
半干旱区油松林分夜间液流变化特征及其影响因子
2021-11-08 08:55徐志彬陈胜楠陈立欣刘清泉刘平生张志强
中国水土保持科学 2021年5期
徐志彬,陈胜楠,陈立欣,刘清泉,刘平生,张志强†
(1.北京林业大学水土保持学院,100083,北京; 2.内蒙古林业科学研究院,010010,呼和浩特)
早期研究认为,植物气孔在夜间会处于关闭状态,因此不存在夜间液流[1-3]。近些年来,越来越多研究表明大多数植物在夜间存在明显的树干液流[4]。受环境条件和自身生理特征影响,植物夜间液流一般占整日液流的5%~25%[5],在极端环境下,这一比例可达到60%[6]。目前,大部分生理生态和生态水文模型均假定,受太阳辐射限制,夜间植物气孔关闭,不存在水分流失[2,7]。显然,这种假设会导致低估植被耗水,从而影响客观评价森林植被变化对水文循环的影响。
夜间液流对植物生长发育有着重要生理意义。植物夜间液流主要用于夜间蒸腾和夜间补水[5]。这一水分运动现象在促进氧气[8]与养分[9]运输的同时,提高了植物黎明前茎干及叶片水势[5]。夜间气孔开放也有助于增加植物次日凌晨固碳量[10],促进植物生长,并在一定程度上减小干旱对植物光合作用及生长发育的影响[11]。此外,夜间液流有助于减少植物木质部栓塞和气穴形成,避免植物因干旱发生水力衰竭甚至死亡现象[11-12]。植物夜间液流受到多种环境因子共同作用。然而,不同生态环境下夜间液流与环境因子相关性仍存在差异[13-16]。王志超等[15]研究发现,在我国南方湿润地区尾叶桉人工林夜间液流速率与饱和水汽压差、夜间风速和日间蒸腾量呈极显著正相关(P<0.01),其认为夜间蒸腾和树干补水共同驱动尾叶桉夜间耗水。Han等[16]发现我国西北半干旱山区华北落叶松夜间液流量与环境因子相关性不显著(P>0.05),但与日间液流量呈显著正相关(P<0.05),因此,认为夜间液流主要用于缓解因日间蒸腾而产生的树体水分胁迫。
植物在不同生态环境下有不同的水分利用策略,生长在干旱及半干旱地区植被夜间失水量要高于其他地区[17-18],这导致干旱及半干旱地区城市树木可能更容易因缺水而面临干旱胁迫[19]。在此背景下,当发生季节性干旱时,植被夜间液流量可能会因环境因子变化产生差异,如不进行准确定量研究,将导致对植被耗水估算产生误差,影响当地水资源决策及管理。目前,有关树木夜间液流变化特征及影响因子相关研究主要集中于湿润区和干旱半干旱山区天然林及人工林,半干旱城区树木夜间液流化特征及其影响因子还有待进一步研究。鉴于此,笔者以我国北方城市绿化最为常见乡土树种油松(Pinustabulaeformis)为对象,采用Granier热扩散探针法[20]连续测定其液流变化,结合同步测定小气候和土壤含水量变化,旨在:1)揭示油松林分夜间液流动态变化特征;2)量化和比较油松林分夜间液流月际变化,讨论夜间液流对月耗水估算影响;3)分析影响油松林分夜间液流主要驱动因子。以期为揭示不同环境因子影响下油松林分夜间液流用途提供依据。
1 研究区概况
研究地点位于内蒙古自治区呼和浩特市赛罕区树木园(E 111°42′, N 40°49′),海拔1 056 m。该园始建于1954年,园区占地22 hm2,当地属于典型温带大陆性气候,年降水量300~450 mm,降雨多集中于7—9月。园内土壤以冲积土为主,pH值8左右,现有乔灌木树种500余种。在树木园内选取具有代表性的60年生油松样地,样地面积0.6 hm2,林分密度705株/hm2,平均树高(10.43±1.22)m,平均胸径(23.3±4.15)cm,平均冠幅(18.02±7.91)m2。
2 材料与方法
2.1 边材面积测定
考虑到林分内样木存在个体差异,将油松样地根据胸径分布,每3 cm划分为1个径阶,共5个径阶,分别为18~≤21、21~≤24、24~≤27、27~≤30和30~≤33 cm,在各径阶选取1~2棵生长状况良好、不偏心、不偏冠的树木作为待测样木(表1)。由于栽植时间较长,油松林内出现自然变异,即油松林内胸径和树高不存在显著相关(P>0.05)。为了避免损伤样木,在生长季初期选择样地内其他20 株样木利用生长锥进行取样,将所得组织染色处理后观察边材厚度,并计算边材面积。利用胸径和边材面积建立关系方程(式1),利用其计算样地内每棵树木的边材面积,按照径阶划分累加后得到样地边材总面积。

表1 样木基本特征Tab.1 Characteristics of sample trees
As=0.231 6DBH2.318 1(R2=0.97,n=20)。
(1)
式中:As为边材面积,cm2;DBH为树木胸径,cm。
2.2 树干液流测定
在2019年生长季(2019年5—9月),采用Granier热扩散探针(Dynamax,USA)对8株油松样木进行树干液流连续测定,使用10 mm TDP探针测量边材厚度<20 mm的样木,使用20 mm TDP探针测量边材厚度≥20 mm的样木。对于TDP探针长度低于边材厚度的样木,使用Mccarthy等[21]提出的裸子植物树干液流校正公式进行校正。在样木北向距离地面高1.3 m处刮去木质部以外的树皮,划定5 cm×5 cm的区域,用电钻打孔后将TDP探针插入。为避免探针机械损伤并减小降水及太阳辐射对测量结果的影响,在探针插入后使用玻璃胶进行密封,并在外部包裹铝箔纸。探针安装完成后,将其与CR1000数据采集器(campbell scientific)及加热器连接,并外接太阳能电池板,使用12 V电压为其持续供电,将程序设定为每30 min记录1次,定期下载数据并对样地及仪器进行维护。
使用Baseliner 3.0将采集的原始温差数据转换为液流数据,将太阳总辐射R<5 W/m2时段内的树干液流定义为夜间液流[8]。
2.3 环境因子测定
在样地10、30和50 cm处埋入5TE土壤水分传感器(Decagon,USA),连接EM50数据采集器(Decagon,USA)每隔30 min采集1次数据,使用10~50 cm土壤含水量平均值计算土壤相对含水量,根据土壤相对含水量判断土壤水分相对状况[22]。同时,在园区空旷位置安装小型自动气象站(Dynamet,USA),每隔30 min采集1次数据,主要监测指标有空气温度、空气湿度、风速、降雨量及太阳总辐射,并通过空气温度及空气湿度计算饱和水汽压差[23]。
2.4 数据处理及分析
使用SPSS 24.0和Excel 2013进行数据处理。因各月夜间液流的启动时间存在差异,本研究根据太阳总辐射R的变化规律将树干液流及环境因子数据划分为夜间(R<5 W/m2)和日间(R≥5 W/m2)2个时段。在日尺度上累加得到林分日间液流量、林分夜间液流量和太阳总辐射,取平均值计算夜间饱和水汽压差、夜间风速、夜间温度和土壤相对含水量。使用Origin 9.0绘制图像,图表中所用数据均为月平均值±标准差。采用配对样本t检验对日间及夜间环境因子,整日液流与日间液流量进行差异性分析。运用偏相关法分析夜间液流与环境因子的相关关系。使用逐步回归分析判断夜间液流贡献率的主要影响因子。
3 结果与分析
3.1 环境因子变化特征
2019年生长季期间,各月气象因子存在明显月变化(图1)。研究期间,饱和水汽压差日均值为(1.13±0.48)kPa,夜间饱和水汽压差在6月达到最大值,并呈减小趋势(图1b)。太阳总辐射日均值为(18.32±5.87)MJ/(m2·d),最大值为28.26 MJ/(m2·d),最小值为2.12 MJ/(m2·d)(图1a)。风速日均值为(0.34±0.13)m/s,日平均风速在5月达到最大值,日间与夜间风速存在显著性差异(P<0.05)。研究期间样地累计降雨313.95 mm,降雨多集中在7—9月,8月累计降雨量最大,达到103.63 mm(图1)。土壤相对含水量大小为8月>9月>7月>5月>6月,7月15日后随着降雨的增加,样地土壤水分条件得到改善(图1c)。
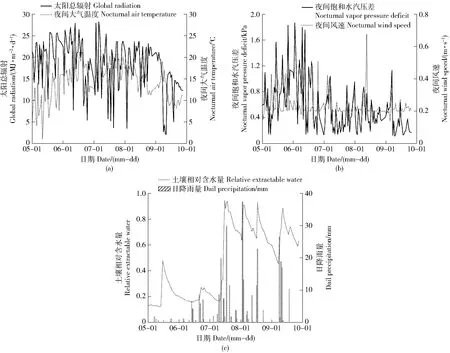
图1 各月环境因子变化Fig.1 Monthly variations of environment factors
3.2 夜间液流速率变化特征
通过观察图2可知,油松林分各月夜间液流速率具有较大差异,但总体趋势相对一致,均为单一递减趋势,但不同月份下降幅度不同。各月在00:00前均可以观测到明显夜间液流,且00:00前液流速率下降幅度更为明显。7、8和9月夜间液流在00:00后逐渐趋于稳定,5和6月夜间液流在00:00后仍有明显下降,但相比00:00前下降趋势较为缓慢(图2)。

图2 各月林分夜间液流速率变化Fig.2 Variations of stand nocturnal sap flow velocity in different months
3.3 夜间液流量与夜间液流贡献率月变化
研究期间,油松林分夜间液流量呈现出明显月变化(图3)。5月夜间液流量最大,可达到2.46 mm,其次依次为9月(1.48 mm)、6月(1.48 mm)、8月(1.37 mm)和7月(1.05 mm)。油松林分各月整日液流量与日间液流量呈极显著相关(P<0.01),且二者呈极显著差异(P<0.01)(表2)。研究期间油松林分夜间液流平均贡献率为7%(P<0.05),5月夜间液流占比要高于其他月份。在林分尺度计算油松生长季月液流量时,若不考虑夜间液流,月液流量将被低估3.0%~11.5%(图3)。

表2 各月林分整日液流量与日间液流量相关性及配对样本t检验Tab.2 Correlation and paired samples t-test betweenstand total daily sap flow and stand diurnal sapflow in different months

图3 各月林分夜间液流量占月累计液流量的比例Fig.3 Proportion of stand nocturnal sap flow to stand monthly total sap flow in different months
3.4 夜间液流的影响因子
在日尺度对2019年生长季油松林分夜间液流量与各环境因子进行偏相关分析。结果显示:油松林分夜间液流量与饱和水汽压差呈极显著正相关(P<0.01),饱和水汽压差是油松林分夜间液流量最主要影响因子,除9月外,饱和水汽压差可以解释各月79.2%、67.2%、71.6%和39.8%夜间液流量变化;7月风速与夜间液流量呈显著负相关(P<0.05),其他月份风速与夜间液流量相关性不显著(P>0.05);土壤相对含水量在6和7月与夜间液流量呈显著正相关(P<0.05),5、8和9月与夜间液流量相关性不显著(P>0.05)(表3);5和6月油松林分夜间液流量与日间液流量相关性不显著(P>0.05),7—9月二者呈显著正相关(P<0.05)(图4)。

图4 各月林分夜间液流量与日间液流量相关性Fig.4 Correlation between stand nocturnal sap flow and stand diurnal sap flow in different months

表3 各月林分夜间液流量与环境因子的偏相关系数Tab.3 Partial regression coefficients between stand nocturnal sap flow and environment factors in different months
利用2019年生长季液流及环境因子数据,对数据标准化后在日尺度上对夜间液流贡献率进行逐步回归分析,在优化剔除风速和土壤相对含水量后得到油松林分夜间液流贡献率与其影响因子的回归方程为
Cn=9.285-8.212Ed+3.475VPD-2.799R总
(R2=0.44,n=153)。
(2)
式中:Cn为油松林分夜间液流贡献率,%;Ed为油松林分日间液流量,mm;VPD为饱和水汽压,kPa;R总为日间太阳总辐射,MJ/(m2·d)。
由式(2)可知,生长季油松林分夜间液流贡献率与饱和水汽压差呈正相关,与日间液流量和日间太阳总辐射呈负相关。不同影响因子对油松林分夜间液流贡献率的作用由大到小依次为日间液流量>饱和水汽压差>日间太阳总辐射。
4 讨论
4.1 夜间液流变化特征
生长季油松林分各月夜间液流速率变化具有一致性,均为单一递减趋势,且00:00前液流速率变化幅度更为明显。00:00前后液流速率变化幅度差异可能与树体水分补充有关。Zeppel等[13]研究发现桉树在日落后5 h完成树体水分补充,张婕等[24]同样发现树木夜间液流速率前半夜较后半夜更为活跃。树木在日间蒸腾旺盛导致蒸腾失水,树干水分严重亏缺,在日落后树体与土壤间会存在较大水势差[25]。因此,夜间液流初始阶段会存在较高的夜间液流来弥补树体水分亏缺[13],因日落后饱和水汽压差、风速及等环境因子会明显低于日间,树木夜间蒸腾量也会明显低于日间,当树体自身水分亏缺得到缓解后,夜间液流量便会随之减小[24]。
本研究发现生长季油松林分夜间液流量变化特征为5月>9月>6月>8月>7月。5和6月油松林分土壤水分条件较差,当处于干旱胁迫的状态下,植物会通过增加夜间液流量来缓解自身水分亏缺,减少植物木质部栓塞和气穴形成,避免植物因干旱发生水力衰竭甚至死亡现象[11-12]。7和8月降雨量远高于5和6月,油松林分土壤水分条件较好,树木在阴雨天也可以补充树体水分,缓解自身水分亏缺,提高树体水势[24],这导致7和8月夜间液流量相对于5和6月有所下降。9月树木生理活动减弱,但油松林分日间液流量仍相对较高,由于阴雨天气的减少,树木需要通过夜间液流弥补因日间蒸腾所导致的水分亏缺,因此,9月夜间液流量相比7和8月有所上升。
植物夜间液流贡献率与自身水分利用策略有关,现有研究发现植物夜间液流占整日液流5%~25%[5]。生长季油松林分夜间液流平均贡献率为7%,各月油松林分夜间液流贡献率存在显著差异(P<0.05)。通过式(2)可知,日间液流量是油松林分夜间液流贡献率最重要的影响因子,5、6月处于旱季,降雨匮乏,土壤含水量不断降低会导致叶片气孔对饱和水汽压差和太阳辐射的敏感性增加,为了减小树体水分流失和木质部空化风险,植物往往会通过关闭气孔来减少日间蒸腾[19]。7月后降雨量增加,土壤水分条件相对较好,树木冠层气孔导度和日间蒸腾量可以维持较高水平,从而导致夜间液流贡献率相对较低。虽然不同月份夜间液流贡献率存在差异,但各月油松林分整日液流量与日间液流量呈极显著差异(P<0.01),在林分尺度计算月液流量时,若不考虑夜间液流,月液流量将被低估3.0%~11.5%
4.2 环境因子对夜间液流量的影响
在日尺度上,生长季油松林分夜间液流量与饱和水汽压差、风速和土壤相对含水量呈显著相关(P<0.05)。除9月外,饱和水汽压与各月夜间液流量均呈显著正相关(P<0.05)。饱和水汽压差和空气温度升高会导致叶片边界层与气孔内腔的水汽压梯度增加,叶片气孔导度上升,水分气化过程变快,促进夜间蒸腾,提高夜间液流量[11,14]。由于夜间平均风速小于日间,夜间风速增加会导致叶片与大气间水汽压差减小,降低空气和叶片温度,导致树木液流活动减弱,树体水分利用效率降低。油松林分夜间液流量与土壤相对含水量在6和7月呈显著正相关(P<0.05),夜间液流量随土壤相对含水量的降低而降低,这与Fisher等[14]的研究结果一致,土壤含水量降低会使得植物组织水力导度和叶片气孔导度降低,叶片水分穿透能力减弱,导致植物液流速率下降[5]。
本研究发现土壤相对含水量降低会导致夜间液流量减少,但在土壤水分条件相对较差的5月,由于高饱和水汽压差及其他因子影响下,油松林分夜间液流量明显高于其他月份,这可能与各环境因子的协同作用有关。夜间液流受多种因素共同影响,这些因素之间也存在着复杂的作用关系,因此不能用单一因子来表征夜间液流驱动力,在未来对夜间液流机制研究中应更加注重不同环境下各因子对夜间液流综合影响。
4.3 夜间液流的用途
植物蒸腾活动与环境因子关系较为密切,在以往研究中多通过分析环境因子与夜间液流相关关系来判断是否存在夜间蒸腾,当夜间液流与环境因子相关性较强时认为夜间液流被用于夜间蒸腾,当与环境因子相关性较差且与日间液流量相关性较强时则认为被用于夜间补水[8]。本研究中,除9月外,生长季油松林分各月夜间液流量与饱和水汽压差呈显著正相关(P<0.05),7—9月油松林分夜间液流量与日间液流量呈显著正相关(P<0.05),这说明半干旱区油松林分存在夜间蒸腾和补水的可能。由于植物夜间蒸腾和补水作用是同时发生,目前为止仍没有有效的方法可以将夜间蒸腾和夜间补水进行定量区分。有学者在树体底部和顶部各安装一个液流探针,将两者差值作为夜间补水量[26],但此种方法没有考虑枝条和叶片储水,夜间液流以整日5%~15%的水分运输速率实现50%~100%叶片膨胀,叶片在夜间吸收的水分大部分通过叶片膨胀进行储存,小部分通过蒸腾损失,叶片储存的水分为次日植物生长提供了条件[10],若忽略枝条和叶片储水,将低估夜间补水量。此外,夜间气孔开放会导致冠层与土壤间水势梯度增加,驱动树体内水分流动,为木质部和冠层韧皮部提供水分补充[18]。因此,不能仅通过夜间液流与环境因子和日间液流量的相关性判断夜间液流的用途。由于实验仪器限制,本研究中没有测量其他生理指标。在未来研究中,可将土壤及叶片水势、树体水容、叶片气孔导度、树木径向生长变化量进行监测,结合树木生理指标,进一步了解油松在不同时期夜间液流调控机制。
在发生季节性干旱时,林分蒸腾量远高于降雨量(图1和图3),树木虽然可以通过降低叶片气孔导度[19]或增加夜间液流量等途径维持自身水分平衡,但气孔关闭会导致植物光合作用降低[27],阻碍植物生长。因此,在半干旱区林业规划中,除考虑美观外,应在结合当地环境条件及树木蒸腾耗水特征,选择气孔调节作用较强的树种进行合理密植,并在季节性干旱发生时适度灌溉,以提高树体水分利用效率,满足树木生长发育需要。
5 结论
1)生长季油松林分夜间液流呈现出明显的日、月变化。在日尺度上,油松林分夜间液流速率呈单一递减趋势,各月在00:00点前均可以观测到明显夜间液流,且00:00点前液流速率下降幅度更为明显;在月尺度上,油松林分夜间液流量5月最大,其次依次为9月>6月>8月>7月。
2)在日尺度上,生长季油松林分夜间液流量与饱和水汽压差、土壤相对含水量和日间液流量呈显著正相关(P<0.05),与风速呈显著负相关(P<0.05)。生长季油松林分夜间液流平均贡献率为7%,不同影响因子对夜间液流贡献率的作用大小依次为日间液流量>饱和水汽压差>日间太阳总辐射。
猜你喜欢
石油石化绿色低碳(2019年6期)2019-01-14
世界家苑(2018年9期)2018-09-18
现代农村科技(2018年3期)2018-01-18
山东林业科技(2017年1期)2017-06-29
电源技术(2015年5期)2015-08-22
电源技术(2015年7期)2015-08-22
应用数学与计算数学学报(2014年1期)2014-09-26
电测与仪表(2014年18期)2014-04-04
汽车零部件(2014年2期)2014-03-11
中国质量与标准导报(2014年5期)2014-02-28
