服冷冻肉的质量缺陷。鉴于冷冻可以纳入正常的商业贮运过程,因此,将冷冻和成熟技术相结合可以在充分利用现行生产规则前体下改善肉的品质,特别是嫩度,但还需要对解冻条件作进一步的研究,以明确成熟、冷冻顺序和参数对肉类品质的影响。
3.3.3 高压技术
高压技术在食品工业中主要用于微生物的灭活和肉嫩度的提高。然而,高压(>400 MPa)诱导的蛋白质变性会导致肉品的持水能力下降、结构改变、氧化变质和变色等品质缺陷[70]。据报道,适当加压(100~200 MPa)会使各方面肉品质得到提升,如更好的水合性质和颜色稳定性[70]以及更高的酶活性[71]。高压可能会通过提高蛋白酶的活性,进而影响肉品的成熟程度和速率。尽管有很多研究报道了适度加压对肉理化性质的有利影响,但对高压处理肉成熟后品质的报道则较少。Hwang等[72]最近研究了高压处理(0.1、150、400 MPa)和不同成熟方法(干法、湿法)对猪背最长肌品质特性的影响,研究发现在400 MPa处理对干法成熟猪肉的嫩度没有影响,但显著降低了湿法成熟的猪肉背最长肌的剪切力。150 MPa处理的湿法成熟样品剪切力最低,且风味与未高压处理的样品无差异。因此,150 MPa处理后加湿法成熟技术在提高肉品嫩度和多汁性方面具有潜在的应用价值,但在消费者的喜爱度方面还需要作进一步的研究。
3.3.4 脉冲电场技术
脉冲电场技术(pulsed electric field,PEF)是一种很有前途的食品保藏和改善食品质量的非热能技术,该技术是将电场强度为0.1~80.0 kV/cm的电场脉冲(持续几纳秒到几毫秒)应用到食品中[73],目前已在一些国家的食品加工中得到商业化应用。成熟前使用脉冲电场可以使肌细胞膜通透性增强,迫使细胞器释放钙离子,从而激活钙蛋白酶,促进肌肉的嫩化;还能造成肌纤维的物理性破坏;促进肌钙蛋白T和肌间线蛋白的水解[74],这些都可以增加肌肉的嫩度。目前关于PEF对牛肉嫩化方面的研究结果还存在争议,肌肉类型、电场强度和成熟时间都会影响PEF改善肉嫩度的潜力。虽然许多研究报道了PEF在牛肉成熟过程中对肌肉嫩度的改善作用,但这些研究没有提供任何关于酶机制的实验证据。Bhat等[74]发现脉冲电场提高了成熟过程中牛股二头肌钙蛋白酶-2的活性,以及肌间线蛋白和肌钙蛋白T的降解程度,但是PEF对成熟期间(1、7、14 d)肌肉的剪切力并没有显著影响,值得注意的是,肉品在成熟前7 d的嫩化速度较快,可能有助于缩短成熟时间,这为PEF诱导嫩化的酶学理论基础提供了一定的技术支撑。
4 结 语
宰后早期肌肉向肉转化过程中会发生复杂的生理生化变化,这些变化从不同方面影响着肉品的最终品质特性。磷酸肌酸的分解、糖酵解和氧化磷酸化进程等能量代谢过程会影响pH值、ATP和代谢物水平以及肌原纤维的性质等;蛋白翻译后修饰会调节宰后能量代谢、Ca2+释放、肌肉收缩和细胞凋亡等过程;宰后酶系统(钙蛋白酶系统、Prdx6、半胱天冬蛋白酶介导的细胞凋亡过程)则会影响结构蛋白的降解、细胞凋亡过程等;而sHSPs在维持蛋白完整性,降低肌原纤维降解方面作用明显,这些生化过程都会对肉的嫩度、保水性、肉色及其稳定性等产生重要的影响。因此,探明并调控宰后的这些生化变化进程可以有效地改善肉的品质。
成熟可以改善肉的嫩度、风味和多汁性等品质,但是目前干、湿法成熟技术还存在各自的局限性,其成熟效果也会受到各种加工条件的限制。这意味着可以继续优化成熟方案,并在成熟过程中引进新兴的物理技术,如冷冻/解冻、电离辐射、高压以及脉冲电场等,克服传统干法成熟技术的成本缺陷和湿法成熟中存在的感官缺陷等,生产出消费者更满意的产品。但是,这些新型技术尚不成熟,还需要进一步的研究与优化,这将是未来肉类科学研究的热点之一。同时,新兴智能监测技术在肉品中的应用也将进一步加快对肉类成熟技术和肉类品质的持续改善。
猜你喜欢
选煤技术(2022年2期)2022-06-06
选煤技术(2022年1期)2022-04-19
肉类研究(2022年3期)2022-04-12
建材发展导向(2022年1期)2022-03-08
科技研究·理论版(2021年20期)2021-04-20
伴侣(2018年6期)2018-06-27
肉类研究(2017年10期)2017-11-20
猪业科学(2017年12期)2017-01-16
肉类研究(2016年6期)2016-07-22
肉类研究(2016年4期)2016-05-17

- 食品科学的其它文章
- 天然抗氧化剂对鱼糜及鱼糜制品抗氧化能力及品质影响的研究进展
- 茶多酚对小麦淀粉理化特性和面包品质的影响及机理
- 运动对苏尼特羊脂肪代谢和肉品质的影响
- 升温方式对鲢鱼肌球蛋白结构和理化性质的影响
- Thermal Unfolding and Aggregation Process of Recombinant Staphylococcal Enterotoxin M (rSEM) Associated with Potential Toxin Responsible for Staphylococcal Food Poisoning
- 小米麸皮水溶性膳食纤维-Cr(III)配合物的合成、表征及其体外抗氧化活性