
参与番茄叶形发育的TCP转录因子的表达及生物信息分析
2021-10-29 06:33张雪莹尹一歌
中国蔬菜 2021年10期
张雪莹 尹一歌 姜 晶 刘 欣
(沈阳农业大学园艺学院,设施园艺省部共建教育部重点实验室,辽宁省设施园艺重点实验室,辽宁沈阳 110161)
TCP 是植物特有的一类转录因子,其命名源自前3 个特征基因家族成员:玉米的teosinte branched 1(TB1)基因、金鱼草的cycloidea(CYC)基因、水稻的proliferating cell factors(PCFs)基因。TCP结构域是TCP 转录因子家族的保守域,是在调节植物生长发育的cyc和tbl的保守域中被发现的(Cubas et al.,1999)。此外,TCP 结构域在信号通路上有3 个磷酸化位点,该结构域是与增殖细胞核抗原(PCNA)基因启动子特异性结合以及与DNA结合所必需的,因此TCP 家族成员可以参与细胞增殖和细胞分化(Koyama et al.,2010)。
植物特异性转录因子TCP 家族可以在细胞分裂与细胞生长的耦合中发挥关键作用(Ingram &Waites,2006),而植物结构和形态的生长发育过程涉及到植物细胞的增殖、分化和生长(Li et al.,2005),因此TCP 可以通过影响细胞的增殖和分化来调控植物的器官形态(Aguilar &Sinha,2013)。已有大量研究表明,TCP 家族转录因子在植物生长发育的多个方面发挥作用,包括植株衰老、昼夜节律、激素信号转导(Manassero et al.,2013)、调控茎尖分生组织(shoot apical meristem,SAM)生长(Koyama et al.,2010)、叶片形态和大小的变化(Ori et al.,2007)以及侧枝的发育(Aguilar et al.,2007)等。AtTCP14在拟南芥种子的萌发过程中能够激活胚胎的生长潜能(Tatematsu et al.,2008),并且能和AtTCP15共同调节节间长度和叶片大小(Kieffer et al.,2011)。研究发现,AtTCP24对拟南芥次生细胞壁的增厚和花药内皮细胞的发育有调节作用(Wang et al.,2015)。在叶用莴苣(生菜)中下调LsTCP4的表达会造成叶细胞分裂异常,植株形成锯齿状或卷曲状的叶缘表型(Seki et al.,2020)。SlBRC1a(SlTCP9)和SlBRC1b(SlTCP7)在番茄腋芽中表达,其中任一基因活性降低都会使番茄分枝数增加(Martín et al.,2011)。
叶片作为植物体重要的营养器官,是由SAM生成的扁平侧器官。叶片发育分为3 个阶段:起始阶段、初级形态阶段、次级形态阶段(Kaplan,2001)。起始阶段,叶芽在SAM 侧面冒出;初级形态阶段,叶片横向膨胀,具有边缘胚芽区的物种会产生小叶(Berger et al.,2009);次级形态阶段,组织开始分化,叶片通过细胞扩张而面积增大。叶片的3 个发育阶段是复杂的、动态的过程,是连续的形态变化和分子水平的变化,因此研究叶片生长发育的分子机制是研究叶片形态变化的基础和前提。在植物中,与WUSCHEL相关的同源基因WOX1可以促进叶片外侧生长,而黄瓜CsWOX1突变体表现出异常的叶片形状和缺陷的次级叶脉,经证实,CsWOX1通过PINOID(CsPID1)控制生长素运输进一步在叶脉发育中发挥作用;同时,CsWOX1可能通过与CsTCP4相互作用影响细胞发育过程,进而改变叶片最终大小(Wang et al.,2020)。有研究发现,NGATHA(NGA)基因家族和STYLISH(STY)基因家族在叶缘的形成中起重要作用(Sohlberg et al.,2006;Martínez et al.,2014),通 过 研 究STY1、STY2双敲除以及NGA1、NGA2、NGA3、NGA4四敲除的拟南芥突变体表明,其叶片比相应野生型的更大、锯齿更多;相反,过表达STY1或NGA基因,突变体叶片具有更平滑的叶缘(Kuusk et al.,2002;Alvarez et al.,2009;Trigueros et al.,2009;Lee et al.,2015)。TCP2、TCP3、TCP4、TCP10、TCP24受miR319 的 调 控,高 表 达 的miR319 会下调以上5 个TCP基因表达水平,使拟南芥产生皱缩叶片(Palatnik et al.,2003)。有研究表明,同时下调NGAs和miR319 调控的TCPs,拟南芥叶片形态发生显著变化,导致叶片边缘不定生长(Alvarez et al.,2016)。在拟南芥中,TCP与包含LBD 域的asymmetric leaves 2(AS2)TF基因相互作用,抑制了KNOX基因在叶片发育过程中的表达(Li et al.,2012),而KNOX1是分生组织的特征基因,在叶片的起始发育阶段起作用(Bar &Ori,2014)。同样,参与植物生长发育的CUC基因家族 中 的TFs、CUC2、CUC3与AtTCP4之 间 相 互作用,调控叶片的复杂程度(Rubio-Somoza et al.,2014)。
番茄(Solanum lycopersicum)是一种世界范围内广泛栽培的经济作物(Teixeira et al.,2005),基因突变和外界生长环境的改变都会对其叶片造成影响(Kessler et al.,2001)。番茄的叶片为羽状复叶,叶片形状、叶面积大小及小叶数量是构成番茄叶片不同形态的重要因素(Shleizer et al.,2011)。目前在番茄中已经鉴定出30 个TCP 转录因 子(Parapunova et al.,2014),其 中lanceolate(LA,SlTCP2)能够调控番茄叶片生长已被证实。研究表明,下调LA的活性可以影响叶缘形态形成的时间(Aguilar et al.,2007),LA可以控制叶片的成熟时间和最终叶形(Shleizer et al.,2011),其突变体会产生小而简单的叶片。有研究发现,叶形与果实品质密切相关,叶片形态的改变对番茄果实可溶性固形物含量和产量有促进作用(Rowland et al.,2020)。因此,研究番茄叶片生长发育的调控机制对改善其产量和品质具有重要意义。本试验通过对可能参与番茄叶形发育的TCP 转录因子进行qRT-PCR,并结合生物信息分析对可能参与叶片发育的SlTCPs 转录因子进行筛选和特性分析,旨在找出影响叶片发育的关键TCP 转录因子,为进一步研究番茄叶片生长发育的分子调控机制奠定基础。
1 材料与方法
1.1 试验材料
试验于2019 年9 月至2020 年9 月在沈阳农业大学进行。供试番茄品种为Micro-Tom、VF36、Ailsa Craig(AC)、LA2532、LA2922,均 来 自 沈阳农业大学蔬菜分子生物学番茄课题组。55 ℃温汤浸种催芽后,在沈阳农业大学蔬菜分子生物学番茄课题组土培室进行培养。Micro-Tom 取顶端0.5 cm 长的新生叶片(幼嫩叶)和发育成熟、完整、展开叶片(成熟叶),液氮冷冻,-80 ℃保存备用。VF36、AC、LA2532、LA2922 取苗龄50 d 左右且发育成熟、完整、展开、没有病虫害的叶片,液氮冷冻,-80 ℃保存备用。
1.2 试验方法
1.2.1 RNA 提取和cNDA 合成 利用Trizol 法提取目标叶片的RNA,使用反转录试剂盒(PrimeScript ™RT reagent Kit with gDNA Eraser-Perfect Real Time,宝日医生物技术有限公司)合成cDNA,-20 ℃保存备用。
1.2.2 实时定量PCR 分析 利用Premier 5.0 软件,以候选目标SlTCPs 转录因子的序列设计引物(表1),委托苏州泓讯生物科技股份有限公司合成。以番茄组成性表达基因actin 为内参,利用SYBR Green PCR Master Mix(PE Applied Biosystems)试剂盒进行实时荧光定量(Real-time quantitative)PCR,所有反应进行3 次生物学重复,采用2-ΔΔCT算法并实时绘制熔解曲线,验证番茄叶片特异表达的候选目标SlTCPs 转录因子。利用IBM SPSS Statistics 进行显著性分析(P<0.05)。
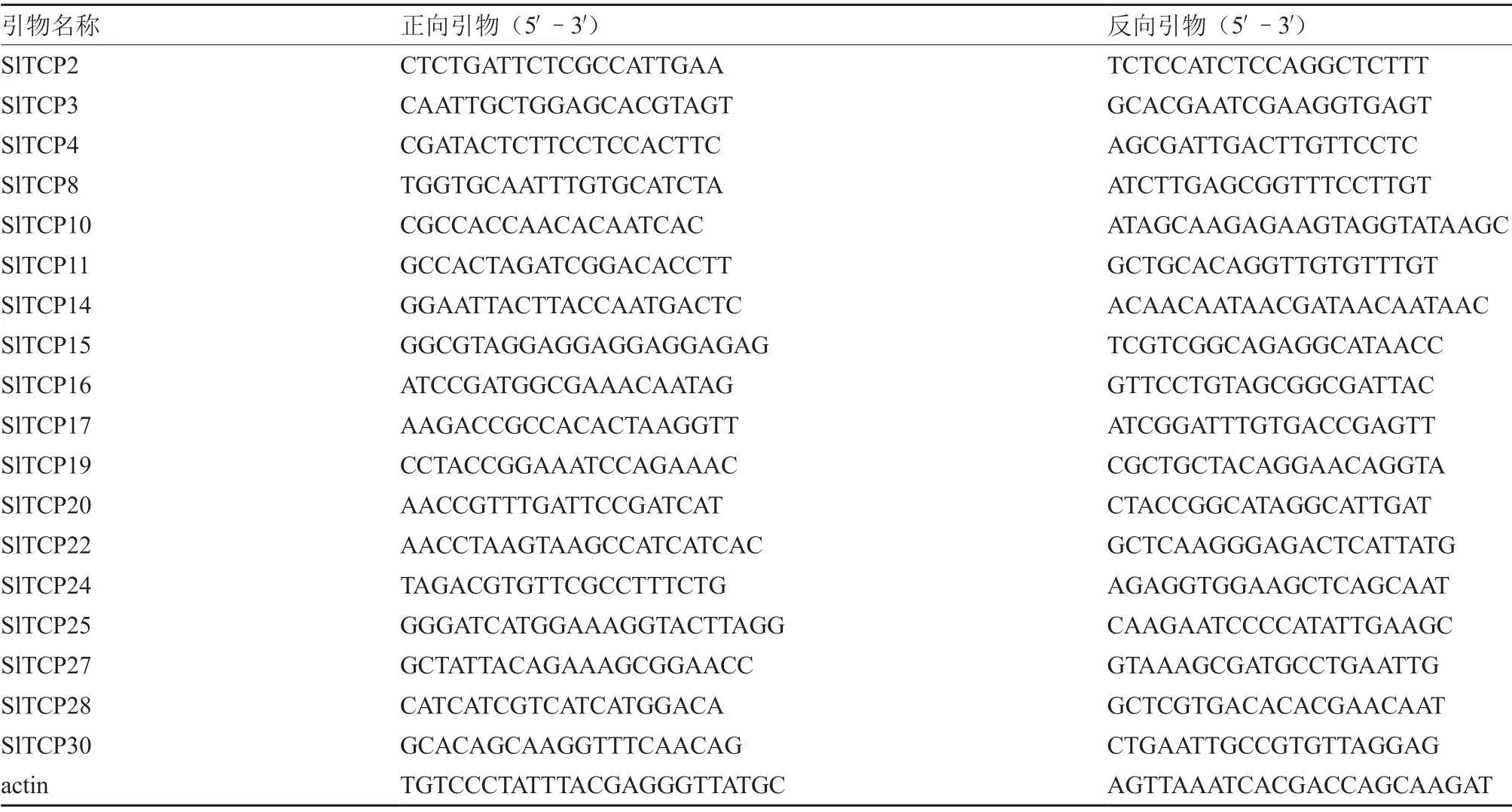
表1 SlTCPs 转录因子RT-PCR 引物序列
1.2.3 番茄TCP 基因转录因子基本信息获取 利用番茄数据库(https://solgenomics.net/)和NCBI数据库(https://www.ncbi.nlm.nih.gov/),查找筛选出参与叶片发育的候选目标SlTCPs 转录因子的基本信息,获得相应的cDNA 序列和蛋白序列并保存为FASTA 格式,利用DNAMAN 进行基因序列比对。
1.2.4 番茄TCP 蛋白质理化性质分析 将获得的候选目标SlTCPs 蛋白的氨基酸序列利用ExPASy(https://web.expasy.org/protscale/)的ProtParam tool分析氨基酸数量、分子量、等电点、不稳定系数、脂溶系数、疏水指数等理化性质。
1.2.5 番茄TCP 蛋白结构分析及亚细胞定位预测 利用DNAMAN 比对候选目标SlTCPs 蛋白一 级 结 构 序 列。利 用GenScript(https://www.genscript.com/psort.html)的PSORT 在 线 预 测候选目标SlTCPs 蛋白的亚细胞定位。利用PredictProtein(https://www.predictprotein.org/)在线预测候选目标SlTCPs 蛋白的氨基酸成分。
1.2.6 番茄TCP 的保守域、系统进化以及保守基序分析 候选目标SlTCPs 蛋白的保守结构域通过Pfam(http://pfam.xfam.org/)在线查找,并绘制成图。利用MEGA 7.0 构建系统进化树,选择邻接法,其他参数默认。利用MEME(http://meme-suite.org/)预测候选目标SlTCPs 蛋白的保守基序,输出的结构域数目为10,经过显著性分析后结构域数目纠正为7。
1.2.7 番茄TCP 的组织表达分析 以番茄功能基因组数据库TFGD(http://ted.bti.cornell.edu/cgi-bin/TFGD/digital/home.cgi)的RNA-Seq 数据为参考,研究候选目标SlTCPs基因在醋栗番茄(Solanum pimpinellifolium)不同组织和发育阶段的表达量。从TFGD 网站的RNA-Seq 数据列表中下载醋栗番茄的转录数据,包括不同开花期(0、10、20 d)、熟果期,以及子叶、下胚轴、分生组织、成熟叶、根、幼花蕾、幼叶等不同组织器官。利用MEV 绘制表达值(FPKM,Fragments per Kilobase Million)热图。
1.2.8 番茄TCP 基因启动子顺式作用元件预测分析 通过番茄数据库(https://solgenomics.net/)获取候选目标SlTCPs基因启动子序列(转录起始位置上游1 000 bp 序列)。通过PLANTCARE(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)在线分析预测顺式作用元件。
2 结果与分析
2.1 初步筛选与番茄叶片发育相关的候选目标SlTCPs 基因
根据番茄基因组数据库,共鉴定出30 个TCP 转录因子,其中有18 个在叶片中相对高表 达(Parapunova et al.,2014),本 试 验 检 测了18 个SlTCP基 因,即SlTCP2、SlTCP3、SlTCP4、SlTCP8、SlTCP10、SlTCP11、SlTCP14、SlTCP15、SlTCP16、SlTCP17、SlTCP19、SlTCP20、SlTCP22、SlTCP24、SlTCP25、SlTCP27、SlTCP28、SlTCP30在番茄叶片发育不同时期的表达水平。结果表明(图1),随着番茄叶片逐渐成熟,SlTCP11、SlTCP15、SlTCP19的表达水平极显著升高;而SlTCP2、SlTCP4、SlTCP8、SlTCP14、SlTCP22、SlTCP24、SlTCP25表达水平则显著或极显著降低。表明,SlTCP2、SlTCP4、SlTCP8、SlTCP11、SlTCP14、SlTCP15、SlTCP19、SlTCP22、SlTCP24、SlTCP25在叶片发育进程中可能具有重要的调控作用。对这10 个目标SlTCP基因进行进一步的生物信息学分析。
2.2 候选目标SlTCPs 的生物信息分析
2.2.1 番茄TCP 转录因子基本信息 通过番茄数据库和NCBI 数据库得到10 个与叶片发育相关的候选目标SlTCP基因的基本信息(表2)。这10 个候选目标SlTCP基因分布在8 条染色体上,其中1号、4 号染色体上各有2 个SlTCP基因。序列分析结果显示,SlTCPs基因的编码区(ORF)长度不等,在606~1 628 bp 之间,且多数>1 000 bp。在GenBank 中,SlTCP25基因的登录号空缺。
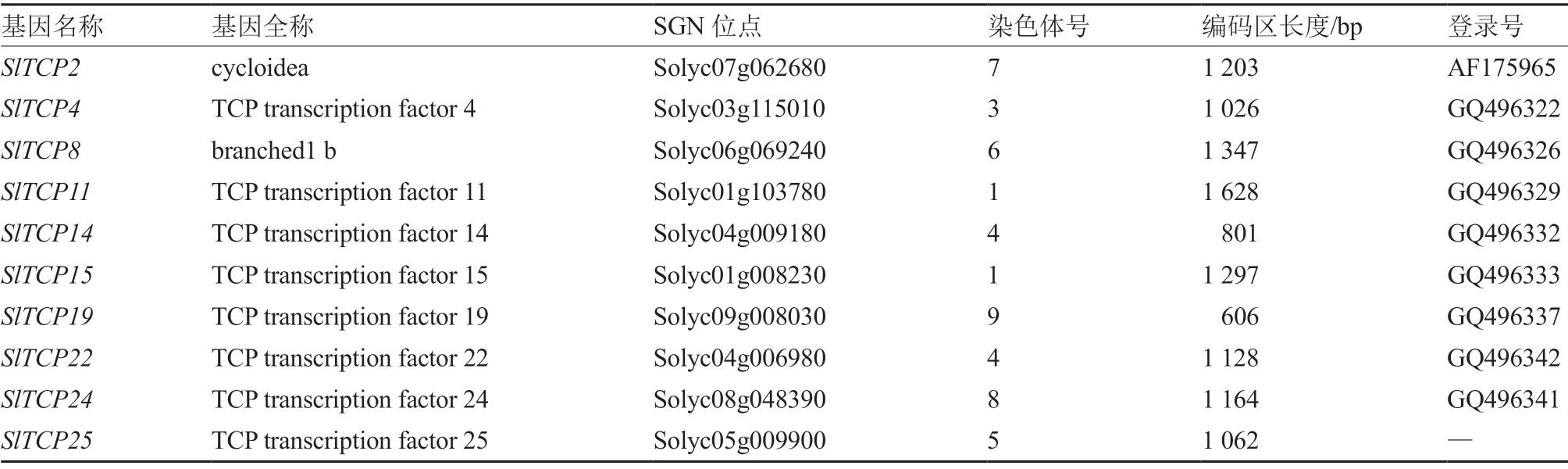
表2 番茄TCP 转录因子的基本信息
2.2.2 番茄TCP 蛋白理化性质分析 根据基因组序列分析验证10 个候选目标SlTCP 蛋白的理化性质。结果表明(表3),SlTCPs 蛋白对应编码的氨基酸长度在201~445 aa 之间,对应的分子量在21 246.00~47 550.61 Da 之间。SlTCP2、SlTCP4、SlTCP24 蛋白的等电点<7,显酸性;其余SlTCP蛋白的等电点>7,显碱性。10 个候选目标SlTCP蛋白的不稳定指数均大于40,均为不稳定蛋白;疏水指数均为负值,均为亲水性蛋白。

表3 番茄TCP 蛋白的理化性质
2.2.3 番茄TCP 蛋白结构、亚细胞定位及氨基酸成分分析 通过比对候选目标SlTCPs 蛋白序列发现,在10 个SlTCP 蛋白的氨基酸序列组成中,位于中间部分约50 aa 长度的氨基酸保守性相对较高,而两侧的序列保守性较低。从图2 可以看出,SlTCP11、SlTCP14、SlTCP15、SlTCP19 蛋白在保守域中的相同位点有碱基缺失,这可能是其发挥不同生物学功能的原因。
通过亚细胞定位预测发现,10 个候选目标SlTCP 蛋白均主要定位在细胞核上。对候选目标SlTCPs 蛋白的氨基酸主要组成成分进行分析,发现其中均含有丝氨酸(Ser),且所占比例相对较高(表4)。丝氨酸在植物生长发育中起重要作用,包括参与细胞组织分化、促进种子发芽等;其他主要氨基酸成分如丙氨酸、甘氨酸、赖氨酸、谷氨酸等对植物的光反应有影响,丙氨酸、亮氨酸、赖氨酸、脯氨酸、天冬氨酸等对植物响应胁迫有积极作用(宗和,2017)。
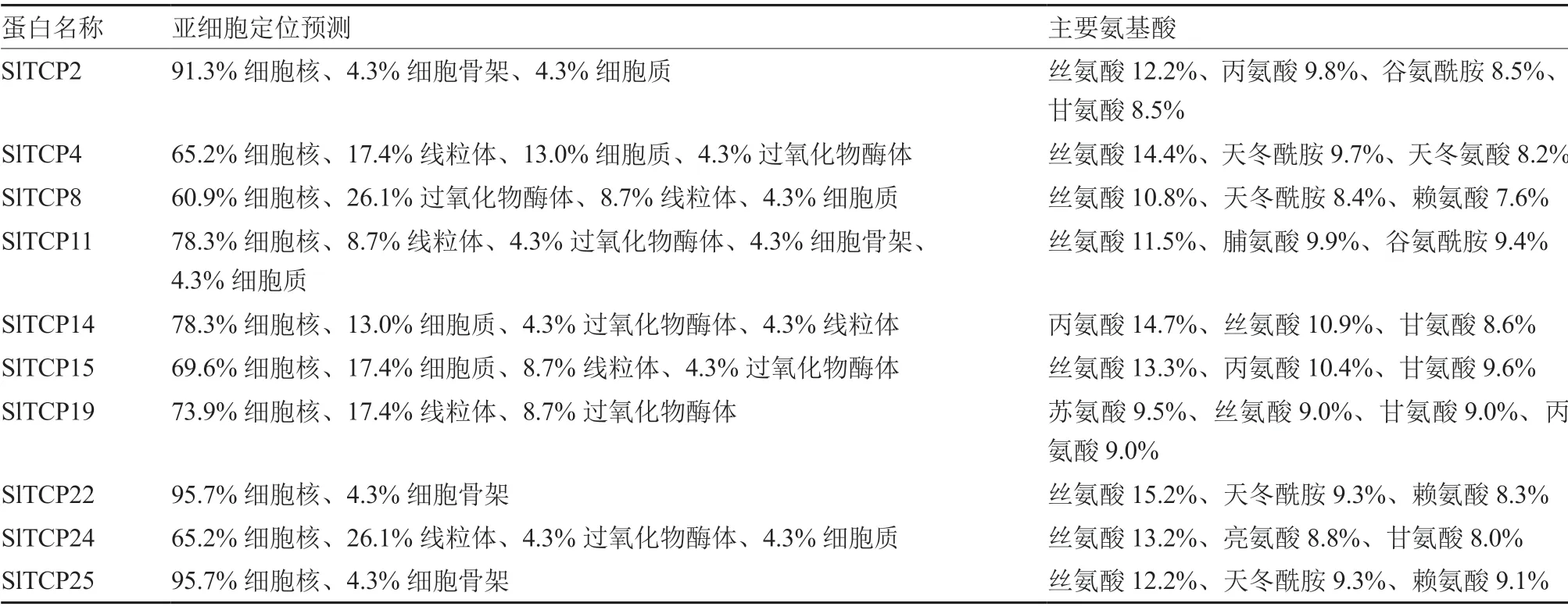
表4 番茄TCP 蛋白亚细胞定位预测和主要氨基酸
2.2.4 番茄TCP 蛋白的保守域、系统进化以及保守基序分析 利用Pfam 工具鉴定发现,候选目标SlTCPs 蛋白均含有不完整的TCP 结构域(图3)。10 个候选目标SlTCP 蛋白序列通过多重序列比对及采用邻接法构建系统发育树(图4)。结果表明,本试验构建的进化树与Parapunova 等(2014)研究所得进化树分支一致。这10 个基因分属TCP 家族的两大类别:Class Ⅰ和Class Ⅱ。SlTCP19、SlTCP11、SlTCP14、SlTCP15属 于Class Ⅰ;SlTCP2、SlTCP24、SlTCP4、SlTCP8、SlTCP22、SlTCP25属于Class Ⅱ,其中SlTCP2、SlTCP24、SlTCP4属 于CIN-like 亚 类,SlTCP8、SlTCP22、SlTCP25属于CYC/TB1 亚类。通过MEME 预测番茄TCP 蛋白的保守基序(图4),10 个候选SlTCP蛋白含有7 种主要基序,其中基序1 保守性最高,是TCP 蛋白的共有基序;SlTCP4、SlTCP24、SlTCP2、SlTCP8、SlTCP22、SlTCP25含有基序2,SlTCP19、SlTCP11、SlTCP14、SlTCP15含有基序5,分类结果与系统进化树的分支相同。同时,部分SlTCPs 在系统进化树中分支相近,表明其在功能上可能存在冗余。
2.2.5 番茄TCP 基因的表达分析 利用TFGD 网站在线分析候选目标SlTCPs基因在醋栗番茄中不同组织和发育阶段的表达模式。分析醋栗番茄不同发育时期不同组织器官的转录组数据发现,候选目标SlTCPs基因在不同组织器官中存在特异性表达,在不同发育时期的相同组织器官中的表达水平也存在明显差异(图5)。SlTCP11、SlTCP15、SlTCP24在各个时期的不同组织器官中,均有相对较高的表达水平;SlTCP2在花、未成熟果实、子叶、叶片中表达水平较高;SlTCP14在下胚轴、营养分生组织、花芽、幼嫩叶中表达水平较高。在成熟叶中,SlTCP2表达量最高,是SlTCP14、SlTCP15表达量的2 倍,但在幼嫩叶中SlTCP24表达量最高,是SlTCP15表达量的3 倍。而SlTCP25在所有组织器官中的表达量都很低,几乎不表达。其中SlTCP4、SlTCP8、SlTCP14、SlTCP15、SlTCP19、SlTCP22、SlTCP24在幼嫩叶和成熟叶中的差异表达水平与qRT-PCR 结果相一致;但SlTCP2、SlTCP11、SlTCP25在幼嫩叶和成熟叶中的表达水平相近,与qRT-PCR 结果略有差异。
对SlTCPs基因转录起始上游1 000 bp 的全部顺式作用元件进行分析,筛选后将其分成三类(图6)。在所有分析的候选目标SlTCPs基因中,光反应相关元件占比最大,为50.00%;环境胁迫相关元件占比约为45.35%,生长发育相关元件占比约为4.65%。SlTCP4含有最多数量的光反应相关作用元件,SlTCP2含有最多种类的胁迫相关作用元件,SlTCP4、SlTCP8、SlTCP25含有与生长发育相关的作用元件。可以看出,候选目标SlTCPs基因均可能参与到光反应路径中,并可能对胁迫应答、激素调节、生长发育起作用。
2.3 不同番茄叶形突变体中关键SlTCPs 基因的表达水平分析
番茄突变体LA2532 的叶片为叶裂加深,且有3~4 对二级小叶的复杂叶片结构,而突变体LA2922 的叶片为多个小叶整合到一整片叶的叶片结构(Hareven et al.,1996)。利用番茄两种不同叶形突变体LA2532、LA2922 鉴定SlTCPs基因的差异表达水平。结果表明(图7、8),与野生型相比,SlTCP8在两种叶形突变体中的表达水平要显著高于其他SlTCPs基因的表达水平。SlTCP4、SlTCP8、SlTCP22、SlTCP25等4 个SlTCP基 因在LA2532、LA2922 中的表达水平显著高于在野生型中的表达水平,表现出相同的表达趋势。但 是SlTCP2、SlTCP11、SlTCP14、SlTCP15、SlTCP19、SlTCP24在LA2532 中表达水平高于在野生型中的表达水平,而在LA2922 中表达趋势则与野生型相反。
3 结论与讨论
本试验在番茄已鉴定的30 个TCP 转录因子中(Parapunova et al.,2014),选出18 个在叶片中高表达的候选SlTCP基因作为研究对象,结合番茄叶片不同发育时期表达水平的差异,筛选出10 个SlTCP基因(SlTCP2、SlTCP4、SlTCP8、SlTCP11、SlTCP14、SlTCP15、SlTCP19、SlTCP22、SlTCP24、SlTCP25),并对其进行生物信息分析。同时,利用番茄两种叶形突变体LA2922、LA2532 对候选SlTCPs基因的表达水平进行分析,最终选出可能参与番茄叶形发育的关键基因SlTCP2、SlTCP11、SlTCP14、SlTCP15、SlTCP19、SlTCP24。
10 个候选目标SlTCP 的基因序列和蛋白序列分布在8 条染色体上,编码区(ORF)长度不等,且SlTCPs 蛋白表现出不同的酸碱性。SlTCPs 蛋白亚细胞定位预测均在细胞核上,推测其参与转录调控或转录后调控,可能对番茄生长发育进程造成直接影响。10 个候选SlTCP基因的启动子序列具有三类作用元件,可能参与番茄生长发育中的光反应、低温诱导、胁迫应答及激素调节。10 个候选SlTCP 蛋白均含有丝氨酸,且丝氨酸对植物的生长发育有积极作用(宗和,2017)。综上所述,10 个候选SlTCP基因均可能在番茄的生长发育过程中起到一定的作用。
根据TCP 结构域的同源性,TCP 蛋白可分为两大类:Class Ⅰ和Class Ⅱ。其中Class Ⅱ又分为两个亚类,即CYC/TB1、CIN-like。ClassⅠ和Class Ⅱ都包括转录激活子和转录抑制子(Manassero et al.,2013)。本试验中,SlTCP2、SlTCP4、SlTCP8、SlTCP11、SlTCP14、SlTCP15、SlTCP19、SlTCP22、SlTCP24、SlTCP25等10 个SlTCP 蛋白均含有TCP 蛋白保守结构域,且具有一段相同的保守基序。在蛋白序列比对中,SlTCP11、SlTCP14、SlTCP15、SlTCP19 与 其他SlTCPs 蛋白相比氨基酸序列有明显不同,且保守结构域相差较大。在构建的系统进化树中,SlTCP11、SlTCP14、SlTCP15、SlTCP19在同一分支上,其他SlTCPs基因在另一分支上,与前人的研究结果相符(Parapunova et al.,2014)。SlTCP2和SlTCP24在相同的分支Class Ⅱ上,同属CINlike 亚类,且SlTCP2和SlTCP24的理化性质相近,组织特异性表达的水平也相似。在进化树中,同一分支的SlTCPs基因结构更为相似,而基因结构越相似,其功能也越相似。在已报道的番茄TCP 家族中,SlTCP2可以调控叶片的发育进程及叶原基形成的动态变化(Shleizer et al.,2011),由此推测SlTCP24可能与SlTCP2在功能上有相似性。在已报道的参与植物叶形调控的TCP 转录因子中,属于Class Ⅱ分支上的TCP 转录因子较多,本试验筛选出的SlTCP11、SlTCP14、SlTCP15、SlTCP19同属于Class Ⅰ分支,蛋白结构上的差异可能会导致SlTCP11、SlTCP14、SlTCP15、SlTCP19在调控番茄叶形机制上较其他SlTCPs基因表现出差异。叶片主要依靠细胞增殖和细胞扩张来促进叶片生长(Manassero et al.,2013),而TCP 转录因子家族在细胞增殖、细胞分裂和细胞分化的调控中起重要作用(Koyama et al.,2010),进而影响植物叶片的形态结构和发育。TCP基因通过与许多分子和信号传导元件相互作用,在生物信号传导网络中发挥着核心作用(Dhaka et al.,2017)。例如,受miR319控制的TCP和NGA联合敲除对叶形发育有显著的加性影响(Alvarez et al.,2016);TCP和CUC通过竞争形成复合物来调控叶片形态(Rubio-Somoza et al.,2014)。这些特性使TCP成为研究植物中组合基因表达和激素串联机制的理想候选者(Dhaka et al.,2017)。
从组织特异性表达分析结果可以看出,SlTCP11、SlTCP15、SlTCP24在所有组织器官中均有相对较高的表达。随着叶片发育进程的变化,SlTCP4、SlTCP8、SlTCP14、SlTCP15、SlTCP19、SlTCP22、SlTCP24的表达水平明显发生改变,因此SlTCPs可能在叶片不同发育时期发挥不同作用。以上大部分SlTCPs基因与TFDG 网站预测的各个发育时期的表达水平相符,但SlTCP2、SlTCP11、SlTCP25在各个发育时期的表达水平与预测略有差异,因此分析造成其表达结果差异的原因可能是由于番茄品种不同及预测误差所造成。进一步研究发现,SlTCP2、SlTCP11、SlTCP14、SlTCP15、SlTCP19、SlTCP24在番茄两种不同叶形突变体的叶片中表达水平呈相反趋势,推测其可能对控制番茄复叶数量及叶裂形成具有重要作用。
目前TCP 转录因子在多种植物中被证明对植株生长发育,尤其是叶片形态的调控起重要作用,但是在具有复叶典型特征的番茄叶片中的研究还不够深入和广泛。已知SlTCP2对番茄叶片形态的构成具有重要调控作用(Shleizer et al.,2011),而其他SlTCPs基因在番茄叶片形态调控上的作用尚不清楚,本试验结果为探究TCP 转录因子家族在番茄叶片形态构成中的分子机制奠定了理论基础。关于番茄SlTCPs家族成员对番茄叶片发育的具体调控功能,还有待通过基因编辑和分子生物学等手段进行进一步深入研究。
猜你喜欢
农业工程学报(2022年1期)2022-03-25
今日农业(2021年21期)2022-01-12
英语文摘(2021年3期)2021-07-22
中国生殖健康(2020年7期)2021-01-18
小星星·阅读100分(低年级)(2019年10期)2019-11-26
家庭医学·下半月(2019年6期)2019-08-16
安徽文学(2019年6期)2019-06-28
作文大王·笑话大王(2018年12期)2018-03-23
奥秘(2015年4期)2015-09-10
红蜻蜓(2015年4期)2015-06-01
