
百山祖常绿阔叶林凋落量的季节变化及其驱动因素
2021-10-19 04:37:20张亚芬许大明蒋凯丽骆争荣
浙江林业科技 2021年5期
张亚芬,许大明,蒋凯丽,骆争荣
(1.丽水学院生态学院,浙江 丽水 323000;2.浙江凤阳山-百山祖国家级自然保护区百山祖管理处,浙江 庆元 323800)
森林生态系统是地球陆地生态系统的主体,它具有很高的生物生产力、生物量以及丰富的生物多样性[1]。在森林生态系统中,植物地上部分将自身的新陈代谢产物返回地上形成的一系列凋落物是连接森林生态系统地下和地上生态过程的关键纽带[2],对生态系统的养分循环起着非常重要的作用[3-4]。一般来说,森林凋落物主要包括枯枝落叶和因各种因素而凋零的植物繁殖和营养器官以及在植物生长代谢过程中所产生的脱离植物主体的一系列代谢产物[5]。凋落物作为微生物降解者的重要食物来源,在保障了整个微生物群落的稳定性的同时,也保证了植物群落养料的供给,使植株能够得到良好的存活条件,从而保障了整个生态系统的稳定[5-6]。
对森林生态系统功能的深入认识离不开对凋落物的研究[6]。森林凋落物的凋落强度是指森林生态系统中单位时间和单位面积上的凋落物总产量,是研究凋落物的重要方面,也是反映森林状况的主要指标[7]。近20年来,我国学者对各种森林类型的凋落量的时间动态进行了大量的观测研究[2,5–7],表明森林凋落量存在明显的林分、地域和季节差异[8-10]。常绿阔叶林作为我国亚热带中东部地区最重要的地带性植被,针对其开展的森林凋落物的长期监测研究仍然较少[11-12]。由于森林凋落量与年均温和年降水量等气候因子有关[8-10],而气候因子又存在较大的年际波动,对森林凋落量进行长期监测研究对于我们真正了解亚热带常绿阔叶林凋落量的季节节律具有十分重要的意义[11]。
气候条件是影响凋落量动态变化的关键因素之一。虽然在较大的时空尺度上,气候因素对我国天然林凋落量的影响已经有了较一致的共识[8],但在较小的时空尺度(如群落内和年内)上,气候因素对森林凋落量的影响存在很多差异[7,13-17]。前人在研究月凋落强度动态变化与气象(气候)因子的关系时,大多采用相关系数[13]、多元线性逐步回归[7,14-15]、主成分分析[16]或冗余分析[17]等统计方法。然而,不管是气象数据还是月凋落强度动态数据都可能存在时间自相关,从而影响上述方法的显著性检验。另一方面,先前的研究大多没有将植物凋落自身节律的影响从凋落强度季节动态和气候因素的关系中剔除。这会给研究结果的解读带来困难甚至偏差。例如,大部分研究都发现凋落强度在秋冬季会出现高峰。这在很大程度上都是由植物的生物学节律等遗传因素决定的,而非全部由秋冬季的低温导致。因此,在分析气候因子与凋落量动态的关系时必须排除上述两个因素的干扰。
本研究以位于百山祖国家公园亚热带中山常绿阔叶林内的5 hm2森林动态监测样地为平台,以该样地7年(2009—2016年)的凋落物产量监测和气象监测数据为基础,分析百山祖常绿阔叶林营养器官凋落物及其各组分(包括叶、小枝、树皮及它们的碎片组成的杂物)凋落强度的月变化,研究降水量和气温对月凋落强度动态变化的影响,以期为该植被类型的保护提供理论依据。
1 材料与方法
1.1 样地概况
本实验设置在位于浙江省百山祖国家公园的百山祖主峰北坡中山地带(海拔约1 400~1 600 m)的5 hm2(250 m × 200 m)固定森林样地中(27°40′54″N,119°03′53″E)。样地设在中亚热带天然常绿阔叶林内,该群落结构完整,物种丰富,区系复杂,受到的干扰相对较少[18]。样地内坡度大多在20°~ 35°,基岩为侏罗纪火成岩,土壤类型为黄壤,pH值在4.5~ 6.5。分别于2003年、2008年和2013年,对样地内所有胸径大于1 cm的木本植物进行了每木调查。调查结果显示,该群落的建群种为短尾柯Lithocarpus brevicaudatus和多脉青冈Cyclobalanopsis multinervis;鹿角杜鹃Rhododendron latoucheae的多度最高,在下木层中占优势;群落垂直层次比较复杂,分层不明显;由于海拔较高,群落中40.5%的种类为落叶树种,但落叶树种的重要值之和仅为15.1%,常绿树种的重要值之和为84.9%;整个群落仅4个针叶树种且其重要值均很低。百山祖地区位于暖温带海洋性群落交错区,属于亚洲的一个特殊区域[19]。受中亚热带季风性气候影响,降水充沛(年降水量为2 294 mm,多集中于4—6月),气候湿润(相对湿度为84%,无霜期为293 d),水热同步状况良好[20]。据地区气象站统计显示,保护区年平均气温仅为13.3℃,气温在-11.8~34.0℃之间,≥10℃的年均生长期积温为4 135℃[20]。
1.2 样品收集及处理
在5 hm2样地内的原有小路两侧分别摆放50个面积为0.71 m ×0.71 m方形凋落物收集器[2]。参照国内其他大样地凋落物收集器设置的方法,百山祖样地收集器设置原则如下:(1)在小道两侧3~ 10 m范围内交替设置;(2)相邻两个收集器之间的间隔大于10 m;(3)兼顾山脊、沟谷、林窗等特殊地形和生境(但并不是完全按比例设置)。为了工作方便,避免在距离胸径大于5 cm的树木半径1.5 m范围内设置收集器;收集器所占位置(0.5 m2)上无胸径大于1 cm的树木。收集器由聚氯乙烯管作支架,上面覆盖网孔为1 mm的玻璃纤维网做成,网深约0.33 m,以防止收集的凋落物外溢及受到污染,距离地面高度0.8~1.1 m。收集器在样地中的分布如图1所示。
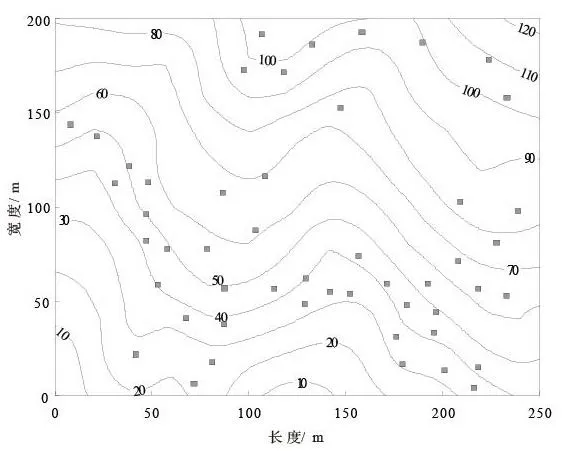
图1 百山祖监测样地凋落物(灰色正方形)收集器分布Figure 1 Location of 50 litter collectors at permanent sample plot in Baishanzu
自2008年8月开始,于每月月中(15日)和月底(30日)将落入收集器的凋落物全部收集起来装入大信封。将样品带回百山祖管理站后日光晒干以防腐烂,然后带回实验室放置于烘箱80℃烘干至质量恒定。烘干后,将这些凋落物样品进行分类鉴定(繁殖器官及营养器官)。
本研究仅分析营养器官凋落物,其组分主要包括植物的枝、叶和树皮。对于树叶,本研究主要鉴定9个优势种,即短尾柯、光叶水青冈Fagus lucida、木荷Schima superba、鹿角杜鹃、褐叶青冈Cyclobalanopsis stewardiana、多脉青冈、厚叶红淡比Cleyera pachyphylla、尖连蕊茶Camellia cuspidata、水丝梨Sycopsis sinensis的树叶;其余树叶再按针叶、其它常绿种类和其它落叶种类分类。小于1 cm2的无法鉴别的碎片(包括碎叶片、芽鳞片等)都计为杂物。枝条和树皮不再细分到种。
1.3 气象数据
2009—2016年的降水量和气温等气象数据来源于庆元县气象台设置于百山祖车根村(海拔约为1 100 m)的自动气象监测站。
1.4 样品数据分析方法
逐月统计2009—2016年所有收集器收集到的营养器官凋落物总质量、各凋落物组分质量,计算各组分的占比。样地月凋落强度为每月每平方米内样地的凋落物产量,计算时用每月收集到的凋落物总质量除以50个收集器的面积[21]。月凋落强度计算公式如下:
组分月凋落强度(g·m-2)=某凋落物组分某月的总产量(g)/(50×0.71×0.71 m2)
计算月均凋落强度时再按月份计算平均值,如1月平均凋落量为2009—2016年各年1月的凋落强度平均值。年凋落强度为各月份平均凋落强度之和。
本研究采用多元线性逐步回归分析降水量和气温对营养器官凋落物及各组分月凋落强度动态的影响。为避免植物器官凋落的生理性节律和气象因子变化的时间自相关对分析产生干扰,本研究先将自变量(单月总降水量和月平均气温)和因变量(各凋落物组分的月凋落强度)的原始数据分别减去该变量的当月平均值(如1月的平均值为2009—2016年各年份1月的平均值),然后进行回归分析。下文统一将经过上述数据处理的自变量和应变量分别称作去平均单月总降水量、去平均月平均气温和去平均月凋落强度。这样做的优点是可以将不同物种自身的凋落节律(只与物种本身的生物学特性有关)造成的影响去除掉,从而真正反映凋落和气候的关系。另外,去平均处理后,可以很大程度上去除变量的时间自相关。处理后的数据实际上更多地反映了相应指标的年际波动。
本研究采用R语言软件进行数据统计整理与分析。
2 结果与分析
2.1 营养器官凋落物的组成
对营养器官凋落物各组分的年均凋落强度分析(表1),发现凋落物中叶凋落物占了最大比例,达87.33%,其次为杂物,占6.76%,然后是枝条和树皮凋落物,分别占4.66%和1.25%。进一步将叶凋落物进行细分,短尾柯的占比最高,达到15.33%;依次为光叶水青冈,占14.01%、木荷占12.04%、鹿角杜鹃占10.76%、褐叶青冈占8.89%。从叶的习性来看,常绿树种叶凋落物占67.07%,落叶树种叶凋落物占19.69%,针叶树种仅占0.56%。
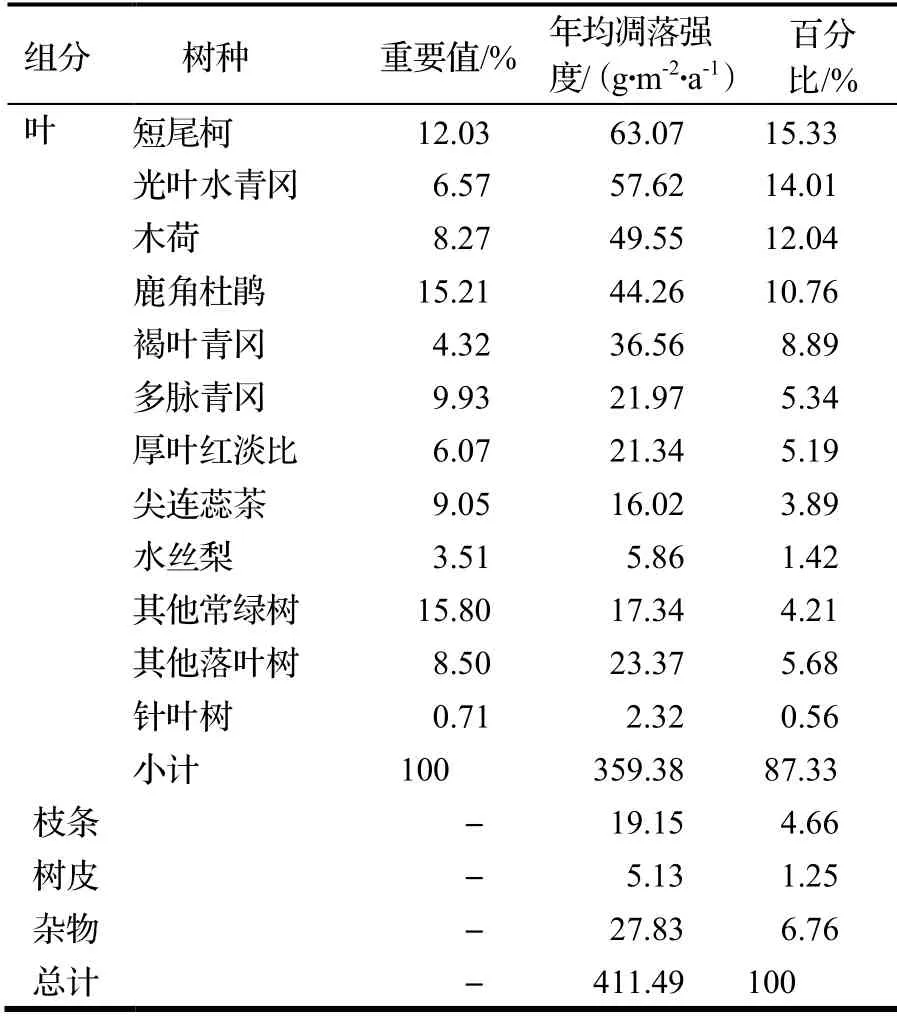
表1 百山祖常绿阔叶林凋落量的组成Table 1 Composition of litter yield at permanent sample plot
2.2 各凋落物组分月凋落强度的动态
由图2可知,百山祖常绿阔叶林营养器官凋落物月凋落强度呈现出明显的季节变化,为双峰型的季节动态。营养器官凋落物月凋落强度分别在4—5月和10月达到高峰。作为凋落物中占比最大的一部分,叶凋落物月凋落强度动态对营养器官凋落物月凋落强度的动态具有决定性影响。从各组分的动态来看,常绿树种主要在5月前后形成树叶凋落高峰,且在10月左右形成树叶凋落小高峰;落叶树种主要在10月左右出现一个树叶凋落高峰;针叶树种主要在12月左右大量落叶;枝条凋落物的凋落高峰期并不明显,2月、4月、8月的枝条凋落物月凋落强度有所升高;树皮凋落物月凋落强度的波动也不明显,一般在4月和7月有所升高。另外,分析结果也显示枝条和树皮凋落物月凋落强度的年际变化(标准差/均值)相对较大。
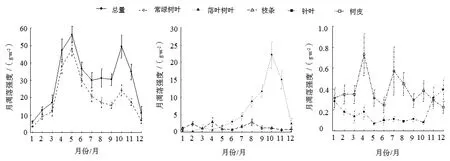
图2 百山祖常绿阔叶林凋落物总量及其组分凋落量的月际动态Figure 2 Month dynamics of total litter yield and each component yield at permanent sample plot in Baishanzu
进一步分析主要优势种的叶凋落物月凋落强度动态,发现各物种的叶凋落动态都有其各自的特性。鹿角杜鹃每年有2个凋落高峰,分别为5—6月和10月,其中第1个高峰期的时间跨度较大,月凋落强度也较大,在10月又出现一个小高峰;短尾柯叶凋落量主高峰也出现在5月,另外,在8月和10月出现了2个小高峰;多脉青冈的叶凋落高峰主要出现在4月,并且在11月出现小高峰;尖连蕊茶的叶凋落仅1个凋落高峰,出现在5月;木荷的叶凋落物月凋落强度动态呈现明显的双峰型,第一个凋落高峰出现在5月,第二个凋落高峰出现在10月,2个高峰时的月凋落强度较接近;作为优势种中唯一的落叶树种,光叶水青冈的叶凋落高峰主要出现在10月;厚叶红淡比的叶凋落高峰也很明显,仅1个凋落高峰,主要出现在5月;褐叶青冈也呈现出双峰型叶凋落动态,第一高峰较高,出现在4月,第二高峰较低,出现在10月;水丝梨的叶凋落高峰不明显,其叶凋落物月凋落强度于5—11月持续维持在较高水平(图3)。
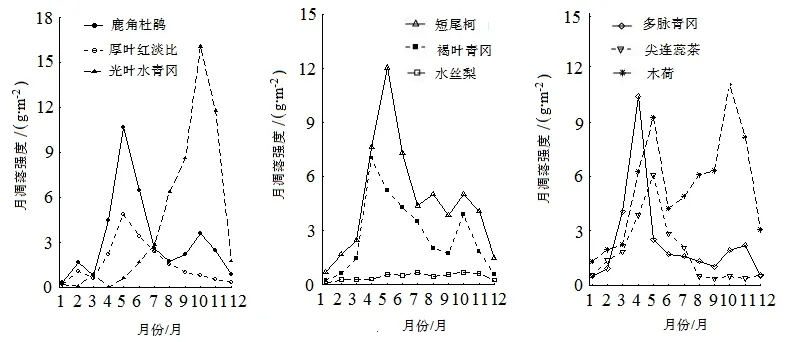
图3 百山祖常绿阔叶林主要优势种叶凋落强度的月际动态Figure 3 Month dynamics of leaf litter of dominant tree species at permanent sample plot in Baishanzu
2.3 月凋落物强度与气候因子的关系
多元线性逐步回归分析的结果表明(图4),去平均处理后,枝条和树皮凋落物、针叶树种树叶凋落物、落叶树种叶凋落物以及各优势种叶凋落物的月凋落物强度与气候因子(单月总降水量和月平均气温)都无显著关系,最优回归方程全部仅含常数项;仅营养器官凋落物和常绿树种树叶凋落物的去平均月凋落强度受去平均单月总降水量的影响。营养器官凋落物去平均月凋落强度(y)与去平均单月总降水量(x)的最优线性回归方程为:y=0.023x+0.444(r2=0.044,P=0.06);常绿树种树叶凋落物去平均月凋落强度(y)与去平均单月总降水量(x)的最优线性回归方程为:y=0.003x+0.012,(r2=0.092,P<0.01)。
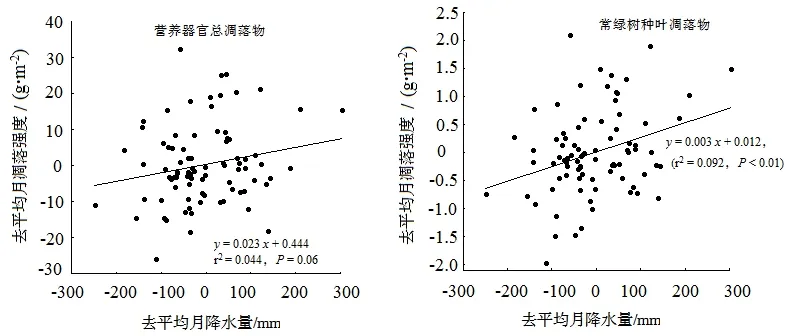
图4 营养器官总凋落量以及常绿树种叶凋落量与月降水量的关系Figure 4 Relationship between total litter yield and leaf litter of evergreen tree species with monthly precipitation
3 讨论
3.1 凋落物组分与季节节律
凋落物组成成分与群落的物种组成结构有关。本研究样地设置在百山祖中山常绿阔叶林内,常绿阔叶树种占有明显优势[18]。百山祖样地内9个优势种个体数多,胸高断面积大,它们的叶凋落物的凋落强度占到百山祖常绿阔叶林营养器官凋落物总凋落强度的76.87%。这些优势种的凋落节律综合作用在很大程度上影响了样地凋落物月凋落强度的年内动态[16]。与胡灵芝等对该样地2009年的凋落物监测数据分析得到的结果不同[20],本研究发现9个优势种的叶凋落物凋落强度占比排序出现了明显变化,尤其是排名前3位的叶凋落强度优势种从光叶水青冈>木荷>短尾柯变为短尾柯>光叶水青冈>木荷。百山祖常绿阔叶林凋落物及其各组分存在明显的年际变化[2]。相比于之前1个年度的观测数据结果,本研究分析了多年监测的数据,得到的结果更接近样地内物种优势度的排序情况,更能真实反映样地凋落物生产的情况。因此,延长观测和实验的年限对于准确理解凋落物凋落强度的时空异质性、组成及其影响因素是十分必要的[22]。
叶凋落物对整个生态系统都具有重要的生态作用。叶凋落物中的各类化学营养成分含量较高,同时可保温、保湿、涵养水土[4]。这在很大程度上保证了整个生态系统的物质循环稳定。与前人的研究结果类似[23-24],叶凋落物是百山祖常绿阔叶林营养器官凋落物的主体,占比达到了87.33%。对于不同生态习性的树种,树叶的凋落节律会有不同的表现[7]。在百山祖常绿阔叶林的样地中,常绿树种的叶凋落高峰主要在5月前后以及10月,而样地内的落叶树种的叶凋落高峰主要在10月左右。针叶树种在12月及1月出现叶凋落高峰。
森林凋落物季节动态一般可分为以下三种类型:单峰型、双峰型和多峰型(或不规则型)[7]。本次对百山祖常绿阔叶林样地中的9种优势种的叶凋落动态研究表明,单峰型凋落的物种较少,主要是尖连蕊茶和厚叶红淡比;木荷的叶凋落表现出了最明显的双峰型动态,于春末(5月)和中秋(10月)出现两大高峰;鹿角杜鹃也在这两个季节出现凋落高峰;来自壳斗科Fagaceae的优势种虽都为双峰型凋落节律,但凋落高峰出现的时间略有差异,多脉青冈、褐叶青冈的第一大高峰出现在仲春(4月),其次为中秋;而光叶水青冈作为落叶树种则与此相反,其第一高峰出现在中秋,第二高峰出现在仲春;短尾柯除在春末出现凋落高峰外,分别在夏末(8月)和中秋出现两个小高峰,短尾柯的叶凋落小高峰与花期和果期重合[19],叶凋落小高峰的出现可能与其开花结实消耗较多养分从而减少了叶的养分供应有关。物种间的叶凋落节律差异与种间亲缘关系可能存在一定相关性,有待于进一步的统计研究。
3.2 营养器官凋落物月凋落强度与雨热条件的关系
凋落物凋落强度是森林生态系统生产力的体现,受制于生物与非生物因素的综合作用[2,7]。气候因素对凋落物的产生具有重要影响[8]。武启骞等对6种温带森林凋落强度年内动态的研究表明,均温、积温和月降水量显著影响凋落物凋落强度的年内动态[14]。在沂蒙山区不同森林类型凋落物凋落强度动态的研究也表明森林凋落物月凋落强度受当月最低和最高气温的影响,阔叶树的月凋落强度受最低气温影响[15]。邓秀秀等对浙江天童亚热带常绿阔叶林的研究表明,凋落物总凋落强度和叶凋落物凋落强度与当月平均气温、最高温和最低温都呈现显著正相关[13]。本研究结果显示,当月平均气温对营养器官凋落物以及各组分的月凋落强度均无显著影响,而单月总降水量显著提高营养器官凋落物尤其是常绿树叶凋落物当月的凋落强度。这与先前的研究结果存在差异,也与本研究通过将各变量减去多年平均值的办法去除了植物器官凋落的生物学节律有关。剥离了植物自身生理节律所带来的凋落强度变化与气候因子周期性变化的干扰因素后,本研究能够更真实地反映气候因子对于凋落强度的影响。
温度过高或过低都会促进植物器官脱落。由于百山祖常绿阔叶林所处的海拔较高,很少出现极端高温。2009—2016年百山祖出现的最高温仅为34.0℃。这一温度不足以促进凋落。另一方面,百山祖常绿阔叶林树种的凋落节律适应了百山祖气温的年内波动,大部分阔叶树种在10月就达到落叶高峰,为后期可能出现的极端低温做好了准备。因此,在去除植物自身凋落节律的影响后,本研究未发现月平均气温对百山祖常绿阔叶林树木营养器官凋落物月凋落强度的影响。森林凋落量的形成是一个复杂的过程,不仅涉及器官的脱落,与树木光合作用产物的积累以及树木对同化产物的分配也有重要关系。有研究表明气象因素对亚热带常绿落叶阔叶混交林凋落物凋落强度季节动态的影响存在滞后[16]。本研究的结果仅说明温度对百山祖常绿阔叶林营养器官的脱落无显著影响,而温度对常绿阔叶林树木的光合产物积累和同化产物的分配的影响有待进一步研究。
本研究发现单月总降水量因素对于对百山祖常绿阔叶林营养器官凋落物的凋落强度具有显著影响。单月总降水量对营养器官凋落物凋落强度成正相关关系,尤其是对常绿树种叶凋落物凋落强度的影响较大。这也意味着降水越大,对整片森林树叶凋落也会有所影响。正常降水对常绿阔叶林的生长是有益的,但降水量过高时,雨水可形成较强的冲击力损毁植物枝叶。很多学者认为台风带来的短期强降雨和大风是影响我国东部沿海地区森林的树叶凋落量的重要因素[13,19,25]。
4 结论
本研究利用百山祖常绿阔叶林2009—2016年的凋落物动态监测数据分析气候因子对该常绿阔叶林营养器官凋落物月凋落强度的影响。研究结果表明,排除凋落物自身凋落节律和气候因子的时间自相关后,仅单月总降水量会影响营养器官凋落物,尤其是常绿树种叶凋落物月凋落强度,月平均气温对营养器官凋落物及其组分的月凋落强度无显著影响。
树木营养器官凋落量的形成是一个复杂的过程,当地光照、风力和土壤条件以及凋落前较长一段时期内(不仅是当月)的降水量和气温都有可能通过影响植物的光合作用、有机物分配和器官脱落等生理过程而影响森林凋落物的凋落强度。戴雯笑等的分析表明百山祖常绿阔叶林凋落物产量存在较大的年际变化[2]。这种变化可能更能综合反映各气象因子的影响。今后,需要积累数据在这方面的开展更深入系统的研究。
猜你喜欢
广西糖业(2024年1期)2024-04-25 17:59:28
今日农业(2021年8期)2021-07-28 05:56:04
书香两岸(2020年3期)2020-06-29 12:33:45
绿色中国(2019年18期)2020-01-04 01:57:08
农家科技下旬刊(2017年8期)2017-11-13 18:09:22
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:01
江苏农业科学(2017年1期)2017-02-27 10:14:38
卫星与网络(2016年12期)2016-02-05 09:23:22
江苏农业科学(2014年7期)2014-09-02 11:04:02
华东师范大学学报(自然科学版)(2014年1期)2014-04-16 02:54:55
