
不同土壤pH 和磷水平下黄腐酸对番茄产量和根际土壤微生态的影响
2021-12-02 12:59张丽丽李继蕊毕焕改史庆华
中国蔬菜 2021年11期
张丽丽 李继蕊 毕焕改 史庆华 巩 彪*
(1 山东农业大学园艺科学与工程学院,作物生物学国家重点实验室,农业农村部黄淮海地区园艺作物生物学与种质创新重点实验室,山东泰安 271018;2 东营市农业农村局,山东东营 257091)
磷(P)是植物生长发育必不可缺的大量元素之一,P 肥在提升作物产量、改善产品品质方面起着至关重要的作用。P 肥施入土壤后在物理、化学反应和生物因素的复杂作用下极易形成难溶性的磷酸盐,继而被微生物固持或被土壤矿物吸附,使得当季作物的P 利用率仅为10%~25%(唐晓乐 等,2012)。我国北方干旱、半干旱地区分布着大面积碱性土壤(pH >7.3),因土壤剖面含有较多的碳酸钙或碳酸氢钙等石灰性物质,又被称作石灰性土壤(冯君 等,2019)。碱性土壤会进一步造成P 元素沉淀,降低作物P 肥利用效率,引起作物生育障碍(Yan et al.,2019)。因此,研究和开发土壤P活化技术既有利于提高P 肥利用效率,又有利于减少P 矿的开采和加工、P 肥流失造成的土壤污染,对维持农业可持续发展和生态平衡具有重要意义。
土壤是一个复杂的生态系统,植物与微生物互作影响土壤的理化性质,进而影响土壤P 素的状态。植物根系通过分泌碳水化合物,增加根际微生物种类和数量(熊明彪 等,2002)。根系和根际微生物均可分泌磷酸酶和有机酸,磷酸酶能降解土壤中的有机磷,如磷脂、糖脂和核酸等,使其转变为无机磷,而有机酸使土壤中的难溶性磷转化为可溶性磷,两者协同,促进土壤P 素的活化(李荣坦 等,2016)。但碱性土壤pH 高,易板结、含氧量低,不利于根系和根际微生物生长。因此,促进根系生长和改善根际微生态成为提高作物P 肥利用率的重要途径。黄腐酸(fulvic acid,FA)作为一种可溶于水的腐植酸,其分子量低、功能团密集、生理活性较强,被广泛用于农业领域。前人研究表明,土壤添加腐植酸或FA 能有效促进植物根系生长,增加侧根数量,提高根毛密度(谢尚强 等,2019)。且腐植酸能显著提高土壤微生物的多样性和数量,增加有益矿质营养活化的脲酶、磷酸酶等土壤酶活性(袁婉潼,2016)。此外,笔者通过水培番茄研究发现,即使不考虑土壤对P 的固定效果,FA 也能通过增强番茄根系泌酸,增加P 素利用率,促进低P 胁迫下幼苗的生长(张丽丽 等,2018)。
虽然关于FA 对根系发育和土壤微生物影响的研究已有文献记述。但不同土壤pH 和P 水平对番茄根际微生态的影响机理尚不明确。而根际微生态是根系-微生物-土壤相互作用微域的生态环境,根际微生态的健康与否决定了P 素的供应强度和有效性。因此,通过研究增施FA 对番茄植株生长和产量、根际微生物数量和群落结构、根际重要土壤酶活性的影响,阐明FA 对不同土壤pH 和P 水平下根际微生态的作用,可为碱土改良和P 肥增效提供理论依据和技术支撑。
1 材料与方法
1.1 试验设计
试验于2019 年2——8 月在山东农业大学园艺实验站进行。采用盆栽方式,于高40 cm、上下口径为45 cm 和38 cm 的大陶盆中填装15 kg 壤土,土壤pH 值为6.5,有机质含量8.1 g·kg-1,碱解氮含量113.6 mg·kg-1,速效磷含量61.5 mg·kg-1,速效钾含量90.8 mg·kg-1。以供试土壤为基础,以K2CO3和KHCO3混合盐(1V∶1V)调整土壤pH 值至8.0,作为碱性土壤,记作pH=8.0;通过添加等量KCl 来抵消配制碱土引入的钾元素误差,作为供试中性土壤,记作pH=6.5。由于土壤pH 经常变化,本试验每隔7 d 测定1 次土壤pH 值,确保土壤pH 稳定在8.0 左右。参照张彦才等(2008)的方法,以0.53 g·kg-1的P 肥(按P2O5计)施入量作为番茄常规施P 水平,设置施磷(+P)和不施磷(-P)处理。参照笔者前期试验筛选的FA(山东泉林嘉有肥料有限责任公司)浓度(张丽丽 等,2018),设置添加0.08 g·kg-1FA(+FA)和不添加FA(-FA)处理。将土壤pH、P肥和FA 处理两两组合,共计8 个处理,分别为T1(pH=6.5、-P、-FA)、T2(pH=6.5、+P、-FA)、T3(pH=8.0、-P、-FA)、T4(pH=8.0、+P、-FA)、T5(pH=6.5、-P、+FA)、T6(pH=6.5、+P、+FA)、T7(pH=8.0、-P、+FA)、T8(pH=8.0、+P、+FA)。供试番茄(Solanum lycopersicumL.)品种为罗拉,幼苗购自山东安信种苗股份有限公司。定植于各处理大陶盆中,每盆1 株。试验采用随机区组设计,3 次重复,每重复9 盆,合计216盆。开花后50 d 左右,每株留6 穗果摘心,田间水、肥及病虫害管理参照常规方法进行。
1.2 番茄植株生物量、产量和P 吸收量测定
记录植株开始生长至采收结束后的根、茎和叶片(包含摘除的老叶)的鲜质量,作为植株生物量。产量采取单株测产。采收结束后将整株植株烘干至衡重,称量记录干质量;再将干样研磨粉碎,采用钼锑抗比色法测定单株P 含量(Gong et al.,2014a)。
P 吸收量=单株干质量× P 含量
1.3 根际微生态分析
于番茄盛果期,采用抖动法收集根际土壤(唐晓乐 等,2012)。采用稀释平板法测定根际微生物数量,以牛肉膏蛋白胨培养基培养细菌,以马丁氏培养基培养真菌,以高氏1 号培养基培养放线菌(巩彪 等,2016)。采用次氯酸钠-苯酚钠比色法测定脲酶活性,采用3,5-二硝基水杨酸比色法测定蔗糖酶活性,采用磷酸苯二钠法测定磷酸酶活性,采用高锰酸钾滴定法测定过氧化氢酶活性(周礼恺和张志明,1980)。
将根际土壤样品交由南京诺唯赞生物科技有限公司,采用16S rDNA 测序技术测定并分析根际细菌多样性,Miseq 测序完成后,将得到的原始下机数据,利用overlap 将双端数据进行拼接,并进行质控、嵌合体过滤,获得高质量的有效数据;再将获得的有效数据进行97%的相似度聚类,过滤singleton 序列以降低假阳性率,获得最终的OTU丰度及代表序列,进一步进行多样性分析、物种分类注释和差异分析等(http://www.vazyme.com/)。
1.4 数据处理
所有试验结果均进行3 次生物学重复,采用SPSS 软件进行差异性检验和相关性分析。
2 结果与分析
2.1 不同土壤pH 和P 水平下FA 对番茄植株生物量、产量和P 吸收量的影响
图1 表明,与中性土壤相比,碱性土壤显著降低了番茄的植株生物量、单株产量和P 吸收量。在两种土壤pH 条件下,增施P 肥均能显著提升植株生物量、单株产量和P 吸收量,但这3 项指标在碱性土壤条件下的增幅均小于中性土壤。说明P肥对促进番茄生长和产量起到重要作用,但是碱性土壤会降低P 肥的有效性,限制番茄植株生长和产量形成。土壤添加FA 可有效提升植株生物量、单株产量和P 吸收量,且FA 对单株产量和P 吸收量的促进效果在“pH=6.5、-P”和“pH=8.0、+P”条件下更加明显。表明FA 能促进番茄植株生长和P 素吸收,在低P 中性土壤条件下增强土壤P的活化,在高P 碱性土壤条件下减少P 的固定。此外,植株生物量、单株产量和P 吸收量在T1(pH=6.5、-P、-FA)与T4(pH=8.0、+P、-FA)、T5(pH=6.5、-P、+FA)与T8(pH=8.0、+P、+FA)间差异不显著,表明在碱性土壤条件下增施P 肥和FA 能缓解碱性土壤对番茄植株生长产生的抑制作用。
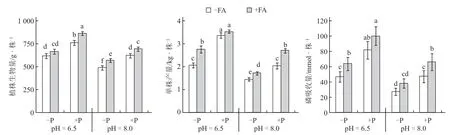
图1 不同土壤pH 和P 水平下FA 对番茄植株生物量、产量和P 吸收量的影响
2.2 不同土壤pH 和P 水平下FA 对番茄根际细菌、真菌和放线菌数量的影响
表1 显示,碱性土壤有利于真菌数量的提升;增施P 肥对真菌数量的影响不显著;中性土壤添加FA 对真菌数量产生促进效果,而碱性土壤添加FA 对真菌数量产生抑制作用。细菌与放线菌数量变化趋势相同:在其他处理条件相同的情况下,碱性土壤降低细菌和放线菌数量;增施P 肥和FA 均有利于细菌和放线菌数量的提升。值得注意的是,T2(pH=6.5、+P、-FA)与T5(pH=6.5、-P、+FA)间的细菌和放线菌数量差异不显著,表明在中性土壤条件下添加FA 对细菌和放线菌数量的影响上可与增施P 肥产生对等效应。T7(pH=8.0、-P、+FA)的细菌和放线菌数量显著高于T4(pH=8.0、+P、-FA),表明在碱性土壤条件下添加FA对细菌和放线菌数量的影响上优于增施P肥。此外,T8(pH=8.0、+P、+FA)分别与T1(pH=6.5、-P、-FA)、T2(pH=6.5、+P、-FA)间细菌和放线菌数量差异不显著,说明在碱性土壤条件下增施P 肥和FA 能缓解碱性土壤对细菌和放线菌数量的抑制作用,且FA 的增效比P 肥更强。
2.3 不同土壤pH 和P 水平下FA 对番茄根际细菌多样性的影响
Alpha 多样性是评估群落生态中物种丰富度的有效指标,可综合反映物种丰富度和均匀度(丁建莉 等,2016)。由Alpha 稀释曲线可知(图2),各处理根际细菌多样性由高到低的排序为:T6(pH=6.5、+P、+FA)>T5(pH=6.5、-P、+FA)≈T2(pH=6.5、+P、-FA)>T1(pH=6.5、-P、-FA)>T8(pH=8.0、+P、+FA)>T7(pH=8.0、-P、+FA)>T4(pH=8.0、+P、-FA)>T3(pH=8.0、-P、-FA),与表1 细菌数量排序类似,表明碱性土壤降低细菌多样性,增施P 肥和FA 可以提升细菌多样性;图2中细菌多样性T5(pH=6.5、-P、+FA)≈T2(pH=6.5、+P、-FA),而表1 中T2(pH=6.5、+P、-FA)与T5(pH=6.5、-P、+FA)的细菌数量差异不显著,也表明细菌多样性和数量之间呈正相关。综上,中性土壤条件下,添加FA 与施用P 肥对细菌多样性影响效果相似,而碱性土壤添加FA 对细菌多样性的提升效果优于增施P 肥,即FA 可能通过活化土壤P 素提高根际细菌多样性。

图2 不同土壤pH 和P 水平下FA 对番茄根际细菌Alpha多样性的影响
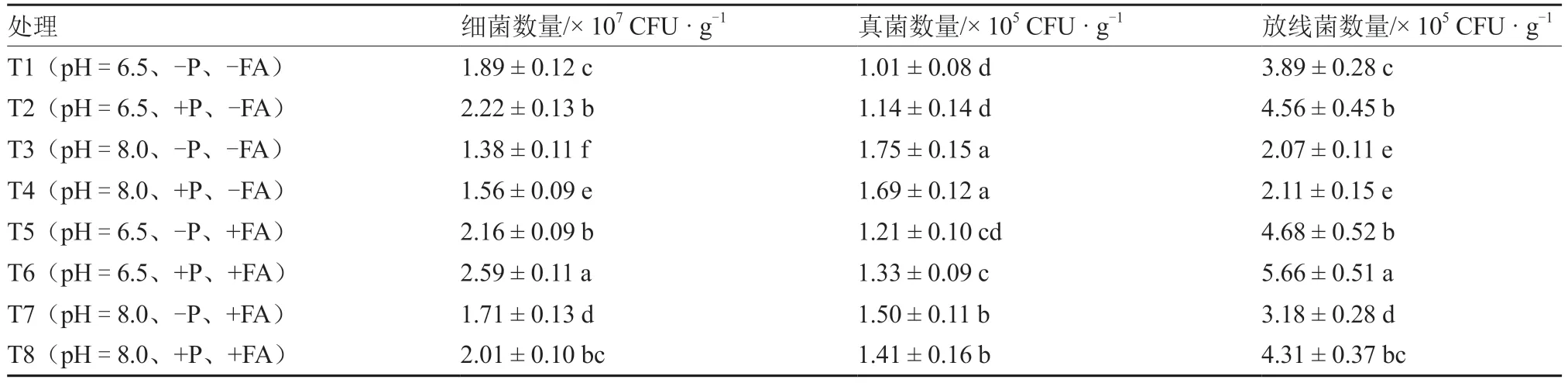
表1 不同土壤pH 和P 水平下FA 对番茄根际细菌、真菌和放线菌数量的影响
主成分(PC)分析是将多个相关变量转化成少数几个能够反映初始全部指标大部分信息的综合变化形式的一种多元统计分析方法(王春萍 等,2017)。图3 显示了处理之间的差异,不同处理间的距离代表其物种组成的差异情况。第1 主成分和第2 主成分的贡献率分别为71.89%和15.04%,总贡献率为86.93%。图中实线圈内的T1(pH=6.5、-P、-FA)、T3(pH=8.0、-P、-FA)、T5(pH=6.5、-P、+FA)、T7(pH=8.0、-P、+FA)均为-P 处理,居于横坐标PC1 的右半部分,表明PC1 可归纳为P 肥效应,由左至右表现为P肥效应降低。而虚线圈内的T3(pH=8.0、-P、-FA)、T4(pH=8.0、+P、-FA)、T5(pH=6.5、-P、+FA)、T7(pH=8.0、-P、+FA)、T8(pH=8.0、+P、+FA)中,除T5(pH=6.5、-P、+FA)外均为碱性土壤处理,居于纵坐标PC2 的上半部分,表明PC2 可归纳为土壤pH 效应,由下至上表现为碱性效应增强。T1(pH=6.5、-P、-FA)与T5(pH=6.5、-P、+FA)、T2(pH=6.5、+P、-FA)与T6(pH=6.5、+P、+FA)、T3(pH=8.0、-P、-FA)与T7(pH=8.0、-P、+FA)、T4(pH=8.0、+P、-FA)与T8(pH=8.0、+P、+FA)4 对处理组间差异为FA。图中4 个箭头所示的方向即为添加FA 后各对比组细菌多样性的PC 变化,其中3 个箭头自右上指向左下,1 个箭头自左上指向右下,表明FA降低了土壤碱性效应,增强了P 肥效应。此外,T3(pH=8.0、-P、-FA)与T7(pH=8.0、-P、+FA)、T4(pH=8.0、+P、-FA)与T8(pH=8.0、+P、+FA)处理组的箭头长度均大于其余2 对处理组,表明FA 在碱性土壤条件下对P 肥的增效更为明显。
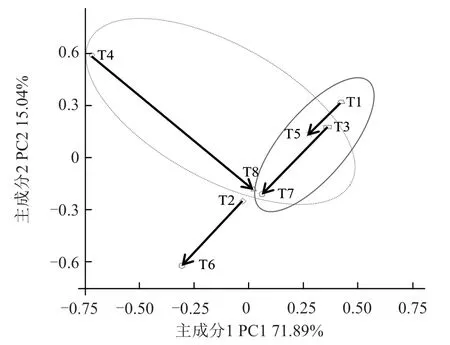
图3 不同土壤pH 和P 水平下FA 对番茄根际细菌多样性的主成分分析
图4-A 为细菌多样性在“科”水平上的聚类分析结果,T1(pH=6.5、-P、-FA)与T5(pH=6.5、-P、+FA)、T3(pH=8.0、-P、-FA)与T7(pH=8.0、-P、+FA)各被聚为一个小类,表明无P 肥施入的情况下,仅添加FA 对细菌多样性的影响很小。T8(pH=8.0、+P、+FA)介于T1(pH=6.5、-P、-FA)和T2(pH=6.5、+P、-FA)之间,说明碱性土壤配施P 肥和FA 能使根际土壤细菌多样性趋于不施FA 的中性土。图4-B 为细菌多样性在“科”水平上的前20 种细菌及其占有比例,本试验8 个处理的细菌多样性在“科”水平上与前20种细菌完全一致,包含:叶绿体菌科(Chloroplast)、鞘脂单胞菌科(Sphingomonadaceae)、黄色单胞菌科(Xanthomonadaceae)、Gp4、Gp6、Gp16、绿湾菌科(Chloroflexi)、红螺菌科(Rhodospirillaceae)、芽单胞菌科(Gemmatimonadetes)、根瘤菌科(Rhizobiales)、放线杆菌科(Actinobacteria)、微单孢菌科(Micromonosporaceae)、芽单胞菌科(Gemmatimonadaceae)、高温单孢菌科(Thermomonosporaceae)、糖杆菌科(Saccharibacteria)、链霉菌科(Streptomycetaceae)、红螺菌科(Rhodospirillales)、生丝微菌科(Hyphomicrobiaceae)和变形菌科(Alphaproteobacteria)。T6(pH=6.5、+P、+FA)为中性土壤配施P肥和FA,其植株长势、单株产量和微生物数量均为最优,且此处理下各类细菌所占比例最为均匀。而碱性土壤〔T3(pH=8.0、-P、-FA)、T7(pH=8.0、-P、+FA)、T8(pH=8.0、+P、+FA)〕则促使叶绿体菌科和鞘脂单胞菌科成为优势菌科,添加FA 可降低碱土造成优势菌科所占的比例。
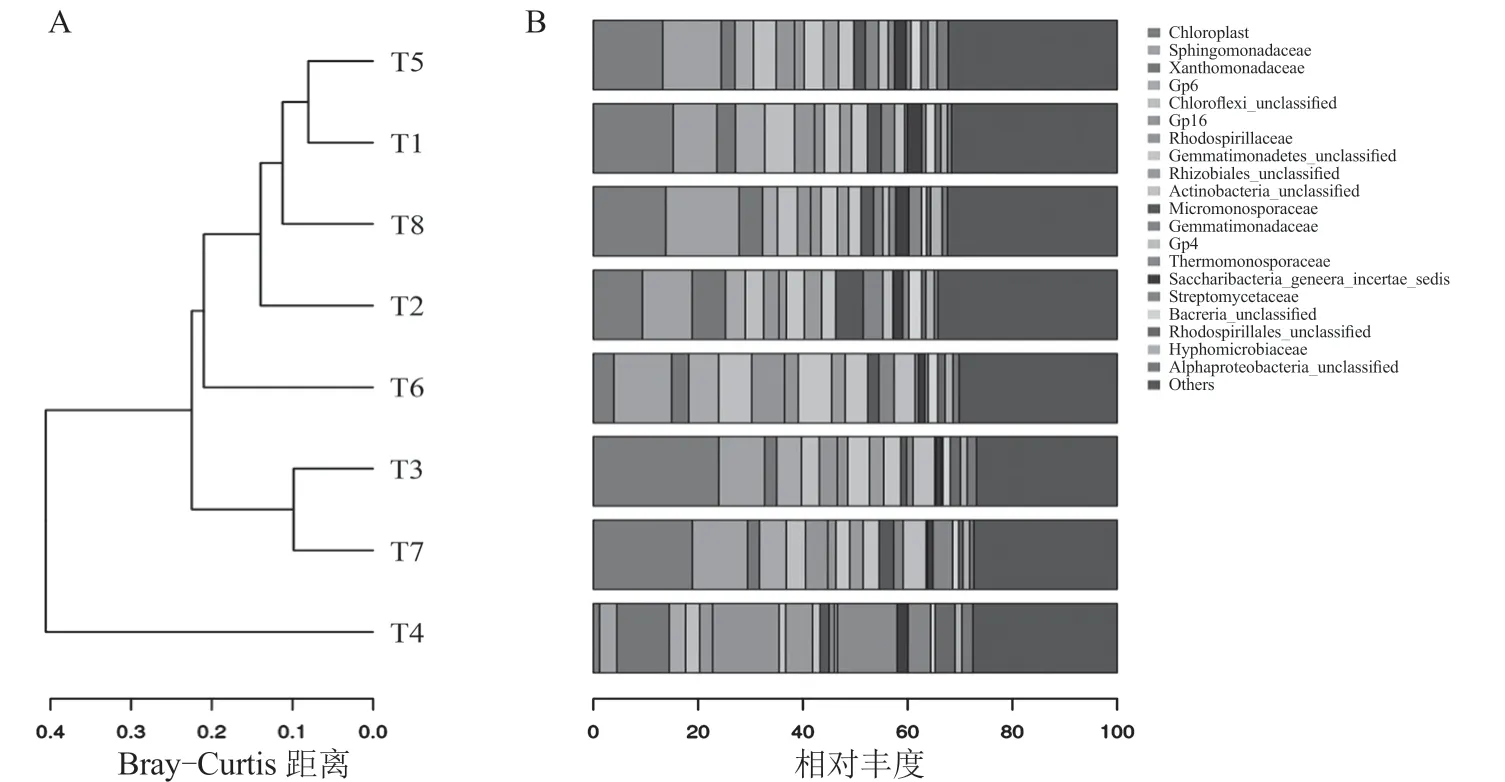
图4 不同土壤pH 和P 水平下FA 对番茄根际细菌多样性的聚类分析结果
2.4 不同土壤pH 和P 水平下FA 对番茄根际土壤酶活性的影响
表2 显示,脲酶和磷酸酶活性在各处理间呈现类似的变化趋势。碱性土壤降低脲酶和磷酸酶活性,而增施P 肥和FA 均能提升脲酶和磷酸酶活性。T2(pH=6.5、+P、-FA)与T5(pH=6.5、-P、+FA)、T1(pH=6.5、-P、-FA)与T8(pH=8.0、+P、+FA)间脲酶和磷酸酶活性差异均不显著,表明FA 能提高P 肥对脲酶和磷酸酶活性的促进效果,且碱性土壤增施P 肥和FA 能使脲酶和磷酸酶活性达到未施肥的中性土壤水平。碱性土壤处理下的蔗糖酶活性显著低于中性土壤,增施P 肥仅在碱性土壤添加FA 的处理〔T7(pH=8.0、-P、+FA)、T8(pH=8.0、+P、+FA)〕间产生显著差异,但在其他处理相同条件下添加FA 均能显著提升蔗糖酶活性。过氧化氢酶在碱性土壤条件下活性上升,增施P 肥和FA 显著降低了过氧化氢酶活性。
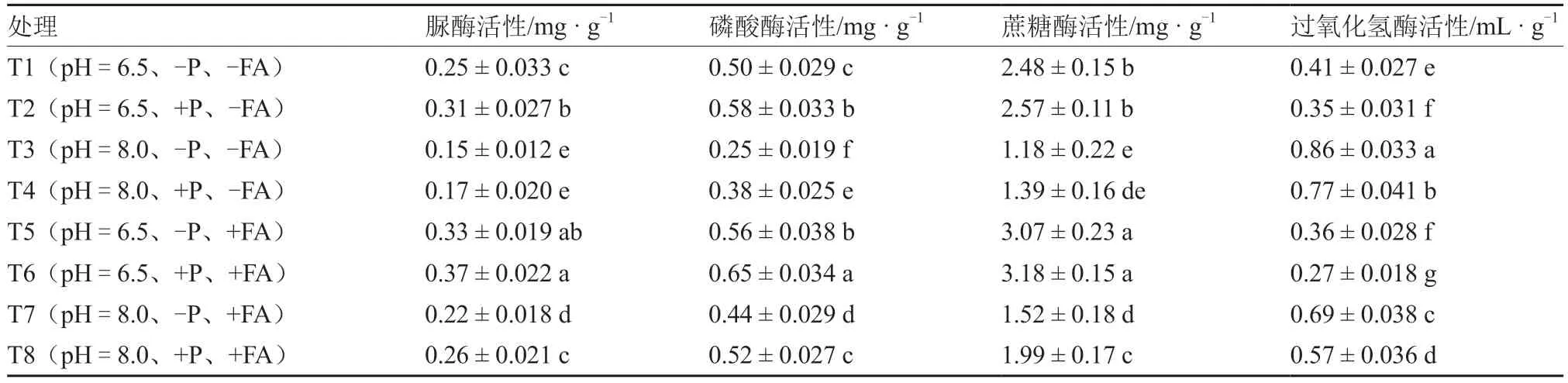
表2 不同土壤pH 和P 水平下FA 对番茄根际土壤酶活性的影响
3 讨论
碱胁迫不仅对作物根系发育造成直接伤害,还会因土壤易板结造成根系缺氧,且P、Ca、Fe、Mg 等矿质元素易沉淀造成根系营养障碍等间接伤害(Gong et al.,2013,2014b)。因此,适量补充易沉淀元素有利于降低碱性土壤对植物生长的抑制作用,这在本试验中添加P 肥后的效果可见一斑。FA 能显著提升P 肥对番茄植株生长和产量形成的作用,提高番茄植株对P 素的吸收量。这可归因于下述3 条有益效果:首先,经FA 处理的植物与外源使用生长素或赤霉素处理的植物具有类似的生长特性,即促进植株尤其是根系的生长,因而FA 又被认定为一种“生物刺激素”(梅慧生 等,1980;周霞萍 等,2006)。根系施用0.08 g·kg-1的FA 能显著提高番茄根系生长量(张丽丽 等,2018),这有利于提高根系在土壤耕层的覆盖度,进而提升P吸收量(韩玉国 等,2004)。其次,FA 分子吸附能力强,能迅速吸附土壤溶液中可溶态P,降低速效态P 在土壤耕层中的流失(刘方春 等,2008),这可能是本试验中FA 配施P 肥效果优于单施FA 的重要原因之一。最后,FA 能影响植物根系分泌物,促进有机酸和磷酸酶外泌(张丽丽 等,2018);同时,FA 可影响根际微生物群落,形成更有利于植物矿质营养吸收和微生物生存的植物-微生物互惠共生关系(唐晓乐 等,2012)。
根际微生物的数量和种类在根际微生态中发挥着重要的作用,在无机盐和有机质等营养物质的转化中也发挥着重要功能(谷岩 等,2012)。腐殖质和根系分泌物是根际微生物的主要碳源(付学琴和黄文新,2014)。碱性土壤抑制植株生长,降低根系分泌物总量,因而根际细菌和放线菌数量(表1)、根际细菌多样性均低于中性土壤(图1)。碱性盐易引起根系死亡(Gong et al.,2015),死亡的根系在土壤中腐解可引起真菌数量上升,这可能是本试验中碱性土壤处理下根际真菌数量高于中性土壤的原因;而碱性土壤增施P 肥和FA 能降低真菌数量,增加植株生物量。已有研究表明,FA 能改变土壤生态系统中细菌、真菌、放线菌的数量,并促进有益菌群的繁殖(张福建 等,2019)。本试验发现在不同pH 和P 水平下增施FA 均能显著增加细菌和放线菌的数量,降低真菌数量,且这种调节作用在碱性条件下更突出(表1)。表明FA 能改善不良土壤环境下的微生物菌群失调,使土壤的微生物群落维持良性发展,促进番茄植株生长和对P素的吸收。
细菌是土壤微生物系统中数量最庞大的种群,对维持整个土壤生态系统的稳定、促进植物生长和矿质营养吸收起着重要的作用(Girvan et al.,2005)。通过16S rDNA 测序技术对番茄根际细菌进行多样性测序分析可知,碱性土壤会引起高温单孢菌科等有害细菌种群所占的相对比例升高,降低Gp16 和链霉菌科等部分有益菌(图4)。在烟草中的研究表明,Gp16 和链霉菌科细菌可以有效拮抗根系真菌性病害,促进根系健康生长(冯吉 等,2016)。施加FA 后,鞘脂单胞菌科细菌所占的相对比例显著上升,糖杆菌科细菌所占的相对比例显著下降。鞘脂单胞菌科细菌能抵抗如枯萎病菌、青枯病菌等多类真菌性病害,提高根系的呼吸和代谢能力(向立刚 等,2019)。前人研究表明,糖杆菌科细菌的数量与植株的P 吸收量成反比,因此降低糖杆菌科细菌的相对含量能有效促进植物对P 素的吸收(李冰 等,2019),这与本试验中番茄植株生长量、对P 素吸收量和微生物变化的趋势相吻合。根际有益细菌种群在碱性土壤条件下急剧下降,导致植株面临非生物和生物交叉胁迫,增加根系病害和坏死程度,造成有害真菌数量上升。而施加FA 后,增加的有益菌群促进根系呼吸和物质代谢,加速了有机酸和磷酸酶的分泌,这不仅有利于建立根际微生物群落多样性,还有利于难溶态和有机态P 素活化,增强植物对土壤P 素的吸收和利用(王若楠 等,2019)。而FA 可有效改良土壤的微生物群落结构,这也是FA 能提高P 素利用和促进植株生长的重要原因。根际土壤酶主要来源于植物根系分泌和微生物释放,微生物数量和种类对土壤酶的构成和活性影响显著(刘丽英 等,2019)。研究表明,脲酶、磷酸酶和蔗糖酶活性与土壤生物学活性、土壤肥力、土壤生产力呈正相关,这些土壤酶活性越高,土壤越肥沃(林耀奔 等,2019)。而碱性土壤对根系和微生物的不良影响会加速生物体内的氧化胁迫,致使过氧化氢酶活性上升,这是生物界普遍存在的防御机制(周礼恺和张志明,1980)。本试验中,土壤中添加FA 后,提升了番茄根际细菌和放线菌数量,增加细菌多样性并平衡种间数量关系,这些结果均有利于土壤脲酶、磷酸酶和蔗糖酶活性的提升。
4 结论
FA 可有效提高番茄植株对P 素的吸收量、促进植株生长和产量形成;且增施P 肥配合FA 添加不仅可恢复弱碱性土壤的番茄生产能力,亦可显著提升P 肥利用效率。这可能与FA 能提高细菌、放线菌数量、细菌多样性和土壤脲酶、磷酸酶和蔗糖酶活性密切相关。本试验结果表明,FA 与P 肥具有相似的根际微生态调控特征,这间接表明FA 能协同增强P 肥利用率,促进根系生长调控根际微生态。
猜你喜欢
现代园艺(2022年19期)2022-09-21
家庭科学·新健康(2022年7期)2022-07-13
中国土壤与肥料(2022年4期)2022-06-14
林业科学(2022年2期)2022-05-11
草地学报(2022年4期)2022-04-25
表面技术(2022年1期)2022-02-12
四川农业科技(2021年7期)2021-12-01
小读者(2021年19期)2021-11-07
作文评点报·中考版(2019年4期)2019-02-18
课外语文·中(2018年10期)2018-11-16
