
Hippo signaling: bridging the gap between cancer and neurodegenerative disorders
2021-11-02 02:57NehaGogiaAnuradhaVenkatakrishnanChimataPrajaktaDeshpandeAditiSinghAmitSingh
中国神经再生研究(英文版) 2021年4期
Neha Gogia , Anuradha Venkatakrishnan Chimata , Prajakta Deshpande ,Aditi Singh, Amit Singh
Abstract During development, regulation of organ size requires a balance between cell proliferation,growth and cell death. Dysregulation of these fundamental processes can cause a variety of diseases. Excessive cell proliferation results in cancer whereas excessive cell death results in neurodegenerative disorders. Many signaling pathways known-to-date have a role in growth regulation. Among them, evolutionarily conserved Hippo signaling pathway is unique as it controls both cell proliferation and cell death by a variety of mechanisms during organ sculpture and development. Neurodegeneration, a complex process of progressive death of neuronal population, results in fatal disorders with no available cure to date. During normal development, cell death is required for sculpting of an organ. However, aberrant cell death in neuronal cell population can result in neurodegenerative disorders. Hippo pathway has gathered major attention for its role in growth regulation and cancer, however, other functions like its role in neurodegeneration are also emerging rapidly. This review highlights the role of Hippo signaling in cell death and neurodegenerative diseases and provide the information on the chemical inhibitors employed to block Hippo pathway. Understanding Hippo mediated cell death mechanisms will aid in development of reliable and effective therapeutic strategies in future.
Key Words: animal models; cell death mechanisms; cell-signaling; chemical inhibitors;Drosophila eye; Hippo pathway; neurodegeneration; neurological diseases; therapeutic targets
Introduction
A fundamental area in biology is to study growth regulation and organ size. Animal organ size complies with a complex set of biological constraints that ensures survival. During organogenesis, patterning and growth are tightly regulated to determine the correct size. Growth regulation determines the number of cells in organ(s) of a multi-cellular organism by balancing the cell biological processes like cell polarity,cell proliferation and cell death (Dong et al., 2007; Penzo-Mendez and Stanger, 2015; Eder et al., 2017). Broadly speaking, growth properties of a developing field can be regulated by cell number alterations or by changes in cell size. Several highly conserved growth regulatory pathways are involved in growth regulation and organ size homeostasis such as insulin signaling pathway, target of rapamycin (TOR),and Hippo signaling pathway. Insulin signaling is involved in maintaining carbohydrate metabolism while interacting with other conserved signaling pathways like TOR, AMP-activated protein kinase (AMPK) and phosphatidylinositol 3-kinase (PI3K)pathways (Nijhout, 2003; Badisco et al., 2013). TOR signaling controls organ size growth by increasing the cell size; whereas Hippo signaling pathway determines organ size by regulating the cell number. To obtain an ideal organ size, optimum levels of Hippo signaling is required during organogenesis(Camargo et al., 2007; Dong et al., 2007; Tumaneng et al., 2012). Downregulation of Hippo signaling causes over proliferation, which results in cancer, whereas upregulation of Hippo signaling triggers cell death and apoptosis resulting in incomplete or reduced organs or cellular dysfunctions. This growth regulatory Hippo pathway is highly conserved from fruit flies to humans and has been known to play a crucial role in growth, cell proliferation and apoptosis (Kango-Singh and Singh, 2009; Zhao et al., 2011; Verghese et al., 2012b;Halder and Camargo, 2013; Han, 2019; Moya and Halder,2019). The purpose of this review is to highlight the role of signaling pathway in two distinct disease like cancer and neurodegenerative disorders.
Discovery of Hippo Pathway
The Hippo pathway, which was initially discovered in theDrosophila melanogaster(a.k.afruit fly) model, and has been a subject of extensive investigation both inDrosophilaas well as mammals in the last two decades.Drosophila, with its arsenal of powerful genetic manipulation tools, allows for cost-effective and large scale screens equipped to answer scientific questions immediately.Drosophilawith a shorter life cycle, strong reproductive ability, and genetic legacy of more than a century of this model system, has proved to be a highly versatile and tractable model for genetic screens. The conservation of genetic machinery from fruit flies to human makesDrosophilaan ideal human disease model (Bier, 2005;Singh and Irvine, 2012; Yeates et al., 2019). Therefore, it not only serves as a model for genetic research but also for a wide array of studies including investigations of basic cellular and molecular mechanisms of human disease. To identify genes involved in growth regulation, an EMS mutagenesis screen was conducted inDrosophilaeye usingeyeless-FLP/FRTsystem that resulted in identification of members of Hippo signaling pathway. The rationale was to identify the genetic mosaic clones, which exhibit phenotypes affecting cell proliferation,cell death and overall organ size in the adult fly eye.
The adult compound eye ofDrosophiladevelops from an epithelial bilayer structure housed inside the larva, which has blue print for the adult eye and the head structures,and is referred to as the larval eye-imaginal disc. During late second instar and early third instar of larval development,a synchronous wave of differentiation emerges from the posterior margin and moves anteriorly, which results in differentiation of retinal precursor cells to the retinal neurons.This wave of differentiation is referred to as Morphogenetic furrow (MF) (Ready et al., 1976). During larval development to pupal metamorphosis, eye imaginal disc differentiates into pupal retina and later into the adult eye comprising of 600–800 unit eyes called ommatidia (Ready et al., 1976; Kango-Singh et al., 2003, 2005; Singh and Choi, 2003; Tare et al.,2013a, b; Gogia et al., 2020a, b).Drosophilaeye model due to its accessibility and dispensability (for viability), combined with its complex architecture and development, availability of fate markers, have made it one of the most intensively studied organs in the fly to model human disease.
The first growth regulation component of the Hippo signaling pathway was reported from the fly model and later it was confirmed that the Hippo pathway is evolutionarily conserved from flies to humans. Hippo pathway, a kinase cascade, also known as Salvador/Warts/Hippo (SWH) pathway, gets its name, “Hippo” from the characteristic overgrowth phenotype of adult structures shown in the pathway component mutants(Justice et al., 1995; Xu et al., 1995; Harvey et al., 2003; Jia et al., 2003; Pantalacci et al., 2003; Udan et al., 2003; Wu et al.,2003; Kango-Singh and Singh, 2009). In 2002, two drosophila lab identified a novel mutation calledsalvador(sav, also referred to asShar-pei) that encodes a WW45 domain protein which exhibits over proliferation phenotypes in theDrosophilaeye (Kango-Singh et al., 2002; Tapon et al., 2002; Huang et al., 2005; Kango-Singh and Singh, 2009; Zhao et al., 2011;Verghese et al., 2012a). Later, mutants for other components likehippo(hpo, a mammalian Ste20-like kinases in mammals),warts(wts, a serine/threonine kinase) that exhibit similar bigger eyes due to increased number of inter-ommatidial cells in the pupal retinae, were identified (Udan et al., 2003; Wu et al., 2003). The kinases Hippo (MST1/2 in mammals), Warts(Wts or LATS1/2 in mammals) and transcriptional co-activator Yki (YAP/TAZ in mammals), form the core components of Hippo signaling pathway (Huang et al., 2005; Figure 1).
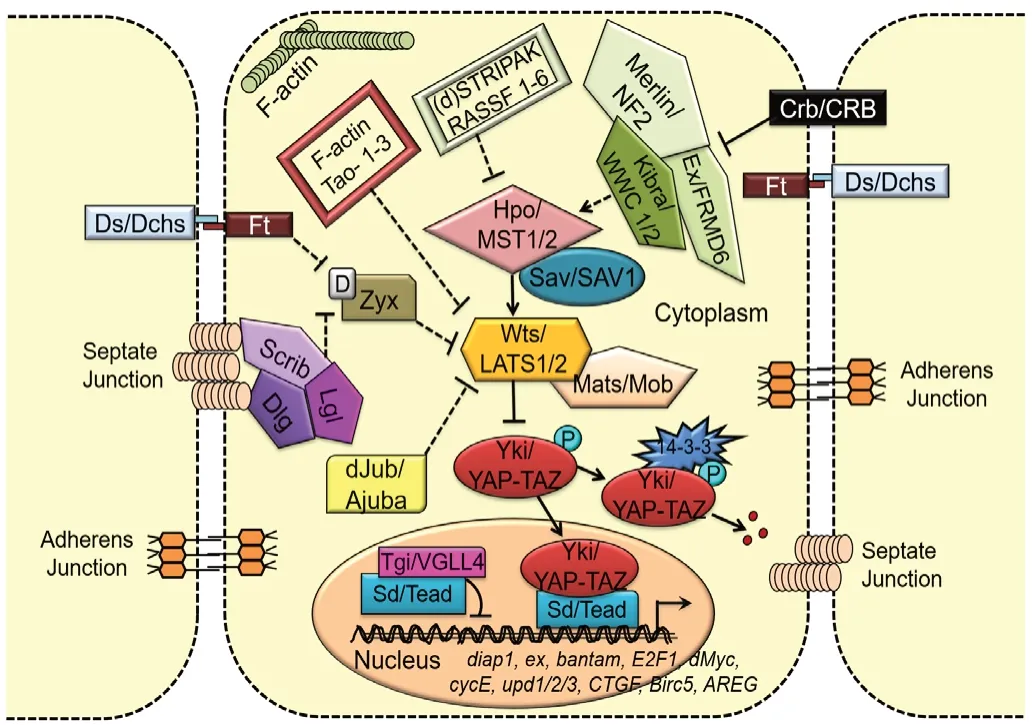
Figure 1 |Evolutionarily conserved Hippo signaling pathway from flies to mammals.
There are several upstream components, which regulate Hippo pathway by influencing the cell polarity, cell junctions,and stress induced responses (Meng et al., 2016). Activation of Hpo, upon phosphorylation by its upstream regulators,results in phosphorylation of Sav, which leads to formation of Hpo-Sav complex. This complex activates its downstream components Warts and Mats (Mob as a tumor suppressor,mammalian homolog Mob) via phosphorylation and forms Wts-Mats complex. The Wts-Mats complex further phosphorylates the transcriptional co-activator Yorkie (Yki or YAP/TAZ in mammals), which binds to the 14-3-3 adaptor proteins, gets restricted to the cytoplasm, and gets degraded(Huang et al., 2005; Zhao et al., 2007; Oh and Irvine, 2008;Kango-Singh and Singh, 2009; Snigdha et al., 2019). However,when Hippo is inactivated, Yki is not phosphorylated;therefore, Yki translocates into the nucleus and binds to a transcription factor Scalloped (Sd, mammalian homolog TEAdomain-containing (TEAD) and induces transcription of its downstream targets. The various downstream targets of Yki are Cyclin E (Cyc-E) required for cell proliferation and cell cycle progression, Myc required for growth, Drosophila associated inhibitor of apoptosis 1 (Diap1), Bantam microRNA important for cell survival (Jia et al., 2003; Wu et al., 2003, 2008; Huang et al., 2005; Nolo et al., 2006; Thompson and Cohen, 2006;Peng et al., 2009; Neto-Silva et al., 2010; Ziosi et al., 2010),Wingless (Wg),Drosophilahomolog of Wnt (in mammals), and ligand for evolutionarily conserved Wg/Wnt signaling pathway(Singh et al., 2002; Wittkorn et al., 2015) and Homothorax(Hth), a MEIS class of transcription factor, act as the negative regulators of eye formation in theDrosophilaeye (Pai et al.,1998; Singh et al., 2011). Downregulation of Hippo pathway causes uncontrolled growth due to increased level of Yki homolog YAP/TAZ results in human cancers and tumors (Harvey and Tapon, 2007) whereas activation of Hippo signaling triggers cell death (apoptosis) (Udan et al., 2003; Verghese et al., 2012a). As such, Hippo pathway has multiple functions,and coordinates a balance between growth, cell survival and death, which is required for organ formation with balanced size and shape (Kango-Singh and Singh, 2009; Han, 2019), and has implications in cancers, cell death, or neurodegeneration.
Multiple Roles of Hippo Signaling Pathway
Apart from its role in growth regulation and organ size control(Xu et al., 1995; Jia et al., 2003; Pantalacci et al., 2003; Udan et al., 2003; Wu et al., 2003; Kango-Singh and Singh, 2009; Han,2019), Hippo pathway and its downstream targets have other functions in cell proliferation, apoptosis, cell differentiation,cell competition, cell contact inhibition, epithelialmesenchymal transition (EMT), innate immunity, regeneration and repair (Mehta and Singh, 2019; Moya and Halder, 2019;Figure 2). Downstream targets of Hippo pathway are crucial for various cellular processes like (a) cell proliferation, cell cycle progression (Cyclin E), (b) inhibition of cell death (Diap1),(c) growth (Myc), and (d) cell survival (microRNA Bantam) (Jia et al., 2003; Wu et al., 2003, 2008; Huang et al., 2005; Nolo et al., 2006; Thompson and Cohen, 2006; Peng et al., 2009;Neto-Silva et al., 2010; Ziosi et al., 2010; Halder and Camargo,2013; Snigdha et al., 2019).
The pathway regulates cell growth and cell survival as evidenced from YAP and TAZ, which promotes tissue growth by regulating TEADs (Goulev et al., 2008; Wu et al., 2008). Hyper activation ofykiinDrosophilaor YAP and TAZ in mammals causes ectopic cell proliferation (Pan, 2010). Disruption of YAP-TEAD interaction leads to promotion and progression of cancer. Dysregulation of the Hippo pathway has been shown to cause loss of contact inhibition, ectopic cell proliferation,accelerated cell cycle advancement, and increased metastasis(Pan, 2010; Nishio et al., 2016). Mutations in Hippo pathway components also confer cancer cells a competitive advantage by activatingYkitarget gene,myc(Neto-Silva et al., 2010;Ziosi et al., 2010). The Hippo signaling pathway also maintains immune homeostasis and regulates immunological functions(Hong et al., 2018; Figure 2). MST1/2 has been found to cross-talk with other important pathways by regulating phosphorylation and activation of critical signaling players such as PKC, FOXO, or AKT of these pathways to regulate both innate and adaptive immunity (Hong et al., 2018; Yamauchi and Moroishi, 2019). Hippo signaling is known to be involved in stem cell and progenitor cell maintenance, renewal, and expansion in a variety of tissues to play a crucial role in regeneration and repair (Wang et al., 2017; Moya and Halder,2019). The pathway has implications in cancers, cell death and neurodegeneration. In the nervous system, Hippo signaling is involved in the proliferation and differentiation of neuronal progenitors (Lin et al., 2012; Wittkorn et al., 2015), migration(Hindley et al., 2016), myelination (Deng et al., 2017),dendritic arborization (Emoto, 2012) and neurodegeneration.
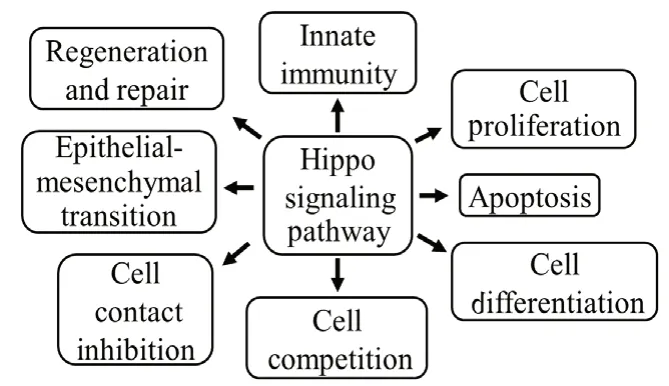
Figure 2 |Schematic representation of multiple functions of Hippo signaling pathway.
Role of Hippo in Cell Death and Neurodegenerative Diseases
Role of Hippo in cell deathNeurodegeneration is characterized by excessive cell death of functional neurons (Singh, 2012). Aberrant activation of Hippo signaling causes cell death, makes it a potential candidate that can link cancer with neurodegeneration. Severalin vitroandin vivomodels includingDrosophila melanogaster(fruit fly),Mus musculus(mouse),Danio rerio(zebrafish), are being used to study the role of Hippo pathway in cell death and neurodegenerative diseases. Hippo signaling has been used to study growth, cell death and to identify potential targets which can block or ameliorate the onset of neurodegenerative diseases (Table 1) (Udan et al., 2003; Wu et al., 2003;Lehtinen et al., 2006; Kango-Singh and Singh, 2009; Lee et al.,2013; Sanphui and Biswas, 2013; Wei et al., 2013; Sarkar et al., 2016, 2018; Mueller et al., 2018; Deshpande et al., 2019;Gogia et al., 2020b; Irwin et al., 2020). These models aid in understanding molecular pathogenesis, mechanism of action,progression and therapeutics for neurodegenerative diseases due to conservation of basic genetic machinery.
Hippo pathway causes cell death by a variety of mechanisms that include -
(a) Inactivation of Yki. Yeast two hybrid screening techniques were employed to identify the downstream targets or partners of Wts, which resulted in identification of Yki/YAP (Yorkshire)as the binding partner of Wts, and as the link between Hpo signaling activation and its transcriptional control. Genetic epistasis experiments revealed antagonistic interaction betweenykiand previously identified components of Hippo pathway such ashpo,savandwts. Furthermore, Hippo pathway negatively regulates Yki activity. LOF of yki resulted in downregulation of Diap1 levels and showed severe growth defects (Huang et al., 2005; Han, 2019).
(b) Upregulation ofwg/WNT, the downstream target of Hippo pathway. Wg, a ligand of Wingless/ Wnt signaling,regulates the expression of Homothorax (Hth), and it acts as a negative regulator of retinal fate. Hyperactivation of Yki by misexpression of yki3SAin the developing fly eye causes ectopic induction of Wg. This ectopic upregulation of Wg (Singh et al.,2002, 2006) causes induction of Hth, suppresses progression of Morphogenetic Furrow (MF) and retinal differentiation (Pai et al., 1998; Singh et al., 2002, 2006, 2011, 2019; Wittkorn et al., 2015) (Figure 3).
(c) Activation of caspases. InDrospohilamodel,dronc(Drosophilacaspase-9 homologue) acts as a transcriptional target of Hippo pathway and regulates cell proliferation and cell death during development (Verghese et al., 2012a).Hyperactivation of Hippo pathway in the fly eye causes induction ofDrosophilaeffector caspases Drice and Dronc,which result in increased cell death (Verghese et al., 2012a).The molecular mechanism underlying Hpo-mediated Dronc regulation is not fully understood.
(d) Activation of evolutionarily conserved c-Jun-amino terminal (NH2)-Kinase (JNK) signaling. Hippo signaling has been shown to play role(s) in neurodegeneration (Azuma et al., 2018; Gogia et al., 2020b; Irwin et al., 2020). Fly model has allowed researchers to exploit genetic tools like GAL4/UAS system to misexpress target gene(s) of interest along the spatial-temporal axes in specific tissues of interest (Brand and Perrimon, 1993). Genetic screens conducted in flies have shown Hippo as a genetic modifier of neurodegeneration as seen in amyloid-beta 42 (Aβ42) mediated neurodegeneration observed in Alzheimer’s disease (AD) and FUS accumulation mediated neurodegeneration as reported in amyotrophic lateral sclerosis (ALS) (Gogia et al., 2020b; Irwin et al., 2020)(Figure 3). Hippo pathway causes cell death by activation of JNK signaling. Interestingly, in AD model, it has been seen that JNK signaling and Hippo signaling are involved in positive feedback loop to trigger neurodegeneration (Irwin et al.,2020).
(e) Oxidative stress mediated activation of MST1-FOXO signaling. Oxidative stress regulates cell survival and homeostasis by unknown mechanisms. Activation of MST1(by oxidative stress) induces cell death in primary mammalian neurons. If MST1 phosphorylates FOXO transcription factors,which disrupts its interaction with 14-3-3 protein and thereby facilitates translocation of FOXO transcription factor into the nucleus resulting in transcription of pro-apoptotic genes that trigger neuronal cell death (Lehtinen et al., 2006; Figure 3).
Role of Hippo in neurodegenerative diseases
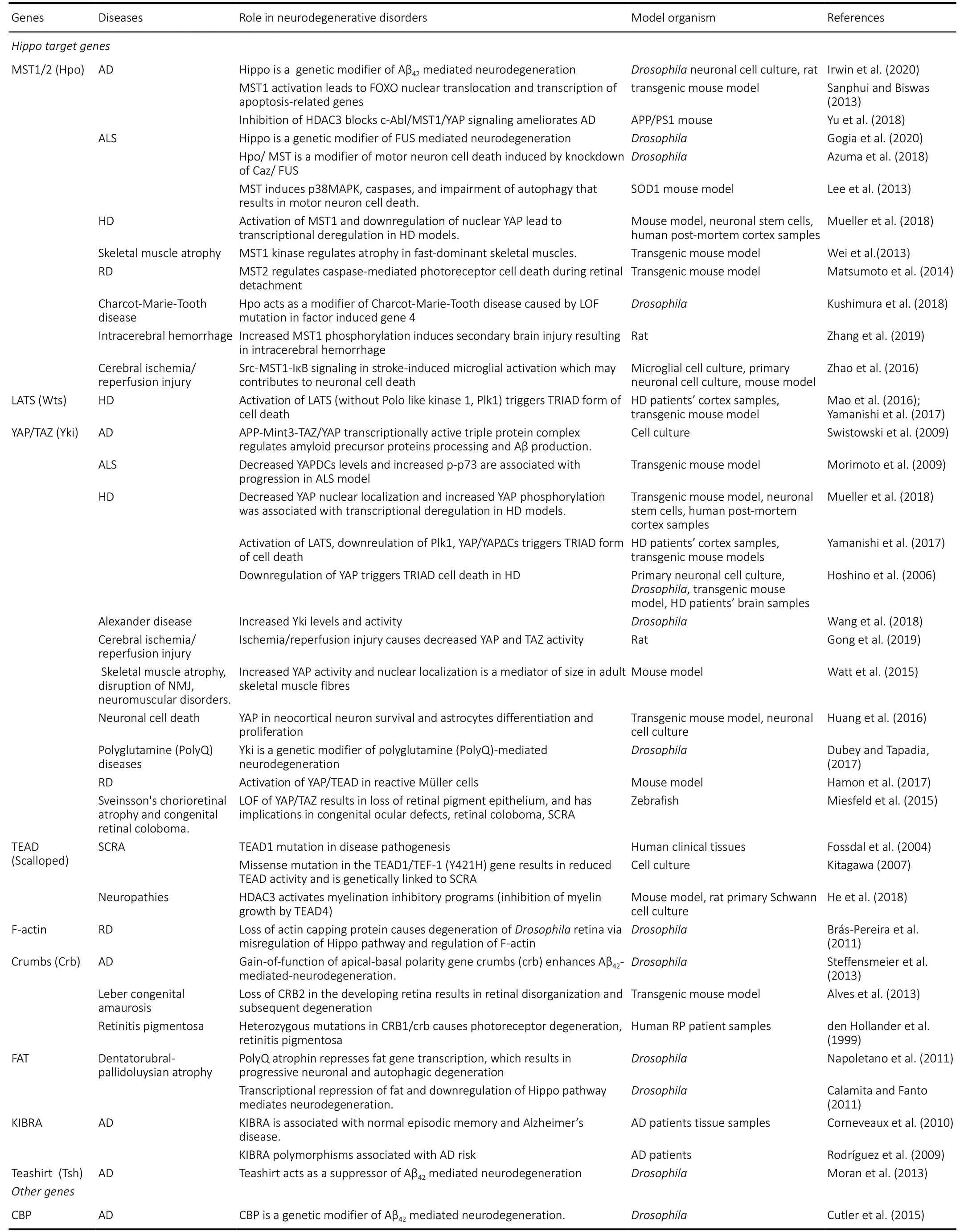
Table 1 |Hippo pathway genes and their roles in neurodegenerative diseases
· AD is an age related progressive neurodegenerative disorder with no cure to date (Tare et al., 2011; Sarkar et al., 2016,2018). It is caused by many factors such as (i) accumulation of extracellular Aβ42 plaques, (ii) aggregation of intracellular neurofibrillary tau tangles, (iii) oxidative stress due to mitochondrial dysfunction, and (iv) genetic factors (Crews and Masliah, 2010; O’Brien and Wong, 2011; Selkoe and Hardy, 2016; Yeates et al., 2019). AD model in flies allows misexpression of human-Aβ42in the retinal neurons and helps in understanding the role of Hippo in cell death and neurodegeneration (Figure 4) (Tare et al., 2011; Moran et al.,2013; Steffensmeier et al., 2013; Cutler et al., 2015; Sarkar et al., 2016, 2018; Irwin et al., 2020). Other AD models in fly model allows ectopic of human Aβ42expression in other neuronal population(s) of the nervous system. Studies conducted using one of those fly models have shown that accumulation of Aβ42activates the Hippo pathway that further activates JNK pathway by a positive feedback loop mechanism to induce cell death (Figure 4; Irwin et al., 2020). These fly models identified other genetic modifiers like homeotic geneteashirt(tsh) as a suppressor (Moran et al., 2013), andcrumbs(crb) with a neuroprotective function in Aβ42mediated neurodegeneration (Steffensmeier et al., 2013). Prostate derived sterile 20 like kinases (PSKs/TAOKs), an upstream regulator of Hippo signaling cascade, has been reported in the pathogenesis of AD. Activation of PSKs, phosphorylates Tau,results in Alzheimer’s pathology as observed inin vitrocell culture models (Tavares et al., 2013; Table 1). In AD, Hippo mediated neurodegeneration is triggered by dysregulation of MST-FOXO3a pathway, which in turn activates downstream apoptosis regulator Bcl-2 (Figure 5) (Sanphui and Biswas,2013; Fallahi et al., 2016). Hippo pathway is modulated by inhibition of Histone deacetylases (HDAC-3), which are abundantly expressed in the brain, and has neurotoxic effects. Inhibition of HDAC-3 Tyrosine-protein kinase ABL(c-Abl), p-MST, p-YAP, reduces oxidative stress and cell death in transgenic mice model of AD (APPswe/PS1dE9) (Figure 5 and Table 1; Yu et al., 2018). Some other Hippo pathway components that play a role in AD are listed in Table 1.
· ALS,a progressive neurodegenerative disorder, which involves death of upper and lower motor neurons in the brain and spinal cord. More than 30 genes including human-Fused in Sarcoma, FUS (Cabeza inDrosophila) and Cu/Zn superoxide dismutase (SOD1) are known to be associated with ALS pathogenesis (Rosen et al., 1993; Rutherford et al.,2008; Kwiatkowski et al., 2009). Misexpression of human FUS or mutant FUS in the fly eye causes degeneration of photoreceptor neurons and recapitulates neuropathological features similar to ALS (Lanson et al., 2011; Gogia et al.,2020b) (Figure 4). Hippo has been identified as a genetic modifier of FUS mediated neurodegeneration (Azuma et al.,2018; Gogia et al., 2020b). In fly model of ALS, accumulation of FUS leads to the activation of Hippo, which induces JNK pathway and triggers neuronal cell death (Gogia et al.,2020b) (Figures 3 and 4). Similarly, SOD1 (G93A) mouse model of ALS also identified MST1 (Hpo) as a key modulator of neurodegeneration and shows that modulation of levels of MST1 (Hpo), p38, MAPK and caspase-9, 3 and other autophagy markers, affects neuronal death (Figure 5) (Lee et al., 2013). Thus, ectopic Hippo pathway induction triggers neurodegeneration in ALS.
· Polyglutamine diseasesare caused by expansion of Cytosine-Adenine-Guanine (CAG) repeats encoding a polyQ(polyglutamine) tract in the respective proteins. This leads to aggregation of insoluble ubiquitinated proteins in different neuronal cell populations of the brain and spinal cord which triggers neuronal death (Paulson and Fischbeck, 1996;McGurk et al., 2015). Hippo pathway and its targets have been extensively studied to decipher the molecular pathogenesis in polyglutamine diseases. Studies conducted in Dentatorubral pallidoluysian atrophy (DRPLA) fly model has shown that presence of extra CAG repeats inatrophin-1(at-1) gene suppresses the transcription of tumor suppressor genefat(ft), an upstream regulator of Hippo pathway, which results in progressive neurodegeneration by regulating autophagy(Calamita and Fanto, 2011; Napoletano et al., 2011). A mouse model reported MST1 as a key player in enhancing fast skeletal muscle atrophy. It reports that increased levels of MST1 in fast skeletal muscles, increases phosphorylation of FOXO3a (a major mediator of muscle atrophy), which leads to its translocation into the nucleus resulting in skeletal muscle atrophy (Figure 5; Wei et al., 2013). Additionally, Yki/ YAP was identified as a genetic modifier of polyglutamine mediated neurodegeneration (Figure 5; Dubey and Tapadia, 2018). It is required to maintain both basal skeletal myofiber mass and size of adult skeletal muscles fibers through its interaction with TEAD. Increased TEAD activity during injury or degeneration of motor nerves mitigates the neurogenic muscle atrophy (Wattet al., 2015). As seen in denervation induced model of skeletal muscle atrophy, increased YAP nuclear localization regulates muscle mass and growth (Watt et al., 2015).
· Huntington’s disease (HD),an autosomal dominant neurodegenerative disorder, is caused by CAG repeat(s)expansion. Accumulation of mutant Htt protein triggers cell death. Studies conducted in mouse model of HD (CAG knock-in HdhQ111/Q111) indicated the role of Hippo pathway in HD (Mueller et al., 2018). In HD, the mutant Htt interacts with YAP and disrupts formation of TEAD/YAP complex,which triggers necrotic cell death known as transcriptional repression induced atypical cell death of neuron (TRIAD)(Mao et al., 2016; Yamanishi et al., 2017). TRIAD is mainly regulated by YAP and Hippo pathway in HD models (Hoshino et al., 2006; Yamanishi et al., 2017). Increased levels of MST1 and decreased levels of nuclear YAP are associated with TRIAD as seen in post-mortem HD cortex and HD mouse brain(Mueller et al., 2018). Activation of LATS1/Wts, suppression of cell cycle regulator Plk1 (mediates balance between TEAD/YAP-dependent necrosis and p73/YAP-dependent apoptosis),and reduced expression of YAP/ YAP∆C isoform in human HD brains, supports occurrence of TRIAD (Yamanishi et al., 2017).Thus, targeting TEAD/YAP transcription based necrosis activity of Hippo pathway can be effectively used for development of therapeutic targets of HD (Mao et al., 2016).
· Parkinson’s disease (PD),a progressive neurodegenerative disease, which exhibits symptoms of memory loss and movement related disorder. PD is characterized by formation of intracytoplasmic Lewy body (LB) inclusion structures,accumulation of α-synuclein in Lewy bodies and loss of dopaminergic neurons of substantia nigra, which result in neuronal death (Feany and Bender, 2000). Mutations in genes including Synuclein alpha (SNCA), leucine-rich repeat kinase 2(LRRK2), PTEN Induced Kinase 1 (PINK1) and others cause PD(Forno, 1996; Hardy et al., 2006). PINK1, is a mitochondrial serine/threonine kinase, which plays a crucial role in viral infection. Upon viral infection, PINK1 inhibits the formation of YAP1/IRF3 complex, and positively regulates retinoic-acidinducible gene-I (RIG-I) which triggers innate antiviral immune response (Figure 5; Zhou et al., 2019).
Role of Hippo in Other Neurodegenerative Diseases
· Retinal degeneration (RD)is caused by deterioration of the retina due to progressive death of photoreceptor cells(Wert et al., 2014). Several Hippo pathway members have known to be involved in retinal degeneration as listed in Table 1. Sodium hyaluronate induced retinal detachment model in MST knock out mice, shows MST-2 as key regulator of caspase-dependent death of photoreceptor cells resulting in loss of vision (Matsumoto et al., 2014). Studies conducted in RD mouse model have shown that Mueller’s glia gets highly activated upon loss of photoreceptor cells and that increases YAP and TEAD1 mRNA and protein levels (Hamon et al., 2017)(Table 1). Mutations in apical-basal polarity geneCrb(Crb2KO mouse using Cre/loxP technology) exhibits Leber congenital amaurosis (LCA) and retinitis pigmentosa (RP) disease, which exhibit characteristic progressive degeneration phenotype during late retinal development (den Hollander et al., 1999;Alves et al., 2013). In RD, the Hippo pathway and actin Capping protein inhibits the accumulation of F-actin. InDrosophilamodel, deregulation of Hippo pathway is associated with the loss of Capping protein and retinal degeneration of the adult fly (Brás-Pereira et al., 2011) (Table 1).
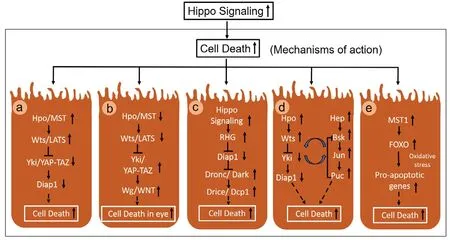
Figure 3 |Gain-of-function of Hippo signaling triggers cell death by various mechanisms.
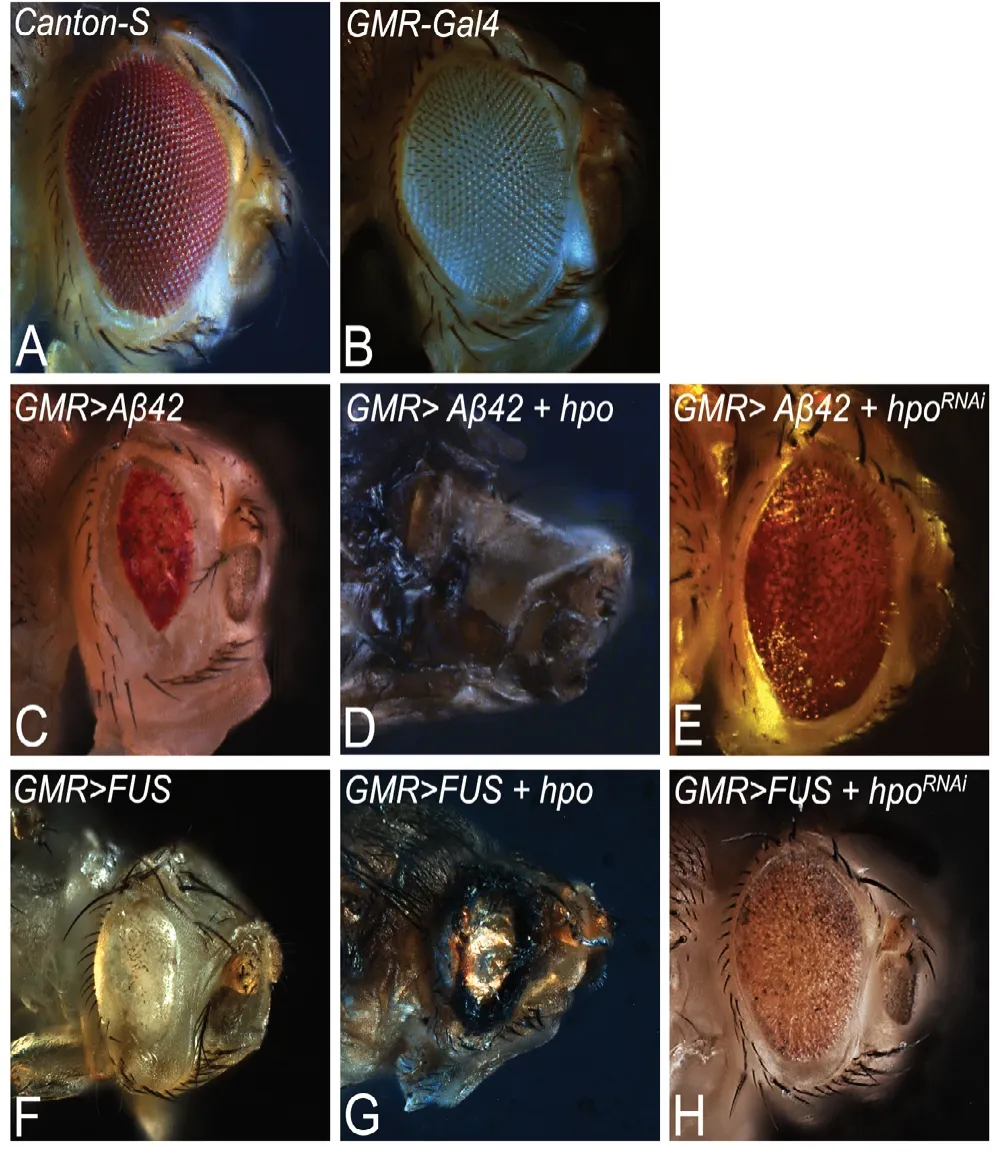
Figure 4 |Drosophila eye model of neurodegenerative disease.
· Sveinsson’s chorioretinal atrophy (SCRA)is an eye disease marked by bilateral chorioretinal degeneration. The disease is caused by missense mutation in gene coding for TEAD1.Missense mutation in human TEAD1 reduces its ability to bind to YAP/TAZ and results in SCRA pathogenesis as seen in mice model (Kitagawa, 2007). A previous study conducted in human cell lines show that mutation inTEAD1gene affects binding site of a retinal co-factor of TEAD1 and causes SCRA(Figure 5; Fossdal et al., 2004). YAP/TAZ-TEAD activity has also been found to be important for progenitor cells to form retinal pigment epithelium and has significant bearing in congenital ocular defects in SCRA and congenital retinal coloboma in Zebrafish Model (Miesfeld et al., 2015).
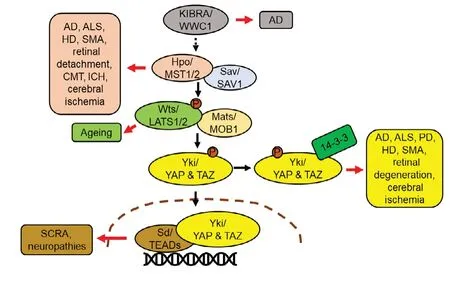
Figure 5 |Hippo pathway members’ aberrant expression leads to neurodegenerative disorders.
Hippo Pathway Members as Putative Therapeutic Targets
Neurodegenerative disorders are heterogeneous group of disorders characterized by reduced size of brain, which results in loss of cognitive and/or motor function(s). These neurodegenerative disorders are associated with lots of morbidity and puts an enormous burden on the society.Thus, there is a desperate need for development of reliable therapeutic strategies. One of the challenges is the lack of validated therapeutic target(s). Cellular therapies offer great promise in this aspect, however, they face obstacles like dealing with neurons that are post-mitotic in nature, and do not possess regenerative potential. An alternative solution for neurodegeneration is to screen for agents that block or prevent neuronal cell death. Blocking cell death machinery is known to have adverse effects, and therefore there is a need for other possible targets. Hippo pathway has been observed to be hyperactivated in several of these neurodegenerative disorders. This raises an important question of whether Hippo pathway or its targets can be utilized as the potential therapeutic targets to control progression of neurodegenerative disorders. Therefore, chemical inhibitors designed to modulate Hippo pathway or its target(s) that can block or delay neurodegeneration via inhibiting Hippo pathway directly or indirectly may serve as effective and reliable therapeutics in future (Schmidt-Erfurth and Hasan,2000; Oku et al., 2015; Qu et al., 2018). Several chemical inhibitors have been identified to modulate Hippo signaling levels in cancer. However, these drugs could also potentially be repurposed to treat neurodegenerative disease as loss-offunction of Hippo signaling causes over proliferation, which result in cancers while gain-of-function of Hippo signaling results in cell death and neurodegeneration (Figure 6) as observed in AD, ALS and other neurodegenerative diseases.Thus these chemical inhibitors of Hippo signaling can be a valuable resource for downregulating Hippo signaling, which can block neurodegeneration.
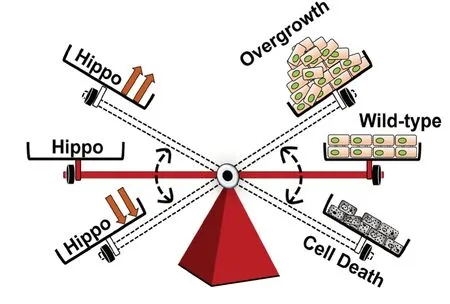
Figure 6 |Role of Hippo signaling during growth and neurodegeneration.
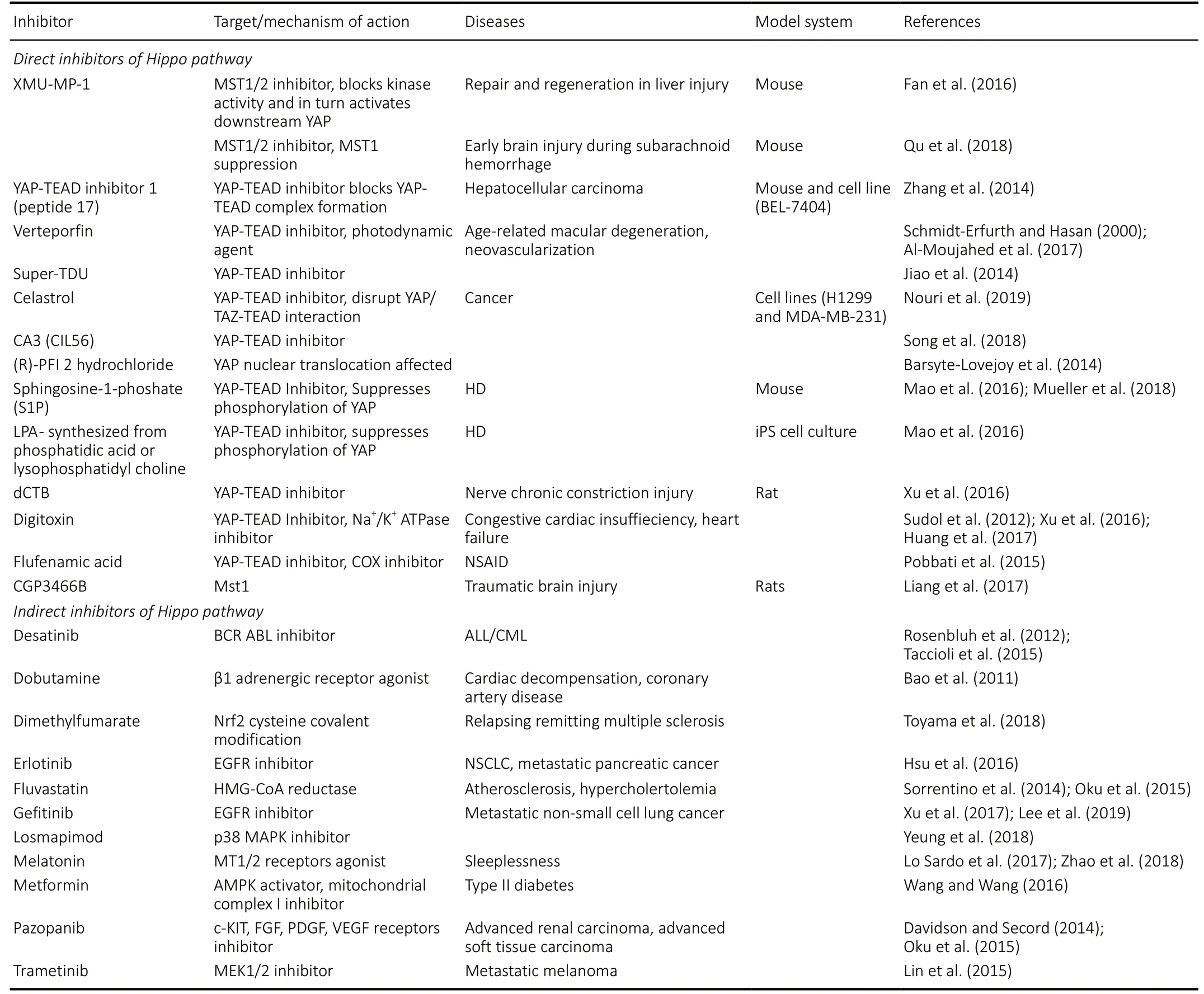
Table 2 |Chemical inhibitors of Hippo target genes may serve as putative therapeutic targets
Hippo pathway can be inhibited by using these chemical inhibitors (Schmidt-Erfurth and Hasan, 2000; Oku et al., 2015;Qu et al., 2018). The chemical inhibitors that target various components of Hippo pathway and are effective in treating growth related or neurodegenerative diseases with their mechanism of action are summarized in Table 2. Currently,there are three possible approaches for directly inhibiting Hippo pathway,viz., 1. Inhibition of Hpo/MST1/2 kinase(Fan et al., 2016; Qu et al., 2018), 2. Alteration of Yki/YAP localization (Barsyte-Lovejoy et al., 2014), and 3. Inhibition of Yki-Sd/YAP-TEAD interaction (Schmidt-Erfurth and Hasan,2000; Jiao et al., 2014; Zhang et al., 2014; Nouri et al., 2019).Alternatively, several small molecule inhibitors have been reported which target(s) other pathway components that interact with Hippo pathway (Oku et al., 2015) for example kinase inhibitors, which target Src family kinase, EGFR, MEK which interact with Hippo signaling (Lin et al., 2015; Oku et al., 2015; Hsu et al., 2016; Wang et al., 2016; Xu et al., 2017;Yeung et al., 2018; Lee et al., 2019) (Table 2). Other examples include inhibitors against Melatonin signaling, Wnt pathway,G-protein coupled receptors, mevalonate pathway which can also inhibit Hippo pathway (Bao et al., 2011; Rosenbluh et al., 2012; Davidson and Secord, 2014; Sorrentino et al.,2014; Oku et al., 2015; Taccioli et al., 2015; Lo Sardo et al.,2017; Toyama et al., 2018; Zhao et al., 2018) (Table 2). Many natural products have also been tested and found effective in alleviating diseases such as AD (Deshpande et al., 2019). A soy protein Lunasin, acts as a neuroprotective agent that can block Aβ42mediated neurodegeneration in AD by downregulating JNK signaling (Sarkar et al., 2018; Deshpande et al., 2019). JNK and Hippo signaling act in a positive feedback loop mechanism(Irwin et al., 2020). Thus, the role of natural product(s) as putative therapeutic targets also holds immense potential in finding cures for neurodegenerative disorders.
The rationale of this kind of studies is to extend the information from the bench to the bedside. Understanding the molecular genetic mechanisms of a disease will allow us to identify the members of signaling pathways which exhibit aberrant signaling in a disease. Our understanding is further expanded from the chemical screens results, which will help in identifying chemical inhibitors that may block function of the signaling pathway members, for example, Hippo signaling in AD. These studies will have significant bearing on finding cures since potential therapeutic targets can be first tested in animal models (pre-clinical), followed by patient cell lines and then expanded to clinical trials. This bench to bedside modular approach to finding therapeutic targets holds immense potential as it cuts down the time and excess expenditure.
Conclusion
Although the relationship between cancer and neurodegenerative disorders is complicated but there are some common connecting links. Interestingly, Hippo signaling pathway can link the two important maladies like cancer and neurodegenerative disorders, which represent opposite ends of a spectrum. On one end of the spectrum is abnormal cell proliferation observed in cancer and on the other end there is abnormal cell death, the hallmark of neurodegenerative disorders. This further resonates with the existing notion that those who suffer with cancer generally have a reduced risk of developing a neurodegenerative disorder(s). Furthermore,patients undergoing chemotherapy regimen for cancer have also demonstrated cognitive impairment.
Since Hippo pathway has been discovered, extensive research has been conducted to study the role of Hippo pathway in development, growth, diseases and in other different contexts. Thus, Hippo exerts profound effects on cellular homeostasis. We understand that activation of Hippo signaling causes cytoplasmic sequestration of Yki/YAP/TAZ, which results in restricted growth response and apoptosis. Alternatively,through non-canonical pathway, YAP interacts with p73 inside the nucleus to trigger apoptosis. More components of Hippo pathway, regulatory mechanism and its relatedness to diseases still need to be elucidated. Our current understanding about the exact regulation and molecular mechanism of proper functioning of Hippo pathway associated with the neurodegenerative disorders is far from complete.
Multiple chemical inhibitors that block function of various components of the Hippo signaling pathway have been identified. Interestingly, majority of these studies emphasize on regulating the cell proliferation aspect of the Hippo signaling pathway. However, with emerging role of Hippo in neurodegenerative disorders, all these candidates can be tested for their efficacy in neurodegenerative disorders.These strategies will help in the development of effective and reliable therapeutic targets with the potential to block,suppress, or delay the onset of diseases.
Acknowledgments:We thank Bloomington Drosophila Stock Center(BDSC) for the Drosophila strains, and members of Singh lab for providing comments on the manuscript. We apologize to the peers whose work could not been cited because of space constraints.
Author contributions:Amit S conceived the manuscript. Amit S, NG, AVC,PD, Aditi S were responsible for manuscript writing, image preparation and editing.
Conflicts of interest:The authors declare no competing interests.
Financial support:The authors are supported by the University of Dayton Graduate program of Biology. This work was supported by NIH1R15GM124654-01 from the National Institutes of Health, Schuellein Chair Endowment Fund, and start-up support from the University of Dayton (to Amit S).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Shukla A, National Institute of Neurological Disorders and Stroke Laboratory of Molecular Biology, USA.

- 中国神经再生研究(英文版)的其它文章
- The use of hydrogel-delivered extracellular vesicles in recovery of motor function in stroke: a testable experimental hypothesis for clinical translation including behavioral and neuroimaging assessment approaches
- Advances in human stem cell therapies: pre-clinical studies and the outlook for central nervous system regeneration
- MicroRNAs in laser-induced choroidal neovascularization in mice and rats: their expression and potential therapeutic targets
- The emerging role of probiotics in neurodegenerative diseases: new hope for Parkinson’s disease?
- The phenotypic convergence between microglia and peripheral macrophages during development and neuroinflammation paves the way for new therapeutic perspectives
- Modeling subcortical ischemic white matter injury in rodents: unmet need for a breakthrough in translational research