
发酵构树饲料对蛋鸡产蛋后期生产性能、蛋品质、血清生化指标和肠道组织形态的影响
2021-11-05 04:00陶振阳张金金赵凌平武晓红王占彬
动物营养学报 2021年10期
陶振阳 张金金 张 邦 赵凌平 武晓红 王 军 王占彬*
(1.河南科技大学动物科技学院,洛阳 471003;2.禾佳农业科技有限公司,宜阳 471600)
随着我国畜禽养殖业的快速发展,对饲料原料的需求量不断增加,传统玉米-豆粕型饲粮对饲料原料特别是蛋白质原料的需求量极大。陶莎等[1]报道,2019年我国蛋白质原料年需求量超过8 000万t,其中豆粕占85%以上;2017年以来受中美贸易摩擦影响,豆粕市场价格呈先扬后抑的走势,极大增加养殖成本和饲料企业经营风险。2020年9月15日国务院《关于促进畜牧业高质量发展的意见》中指出,要健全饲草供应体系,因地制宜推行粮改饲,开发利用杂交构树(Broussonetiapapyrifera)、饲料桑等新饲草资源,调整优化饲料配方结构,促进玉米豆粕减量替代。因此,木本饲料资源的科学开发利用对促进畜牧业可持续发展意义重大。
构树具有热量高、速生、适应性强、易繁殖、轮伐期短的特点,广泛分布于我国温带、热带,在平原、丘陵或山地都能生长[2]。杨祖达等[3]报道认为,构树叶粗蛋白质(CP)含量达24%,通过小鼠急性毒性试验证明,急性毒性半数致死剂量(LD50)>12 g/kg,属实际无毒级,是优良的蛋白质饲料。杂交构树叶CP含量为26.1%,粗脂肪(EE)含量为5.2%,仅从营养成分看可替代一部分豆粕[4]。杂交构树作为一种新型的粗饲料,其不同部位的化学成分不同[5]。构树叶营养成分含量高且更易消化,由于其碳水化合物水溶性较低,不易直接被动物消化吸收,所以通过对杂交构树进行发酵处理来提高构树饲料的品质是必要的,建议在与饲料混合前机械分离叶与茎,以提高构树的利用效率。张兴等[6]在湘沙猪配套系商品猪饲粮中添加20%、30%全株发酵构树饲料,发现对其生长性能无显著影响,且能改善肌肉风味和品质。宋博等[7]在三元杂交育肥猪低蛋白质饲粮中分别添加10%全株构树干粉和全株发酵构树饲料,发现对其生长性能无负面影响;添加10%全株发酵构树饲料使育肥猪背最长肌中游离氨基酸和肌内脂肪含量显著增加,有效降低脂肪率和平均背膘厚,改善肌肉风味。宋柏螈等[8]在奶牛全混合日粮中添加8%和12%杂交发酵构树饲料,发现能降低牛乳体细胞含量,并提高乳清抗氧化能力。梁春宇等[9]在泌乳前期荷斯坦经产奶牛饲粮中添加7%杂交青贮构树,发现对奶牛的采食量和产奶量均无显著影响。Si等[10]报道认为,饲粮中添加10%~15%杂交青贮构树饲料能降低泌乳奶牛牛奶中体细胞数,增加牛奶中多不饱和脂肪酸含量,改善牛乳品质,并提高奶牛免疫功能和抗氧化功能。林萌萌等[11]报道认为,饲粮中用青贮杂交构树替代部分蛋白质饲料及青贮饲料对肉羊的采食量、日增重和料重比均无显著影响,但适宜替代量还有待研究。黄炎坤等[12]用青贮构树枝叶替代25%稻糠饲喂221日龄皖西白鹅,发现其可显著加深蛋黄颜色,且对鹅的生产性能和繁殖性能无显著影响。李艳芝等[13]报道认为,饲粮中添加1.5%~2.0%构树叶可提高45周龄海兰褐蛋鸡的生产性能和蛋品质。研究认为,将构树叶自然干燥,粉碎后过筛,制备发酵原料(80%构树叶粉+20%玉米粉),分别喷入2%枯草芽孢杆菌GZ3和1%枯草芽孢杆菌HS3菌液密封发酵8 d后,在35日龄爱拔益加(AA)肉仔鸡饲粮中分别添加3.75%、5.00%和6.75%发酵构树叶进行饲喂,结果表明,构树叶经发酵后粗纤维(CF)含量不同程度降低,CP含量显著增加,未对AA肉仔鸡的生长性能造成不良影响,且可降低皮脂厚、腹脂率和肌间脂肪宽,有效减少肌肉滴水损失和烹饪损失,改善鸡肉风味[14-15]。吴建平等[16]在饲粮中添加2%、4%和6%构树叶粉,发现对35日龄雌性良凤花肉鸡的CP、粗灰分(Ash)、钙(Ca)和磷(P)的表观利用率及生产性能、血清生化指标和胴体性状均无显著影响,表明构树叶作为非常规蛋白质饲料原料应用在良凤花肉鸡饲粮中是可行的。
目前有关研究主要集中在将构树叶或构树叶粉直接饲喂蛋鸡,用发酵全株杂交构树枝叶饲料饲喂产蛋后期蛋鸡的研究未见报道。本试验旨在探究发酵构树饲料对产蛋后期蛋鸡的饲喂效果及选用发酵构树饲料替代部分玉米、豆粕的可行性,为推进发酵构树饲料在蛋鸡生产中的推广应用提供科学参考依据。
1 材料与方法
1.1 试验材料
试验用发酵构树饲料由洛阳市宜阳县某公司提供。选用杂交构树101,在株高90~120 cm时刈割、切断、粉碎,添加发酵益生菌(乳酸菌)和酶制剂(纤维素酶、甘露聚糖酶、木聚糖酶、β葡聚糖酶)充分混合后,装袋密封发酵20 d,制成发酵构树饲料。全株构树发酵前后营养成分见表1,发酵构树饲料氨基酸组成及含量见表2。
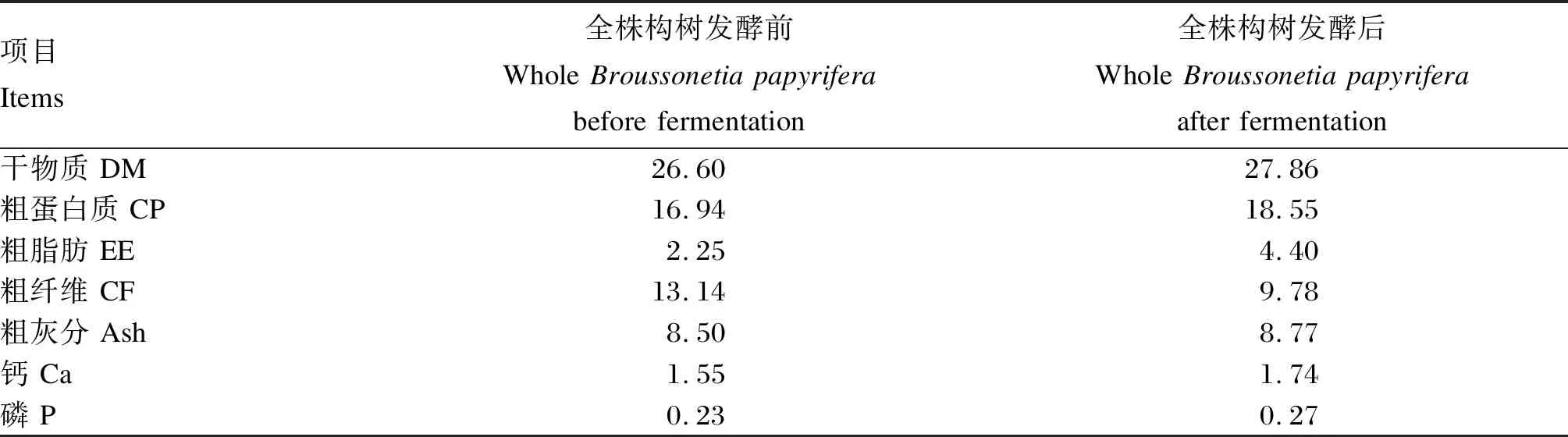
表1 全株构树发酵前后营养成分比较(风干基础)

表2 发酵构树饲料氨基酸组成及含量
1.2 试验设计
试验于洛阳市新安县红雨养殖场进行,选择67周龄、精神状态良好、产蛋率相近的海兰褐蛋鸡360只,随机分为4组,每组6个重复,每个重复15只鸡。对照组饲喂基础饲粮,Ⅰ、Ⅱ、Ⅲ组分别饲喂用1.5%、3.0%和4.5%发酵构树饲料替代基础饲粮中部分玉米和豆粕的试验饲粮。预试期7 d,正试期56 d。基础饲粮参照NRC(2004)、《鸡饲养标准》(NY/T 33—2004)及《2015版海兰褐商品代蛋鸡饲养管理手册》配制,为玉米-豆粕型粉状饲料。试验饲粮组成及营养水平见表3。
1.3 饲养管理
鸡舍为半开放式,试验蛋鸡3层全阶梯式笼养,每笼3只鸡。舍内采用自然通风搭配纵向负压通风,自然光照结合人工补光恒定光照16 h/d。试验全期每天07:30、14:30及19:30各喂料1次,蛋鸡自由采食、饮水,每天08:00清粪1次,笼舍消毒和蛋鸡常规免疫均按照鸡场常规饲养管理程序进行。
1.4 测定指标及方法
1.4.1 生产性能
试验期间,每天17:00收集鸡蛋,详细记录各组蛋鸡每重复每天产蛋数、破软蛋数、蛋重和鸡只死淘数,计算死淘率。每周统计各组蛋鸡每重复采食量,统计平均日采食量、平均蛋重、平均日产蛋量、产蛋率及料蛋比。
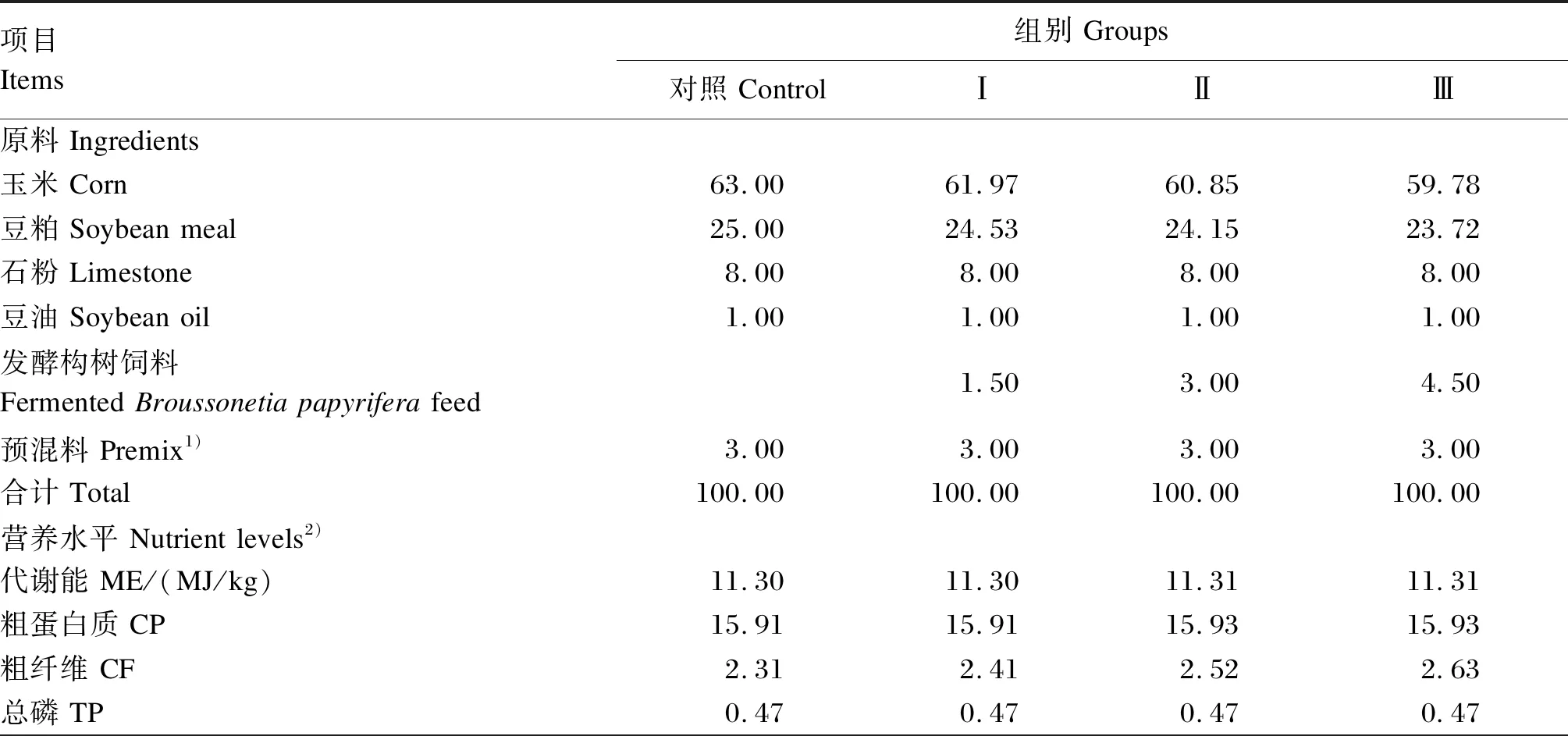
表3 试验饲粮组成及营养水平(风干基础)

续表3项目Items组别 Groups对照 Control ⅠⅡⅢ钙 Ca3.603.623.643.66赖氨酸 Lys0.810.810.810.81蛋氨酸+半胱氨酸 Met+Cys0.560.560.560.56
1.4.2 蛋品质
于试验第28天和第56天,每个重复采集6枚鸡蛋用于蛋品质测定。蛋重、蛋壳颜色、哈夫单位、蛋白高度、蛋黄颜色使用蛋品质分析仪(ORKA EA-01,以色列)测定;鸡蛋纵径长、横径长用电子游标卡尺测定并计算蛋形指数(纵径长/横径长);使用强度测定仪(NFN388,日本)测定蛋壳强度;用镊子除去内壳膜后,使用螺旋测微器分别测量蛋壳锐端、钝端及中间部分的厚度计算蛋壳厚度;使用电子分析天平(Mettler-Toledo MS-TS,瑞士)测量蛋黄重和蛋壳重并计算蛋黄相对重(100×蛋黄重/蛋重)和蛋壳相对重(100×蛋壳重/蛋重);蛋黄总胆固醇(TC)含量采用试剂盒(购于南京建成生物工程研究所)使用酶标仪测定。
1.4.3 血清生化指标
于试验第55天,每个重复随机抽取1只蛋鸡,空腹12 h后,真空采血管翅下静脉采血3.5 mL,常温静置1 h后于离心机4 000 r/min离心10 min,分离血清,分装后于-20 ℃冰箱保存待测。采用动物专用全自动生化分析仪(IMagic-V7,库贝尔)测定血清总蛋白(TP)、白蛋白(ALB)、球蛋白(GLO)、甘油三酯(TG)、尿酸(UA)、总胆固醇(T-CHO)、高密度脂蛋白胆固醇(HDL-C)、低密度脂蛋白胆固醇(LDL-C)、葡萄糖(GLU)含量及碱性磷酸酶(ALP)、丙氨酸氨基转移酶(ALT)、天门冬氨酸氨基转移酶(AST)活性。
1.4.4 肠道组织形态
于试验结束当天,每个重复随机选取1只蛋鸡,禁食12 h,颈动脉放血致死,解剖后取十二指肠、空肠和回肠中段各4 cm左右肠道样品,生理盐水缓慢冲洗肠道内容物后,4%多聚甲醛溶液固定24 h。将样品用浓度梯度逐级递增的乙醇溶液脱水,二甲苯冲洗后进行浸腊包埋,并用石蜡切片机修整后修整至4 μm厚度制成切片,进行苏木精-伊红(HE)染色,中性树胶封片,显微镜镜检。使用图像分析软件(Case Viewer),测量并记录绒毛高度(villous height,VH)、隐窝深度(crypt depth,CD),计算绒毛高度/隐窝深度(VH/CD)。
1.5 数据统计与分析
使用Excel 2003初步整理试验数据,然后使用统计软件SPSS 20.0对试验数据进行单因素方差分析(one-way ANOVA),用Duncan氏法对分析结果进行多重比较,P<0.05表示差异显著,数据分析结果用“平均值±标准差”表示。
2 结 果
2.1 发酵构树饲料对蛋鸡产蛋后期生产性能的影响
由表4可知,与对照组相比,Ⅰ、Ⅱ和Ⅲ组蛋鸡的产蛋率、平均蛋重、平均日采食量和料蛋比均无显著差异(P>0.05);与对照组相比,Ⅲ组的平均日产蛋量降低1.8 g,差异显著(P<0.05),Ⅰ组降低0.13 g(P>0.05),Ⅱ组增长0.68 g(P>0.05);各试验组的死淘率与对照组相比显著降低(P<0.05)。
2.2 发酵构树饲料对蛋鸡产蛋后期蛋品质的影响
由表5可知,与对照组相比,试验各组蛋鸡的蛋形指数、蛋壳颜色、蛋壳强度、蛋白高度、哈夫单位、蛋黄颜色、蛋壳厚度、蛋黄相对重、蛋壳相对重及蛋黄总胆固醇含量均无显著差异(P>0.05)。

表4 发酵构树饲料对蛋鸡产蛋后期生产性能的影响

表5 发酵构树饲料对蛋鸡产蛋后期蛋品质的影响
2.3 发酵构树饲料对蛋鸡产蛋后期血清生化指标的影响
由表6可知,Ⅱ组蛋鸡的血清TC含量显著低于对照组、Ⅰ和Ⅲ组(P<0.05);Ⅰ、Ⅱ、Ⅲ组的血清TP、ALB、GLO、TG、UA含量较对照组有升高趋势,但未达到显著水平(P>0.05);Ⅱ、Ⅲ组的血清GLU含量和AST活性与对照组相比有所降低,但差异不显著(P>0.05)。各组间蛋鸡的血清AST、ALT、ALP活性和HDL-C和LDL-C含量均无显著差异(P>0.05)。
2.4 发酵构树饲料对蛋鸡产蛋后期肠道组织形态的影响
由表7可知,与对照组相比,Ⅰ、Ⅱ、Ⅲ组蛋鸡的十二指肠、空肠和回肠绒毛高度、隐窝深度及绒毛高度/隐窝深度均无显著差异(P>0.05)。
3 讨 论
3.1 发酵构树饲料对蛋鸡产蛋后期生产性能的影响
张兴等[6]分别在湘沙猪配套系育肥猪前期(50 d)和后期(53 d)饲粮中添加20%、30%发酵构树饲料,发现对猪的生长性能无显著影响。宋博等[7]在三元杂交育肥公猪低蛋白质饲粮中添加10%全株发酵构树饲料,发现对猪的生长性能没有负面影响。李海新[17]研究发现,在生长猪饲粮中添加10%和15%发酵构树叶时,平均日增重、平均日采食量和料重比呈下降趋势,当添加量为20%时对生长猪的育肥性能产生负面影响。邓铭等[18]报道认为,在断奶仔猪饲粮中用发酵构树饲料替代9%的基础饲粮时,平均日增重和平均日采食量有下降趋势。但林萌萌[19]等报道认为,用全株发酵构树替代育肥猪蛋白质饲料(豆粕)时,替代量不宜超过3%,替代量越高,育肥性能越差。上述研究结果的差异可能与试验猪所处时期不同和发酵构树饲料的发酵工艺(如发酵菌种、浓度,发酵时间)不同有关,具体原因尚需进一步研究。目前有关发酵构树饲料在蛋鸡饲粮中的应用研究还鲜有报道。本试验中,随着发酵构树饲料替代比例的增加,蛋鸡的产蛋率、平均日采食量、料蛋比和平均蛋重未见不利影响,但发酵构树饲料替代基础饲粮中玉米和豆粕量达4.5%时产蛋量显著降低。杂交构树经过生物发酵后有效钝化底物中抗营养因子,降解部分蛋白质、脂肪及多糖(纤维)等大分子物质,积累有益代谢产物,可溶性多肽、短链脂肪酸和有机酸等,形成营养丰富、益生菌含量高且适口性好的饲料[20]。在配制试验饲粮时发现,将发酵构树饲料与基础饲粮粉碎混合后呈淡绿色,气味酸甜清新,试验蛋鸡采食积极性高,发酵构树饲料的适口性好且对蛋鸡有一定的诱食作用。随着发酵构树饲料替代比例的增加,饲粮CF水平提高导致胃中产生过量丙酸,使蛋鸡易饱腹感增强,平均日采食量下降,产蛋率有降低的趋势,试验组的死淘率显著降低,导致平均日产蛋量降低。
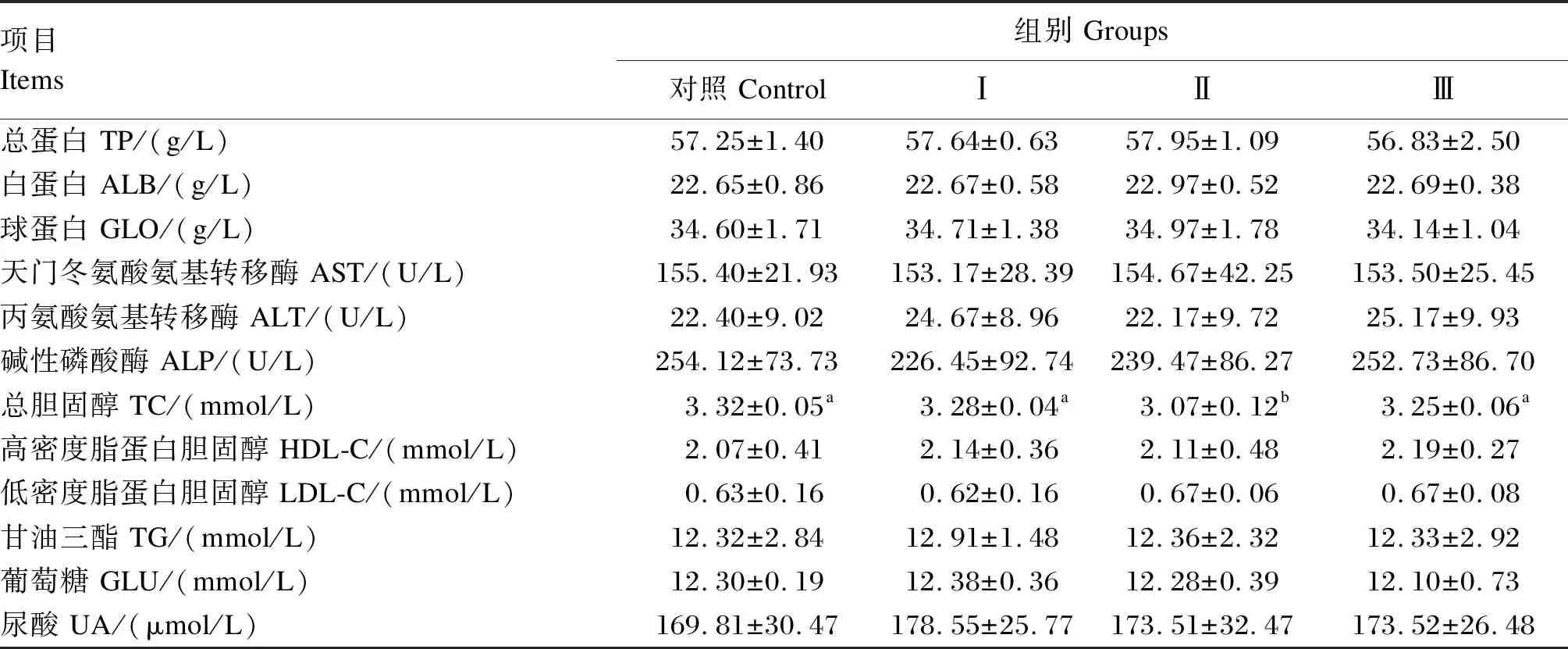
表6 发酵构树饲料对蛋鸡产蛋后期血清生化指标的影响

表7 发酵构树饲料对蛋鸡产蛋后期肠道组织形态的影响
3.2 发酵构树饲料对蛋鸡产蛋后期蛋品质的影响
蛋品质主要通过蛋形指数、蛋壳强度、蛋壳厚度、哈夫单位和蛋黄颜色等指标综合反映[21-24]。蛋形指数是描述鸡蛋形状的参数,对食用营养价值无影响,但关系到孵化率和破蛋率,影响种用价值[25-27]。标准禽蛋形状为椭圆形,蛋形指数介于1.30~1.35,本试验中,试验组与对照组的蛋形指数均在上述区间,且无显著差异,属合格商品蛋。蛋鸡品种不同蛋壳颜色不同;同种蛋鸡饲喂不同饲料产出的鸡蛋蛋黄颜色多种多样,由浅黄色到金黄色再到深橙色不等,蛋黄颜色取决于蛋鸡从饲粮中摄取和沉积类胡萝卜素的数量,蛋鸡自身不能合成类胡萝卜素,只能从饲粮中摄取[28-29]。黄竹等[30]报道,在蛋鸡基础饲粮中添加不同水平发酵饲料对蛋黄颜色有不同程度的提高。李艳芝等[13]研究表明,在45周龄海兰褐蛋鸡饲粮中添加1%~2%构树叶使蛋黄颜色显著提高。本试验研究表明,随着发酵构树饲料在产蛋后期蛋鸡饲粮中替代量的增加,蛋黄颜色有提高的趋势,与上述研究结果一致,提示发酵构树饲料替代基础饲粮中部分玉米和豆粕对蛋鸡产蛋后期蛋品质无影响。
3.3 发酵构树饲料对蛋鸡产蛋后期血清生化指标的影响
TP包括ALB和GLO,具有维持机体血管中正常的胶体渗透压和酸碱度的作用,对运输各种代谢产物及调节运输物质起着重要的生理作用[31],与机体免疫功能密切相关。彭箫等[25]在54周龄京红蛋鸡饲粮中添加30 g/kg发酵饲料,发现未对血清TP、ALB和GLO含量产生显著影响,本试验结果与上述试验结果一致,这表明在蛋鸡产蛋后期饲粮中添加发酵构树饲料对蛋鸡的蛋白质分解代谢无影响。
血清中的酶具有极高的特异性,当机体出现应激或发生病变时其活性会发生一些变化,临床上通过血清酶学检测来判断器官的功能状态和机体代谢水平。AST、ALT和ALP活性能反映肝脏代谢蛋白质和氨基酸的能力,以及肝脏的健康状况。本试验中,各组间蛋鸡的血清AST、ALT和ALP活性无显著差异,表明发酵构树饲料对蛋鸡的肝脏功能无不利影响。
GLU是供应机体生命活动的能量来源,大部分储存在肝脏和肌肉中,少部分随血液运送到组织和器官进行氧化和分解,为生命活动提供能量,正常情况下,由胰岛素等激素参与,使糖的合成与分解代谢处于动态平衡,血糖保持相对稳定[32]。司丙文等[33]研究表明,在杜寒杂交肉羊饲粮中添加青贮杂交构树对血清GLU、UN、TP及TG含量无显著影响。本试验结果也表明,饲粮中用发酵构树饲料替代基础饲粮中部分玉米和豆粕对蛋鸡的血清GLU、TP和TG含量没有产生显著影响,与前人研究结果一致。
血清中TC、HDL-C和LDL-C是反映机体脂质代谢水平的重要指标。彭箫等[25]在京红蛋鸡产蛋后期饲粮中添加30 g/kg的发酵饲料,发现显著降低蛋鸡的血清和肝脏TC含量。邓铭等[18]用6%发酵构树饲料替代35日龄断奶仔猪基础饲粮30 d后,仔猪的血清TC含量显著降低。本试验结果表明,以3%发酵构树饲料替代67周龄海兰褐蛋鸡饲粮中玉米和豆粕会显著降低血清TC含量,说明3%替代比例能促进蛋鸡脂质代谢能力。HDL-C被称为“好胆固醇”,它通过促进胆固醇逆向转运、抗氧化和抗炎等机制来实现抗动脉粥样硬化;反之,LDL-C会促进脂肪在血液中沉积形成动脉粥样硬化,导致血管壁损伤,对机体有负面影响,因此又被称为“坏胆固醇”[34]。本试验中,随着发酵构树饲料替代量的增加,HDL-C含量有上升趋势,说明一定比例的发酵构树饲料对蛋鸡脂质代谢有促进作用。
UA是由蛋白质代谢终产物——氨转化而来,血清中UA含量减少可反映机体对蛋白质沉积好,氮利用率高[35]。本试验中,各组间血清UA含量无显著差异,这说明一定比例的发酵构树饲料不影响蛋鸡对氮的利用率。
3.4 发酵构树饲料对蛋鸡产蛋后期肠道组织形态的影响
小肠是动物吸收营养物质和水分的重要场所,同时也是最长的消化器官,食物进入小肠内会刺激肠道分泌肠液,经化学和机械性消化后转化为营养物质被小肠黏膜吸收,同时也可阻碍有害物质的入侵,是机体维持内环境稳态的重要屏障[36]。小肠内壁有许多褶皱,褶皱上的凸起为小肠绒毛,可以增加养分与水分吸收的表面积,而肠道绒毛高度增加进一步增加小肠内壁与食糜接触面积,使机体更高效的吸收利用营养物质[37]。小肠绒毛延伸向下的底部就是肠道隐窝,隐窝底部为干细胞,制造出短暂分裂细胞(TAC),能在短时间增生并向上移动,TAC在移动过程会分化为杯状细胞与肠细胞,吸收营养物质并更新肠细胞,隐窝深度降低表明肠道上皮细胞分化率的增加,促进肠绒毛生长[38]。本试验结果表明,在蛋鸡产蛋后期用发酵构树饲料替代基础饲粮中部分玉米和豆粕对蛋鸡的肠道组织形态无不良影响。
4 结 论
不同比例发酵构树饲料替代蛋鸡产蛋后期饲粮中部分玉米和豆粕对蛋品质和肠道组织形态无显著影响;4.5%替代比例显著降低产蛋量,3.0%替代比例显著降低血清TC含量,从生产角度分析,3.0%替代比例效果最佳。
猜你喜欢
动物营养学报(2022年8期)2022-09-01
中国饲料(2021年17期)2021-11-02
21世纪(2019年12期)2019-12-18
绿色中国(2019年16期)2019-11-26
中国林业产业(2019年3期)2019-03-25
小哥白尼(野生动物)(2018年9期)2018-12-08
中国扶贫(2017年16期)2017-08-16
湖南农业(2017年1期)2017-03-20
西南学林(2016年2期)2016-11-08
动物营养学报(2015年9期)2016-01-07
