
基于三江源高寒草甸群落结构变化评估围栏封育对草地恢复的影响
2021-10-12 01:12:42李宝林袁烨城祁佳丽
生态学报 2021年18期
刘 岩,李宝林,*,袁烨城,祁佳丽,李 影,李 睿
1 中国科学院地理科学与资源研究所 资源与环境信息系统国家重点实验室,北京 100101 2 中国科学院大学,北京 100049 3 国家环境保护青藏高原生态环境监测与评估重点实验室,西宁 810007
三江源是中国江河中下游地区以及东南亚国家生态安全和区域可持续发展的重要生态屏障[1]。作为国家重点生态功能区的重要组成部分,过去几十年间三江源在全球气候变化与人类活动的双重影响下,生态系统退化明显[2-3]。鉴于此2005年和2014年国务院分别规划投资75亿元和161亿元启动了三江源生态环境保护与建设工程一期和二期工程,实施退牧还草、黑土滩治理、湿地保护等生态保护与修复项目[4-5]。因此,客观评价区域生态质量状况及其变化,对国家生态保护成效评估及生态保护政策的制定具有重要意义。
遥感数据覆盖范围广泛且获取方便,是进行大区域生态评估常用的技术手段。目前在三江源地区进行的生态评估主要是利用遥感手段和地理信息系统技术,侧重从生态系统类型、生态系统生产力和生态系统服务角度进行评估。在国家实施三江源保护工程后,三江源地区生态系统格局稳定少动,水体与湿地生态系统整体有所恢复[6-9];植被指数、净初级生产力、地上生物量等表征草地生产力指标的监测也表明植被呈现恢复趋势,生态状况逐渐改善[10-19],但也有一部分研究结果表明高寒草地生产力下降或基本保持稳定[20-23];另外,生态系统服务功能也呈好转趋势[24-26]。
遥感技术虽然对三江源区域宏观生态状况评估具有优势,但无法反映生态系统结构的变化。生态系统整体生产力提高,不一定代表着草地群落结构向好的方向转变。目前对三江源草地群落结构变化评估主要是基于地面样方调查数据来进行,多数研究结果表明围栏封育提高了地上生物量,草地禾本科和莎草科等优良牧草增加,生态状况改善[10,16-17,27-30]。但部分学者也发现部分地区在围栏封育后,虽然地上草地盖度与地上生物量恢复,但没有改变杂草的主导地位,莎草科等优良牧草覆盖度减少[31-33],杂草生物量比禾本科增加幅度更大,甚至出现了禾本科和莎草科植物几乎消失的情况[34]。围栏封育导致毒杂草比例增加在其他区域的研究中亦有过很多报导,Li等在内蒙古高原沙漠化草地禁牧26年后发现一年生杂草显著增加并在群落中占据了主导地位[35],1998—2016年在内蒙古进行的禁牧实验发现在长达18年禁牧后杂草生物量占比显著增加,而禾本科的百分比显著减少[36],在美国[37-38]、土耳其[39]、捷克[40]、苏格兰[41]、伊朗[42-43]等地区的围栏实验也观测到了禁牧后杂草生物量占比显著提高的现象。从现有的研究成果来看,围栏封育并不一定会改善草地植物群落结构。在三江源地区,由于地面监测数据获取困难,现有研究多样点数量有限且缺乏连续长时间的观测,给三江源区围栏封育效果评价带来了很大的不确定性。本文拟基于多监测站点长时间序列草地各经济类群地上生物量的变化来评价三江源草地群落结构的变化,从而评估三江源地区生态保护与修复措施的成效,为生态保护与修复绩效评估以及生态保护政策制定提供可靠的依据。
1 研究方法
1.1 研究区概况
三江源是指长江、黄河、澜沧江的源头地区,位于中国青海省南部,地理位置介于北纬31°39′—36°16′、东经89°24′—102°23′之间,总面积约36.3万km2,行政区域包括玉树、果洛、海南、黄南四个藏族自治州的16个县和格尔木市的唐古拉山乡(图1)。研究区以山原和峡谷地貌为主,海拔4000—6000m,西北-东南走向的巴颜喀拉山和阿尼玛卿山将三江源分为以果洛州为主体的东北部黄河源区和以玉树州为主体的西南长江、澜沧江源区,地势由东南向西北逐渐升高。三江源属典型的高原大陆性气候,冷暖交替、干湿分明、水热同期,年平均气温为-5.6—7.8℃,年总降水量在262—772mm之间。植被类型以草地为主(面积占比约为68%),包括高寒草甸、高寒草原、沼泽湿地和温性草原等[13],另外还分布有针叶林、灌丛、高山植被等。土壤类型按海拔从高到低依次分布有高山寒漠土、高山草甸土、高山草原土、山地草甸土、灰褐土、栗钙土和山地森林土,沼泽化草甸土也较为发育,冻土分布广泛。虽然在三江源生态保护工程开始实施后开始减畜,但三江源地区草地多年平均载畜总量高达2003.7万羊单位,仍处于超载状态[44]。
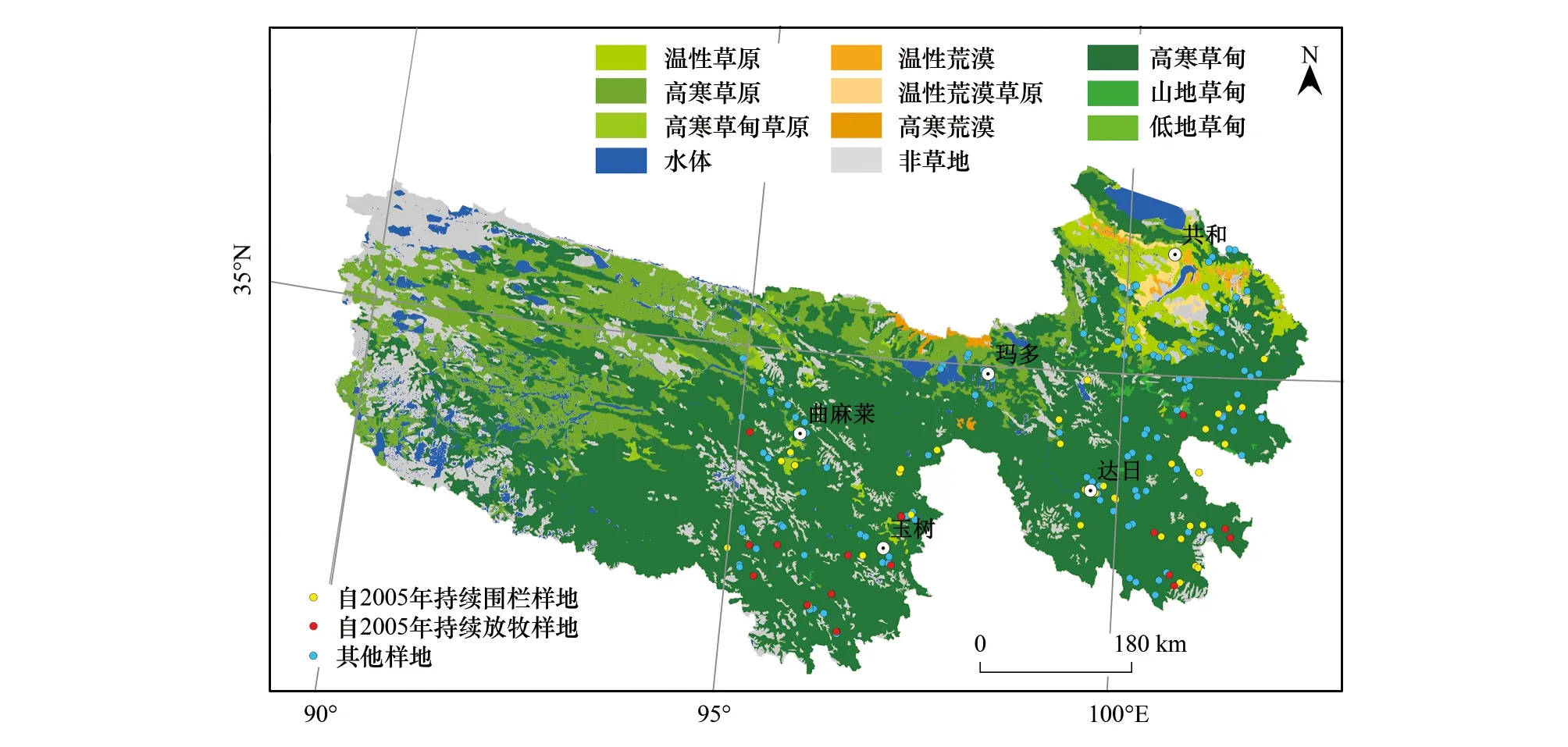
图1 研究区地理位置、植被类型及采样点分布图Fig.1 The location,grassland type and sampling points of the Three River Headwater Region
1.2 数据来源及预处理
研究中使用的数据包括牧草营养枝高度、生殖枝高度、植被盖度以及地上生物量四个指标,数据来源于青海省草原总站,数据获取的时段为2005—2017年。数据采集时间一般在夏季天然草地牧草生长盛期,多为8月中上旬。样地面积不小于1km2,每个样地布设3—6个样方,样方之间的间距为250m左右,样方面积为1m×1m。地上生物量测定时在样方内将牧草齐地面刈割,按禾本科、莎草科、豆科、可食杂草、不可食杂草和毒草等经济类群分别称重,其中禾本科和莎草科牧草统计的是牲畜喜食优良牧草,两个科中的毒杂草统计到其他相应的各毒杂草经济类群中。在各经济类群生物量基础上,统计了总生物量和可食草总量以反映草地总生产力与经济效用,统计了禾本科和莎草科牧草总生物量以及毒杂草总量来反映植被退化状态。另外,为反映围栏封育对草地植物群落结构的影响,还计算了各经济类群、可食草总量、毒杂草总量以及禾莎总量占总生物量的比例。
2005—2017年三江源区共布设了227个样地,由于布设的样地是逐年增加的,样地数量在不同年份差异较大。为保证数据的可比性,本研究只选择了研究时段内序列完整的监测数据。其中,围栏封育样地从三江源保护工程开始实施的2005年开始围封,且在研究时段内持续禁牧,放牧样地从2005年开始持续放牧。在围栏和放牧样地中分别有39个和18个满足要求,其中包括八个对照样地,样点分布如图1所示。在数据分析时,对比围栏内外各个指标统计量大小时采用了8对对照样地数据。为尽量反映三江源围栏封育整体效果,在对比围栏内外变化趋势时,围栏内外都采用了全部样地数据进行分析。
1.3 数据分析
采用Shapiro-Wilk检验和Kolmogorov-Smirnov检验,对牧草营养枝高度、生殖枝高度、盖度、各经济类群生物量、可食草总量、毒杂草总量、禾莎总量、总生物量以及各经济类群、可食草总量、毒杂草总量以及禾莎草总量占总生物量比例等指标进行统计检验后,发现多不服从正态或对数正态分布。因此对各指标在围栏区与放牧区总体大小差异进行统计检验时采用了Mann-Whitney U检验方法,总体大小比较时统计量采用中位数。
除了判断围栏区与放牧区各指标总体大小差异以外,本文还采用最小二乘法拟合各指标中位值的时间序列变化趋势,采用F检验对拟合方程进行可靠性检验,并在此基础上比较围栏区与放牧区各指标总体大小变化趋势的差别,具体计算公式如下:
(1)
ACRi=TCRi/(tn-t1)
(2)
y=a+β×t
(3)
式中,TCRi表示研究时段内第i个指标总变化率,ACRi表示研究时段内第i个指标年变化率,t1和tn分别为研究时段的起始和终止年份。y为某指标值,t为年份,a和β为拟合方程的参数,采用最小二乘法拟合获得。
2 研究结果
2.1 主要指标总体大小
2.1.1高度、盖度与总生物量
围栏封育显著提高了牧草的高度和地上生物量,盖度略有提高但不显著。2005—2017年从围栏内外对照数据来看,围栏内营养枝平均高度、生殖植平均高度和地上总生物量都显著高于围栏外对照区。围栏内营养枝平均高度中位值为6.7cm,围栏外为6.0cm,围栏内比围栏外高11.6%(P=0.043),围栏内生殖枝平均高度中位值为17.5cm,围栏外为15.1cm,围栏内比围栏外高15.9%(P=0.008),围栏内地上生物量中位值为4569.8kg/hm2,围栏外为3741.1kg/hm2,围栏内比围栏外高22.2%(P<0.001)(表1)。

表1 围栏与放牧区主要指标总体大小差异Table 1 Significance test between two averages of main indicators between fenced and grazed samples
2.1.2各经济类群生物量
2005—2017年只有禾本科、可食草总量和禾莎总量在围栏封育区和放牧区有显著的差异(P<0.05),而莎草科、豆科、可食杂草、不可食杂草以及毒草则没有显著差异。围栏封育区禾本科生物量中位值为1187.7 kg/hm2,放牧区为792.2kg/hm2,围栏区比放牧区高49.9%。围栏封育区可食草生物量中位值为3554.5 kg/hm2,放牧区为2861.7kg/hm2,围栏区比放牧区高24.2%。禾莎总量在围栏封育区中位值为2207.3 kg/hm2,在放牧区为1654.5 kg/hm2,围栏区比放牧区高33.4%(表2)。

表2 围栏与放牧区各功能群生物量总体大小检验Table 2 Inspection of the difference in biomass of functional groups in fenced and grazed sites
2.1.3各经济类群生物量占比
2005—2017年6个经济类群中莎草科生物量占比最高,在围栏封育区和放牧区分别为26.7%和28.6%;其次为可食杂草,占比分别为25.4%和24.5%;禾本科占比分别为22.2%和20.1%;不可食杂草为12.3%和13.0%;毒草占比分别为9.8%和10.1%;豆科牧草占比最小,都为3.6%。各经济类群生物量占比在围栏封育区和放牧区呈现出相似的规律,各个指标在围栏内外都没有显著的差异(表3)。
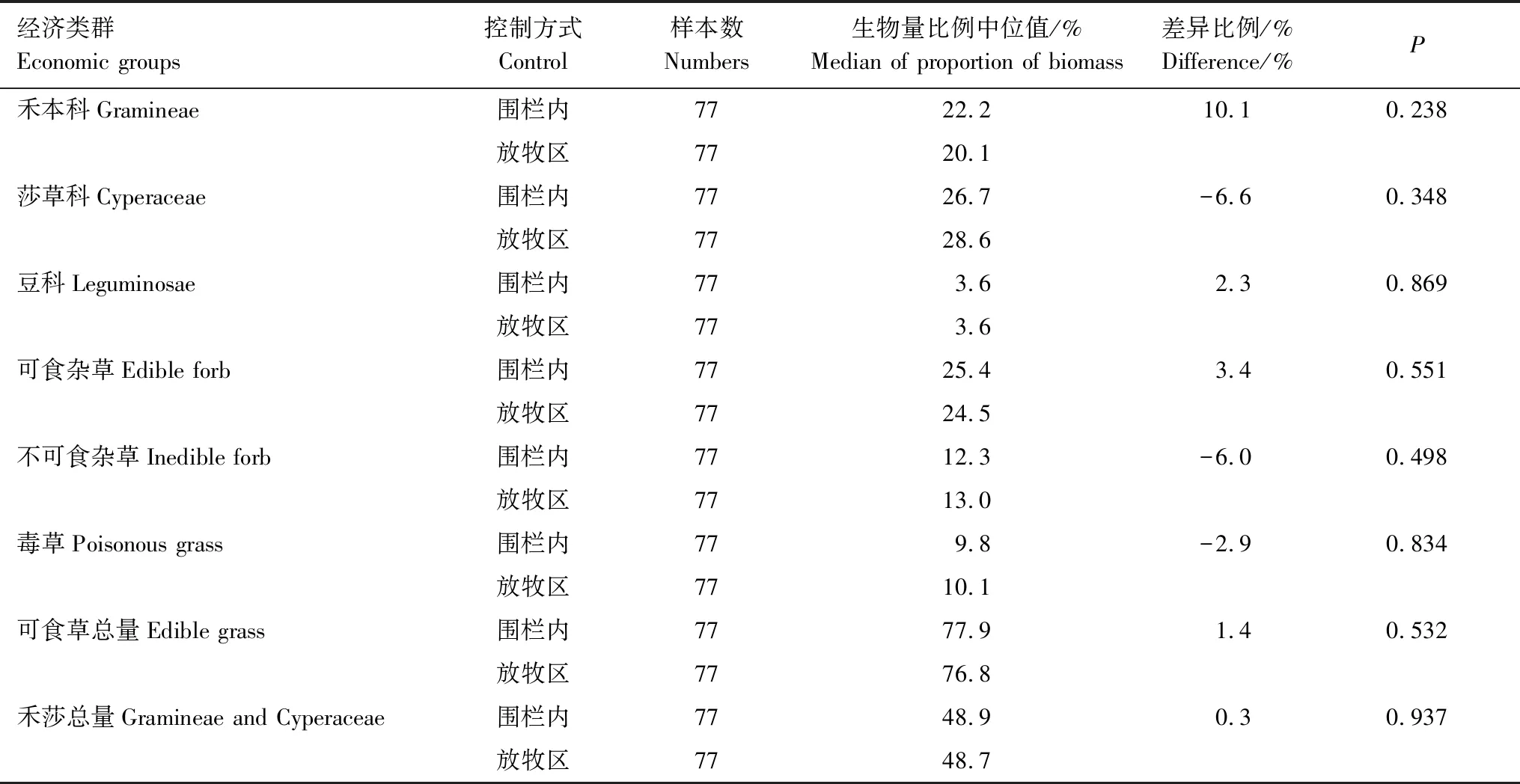
表3 围栏与放牧区各功能群生物量比例总体大小Table 3 The difference in biomass proportion of functional groups in fenced and grazed sites
2.2 主要指标变化趋势
2.2.1高度、盖度与总生物量
2005—2017年总生物量在围栏封育区和放牧区都没有显著的趋势性变化(P>0.1),营养枝平均高度、生殖枝平均高度与植被盖度在围栏封育区都呈现出显著的减少趋势(P<0.05),而在放牧区变化趋势都不显著(P>0.1)。在围栏封育区,研究时段内围栏封育区营养枝平均高度减少了34.8%,平均每年减少2.9%;生殖枝平均高度减少了38.4%,平均每年减少3.2%;植被盖度减少了4.8%,平均每年减少0.4%(图2)。

图2 围栏封育区与放牧区主要指标变化趋势 Fig.2 Change trend of main indicators in fenced and grazed sites表示所用拟合方程的显著性水平;**表示显著(P<0.05),***表示非常显著(P<0.01)
2.2.2各经济类群生物量
2005—2017年各经济类群生物量多数未表现出明显的趋势性变化,只有毒草和可食杂草在围栏封育区和放牧区表现出趋势性变化(P<0.1)。在研究时段内围栏封育区毒草生物量共增加了335.0%,平均每年增加27.9%;可食杂草增加了26.2%,平均每年增加2.2%。在研究时段内放牧区毒草增加了136.6%,平均每年增加11.4%;可食杂草减少了29.5%,平均每年减少2.5%(图3)。

图3 围栏封育区与放牧区各经济类群生物量变化趋势Fig.3 Change trend of biomass of different economic groups in fenced and grazed sites表示所用拟合方程的显著性水平,*表示边际显著(P<0.1),**表示显著(P<0.05)
2.2.3各经济类群生物量占比
2005—2017年各经济类群生物量占比多数表现出明显的趋势性变化(P<0.1),但在围栏封育区与放牧区具有明显的差异。在5个基本经济类群中,围栏封育区禾本科、莎草科、可食杂草和毒草各功能群生物量占比有显著的趋势性变化;放牧区可食杂草、不可食杂草与毒草具有明显变化趋势(P<0.1),豆科牧草生物量占比在围栏封育区和放牧区都没有显著的变化趋势。在围栏封育区和放牧区,可食草总量都具有显著的变化趋势,而禾莎草总量则只有围栏封育区具有显著的变化趋势(P<0.1)。
在围栏封育区禾本科和莎草科生物量占比都呈显著减少趋势,研究时段内分别减少了48.2%和23.9%,平均每年分别减少4.0%和3.3%;毒草占比增加了230.2%,平均每年增加19.2%;可食杂草增加了42.0%,平均每年增加3.5%;禾莎草总量占比减少了31.2%,平均每年减少2.6%。在放牧区,可食杂草占比在研究时段内减少了37.7%,平均每年减少3.1%;不可食杂草占比增加了113.9%,平均每年增加9.5%;毒草在研究时段内增加了170.3%,平均每年增加14.2%。围栏封育区和放牧区的可食草总量占比在研究时段内分别减少了13.5%和14.9%,平均每年减少1.1%和1.2%(图4)。

图4 围栏封育区与放牧区各经济类群生物量占比变化趋势Fig.4 Change trend of biomass ratios of different economic groups to total biomass in fenced and grazed sites表示所用拟合方程的显著性水平,*表示边际显著(P<0.1),**表示显著(P<0.05),***表示非常显著(P<0.01)
3 讨论
3.1 围栏封育区群落构成变化形成机制
围栏封育区草地地上生物量在时间序列上表现为波动中保持稳定(图5),但其内部种群结构发生了变化。具体表现为减少未退化状态下作为建群种的禾本科和莎草生物量占比,增加杂草与毒草占比。研究时段内禾本科和莎草科生物量占比分别减少48.2%和23.9%,毒草增加了230.2%,可食杂草增加了42.0%,而从整个可食草量占比来看,也降低了13.5%(图3)。从放牧区来看,禾本科和莎草科生物量占比都较稳定,未呈现显著性变化,生物量占比变化主要通过减少可食杂草增加不可食杂草和毒草来实现(图4)。研究时段内,可食杂草减少了37.7%,不可食杂草和毒草分别增加了14.6%和170.3%。可见围栏封育区尽管总生物量保持稳定,但群落结构已呈现出显著的恶化趋势,且比同期自由放牧区恶化更为突出。这与Li等[31]在三江源的研究结果类似。

图5 围栏封育区与放牧区地上生物量年际变化Fig.5 Variation of aboveground biomass in fenced and grazed sites
围栏区莎草科生物量占比下降主要是由于莎草科牧草在与禾本科和毒杂草对光照的竞争中处于劣势。在放牧压力消除后,由于禾本科牧草和毒杂草等多植株高大,枝叶繁盛,在光热资源竞争中处于优势地位[45-46],而植株低矮的莎草科牧草作为下繁草,由于植株被遮阴导致光合速率降低[47],在竞争中处于弱势地位,尽管不再被牲畜啃食,生物量占比也会逐年下降。
毒杂草尤其是毒草快速增长,首先与放牧压力消除后其对光热资源的强竞争能力有关。青藏高原的毒杂草主要有黄花棘豆(Oxytropisochrocephala)、狼毒(Stellerachamaejasme)、醉马草(Achnatheruminebrians)、黄帚橐吾(Ligulariavirgaurea)等[48]。毒杂草大多株体较高,具有繁茂的叶层以及较大的比叶面积,在与禾本科和莎草科尤其是莎草科植物对光热资源的竞争中处于优势地位,导致天然草地植物禾本科和莎草科牲畜喜食的优良牧草逐渐减少,草地经济价值逐渐降低[49-50]。其次,由于放牧压力的消除,毒杂草不再遭受牲畜的踩踏,植株相对较高大的茎不再折断,叶层也就更为繁盛,使禾本科与莎草科这些耐牧牧草进一步丧失了竞争优势。另外,毒杂草多以有性生殖为主,繁殖能力强,且生长迅速,适应性强,进一步加剧了毒杂草的蔓延,从而使毒杂草生物量占比逐渐增加[51-53]。
禾本科牧草在围栏封育后占比降低,主要与其同毒杂草竞争中失去对光热资源的竞争优势有关,可能还与禾本科与莎草科分布区地表会产生更多枯落物有关。天然草地中禾本科与莎草科植株多呈现为紧伏地表的流线型垫状体,有效地减少了植物体的受风面积,可以保留更多的枯落物,而毒杂草植株高大且比叶面积大,在枯黄后枯落物尤其是叶子会快速被秋冬季大风所侵蚀,地表很难保留较多的枯落物。从表4可以看出,禾本科生物量占比与返青期和主要生长季平均气温具有良好的相关关系,而毒杂草受温度影响则小得多(表4)。在三江源低温严寒环境下,较多的枯落物限制了禾本科(包括莎草科)牧草对光热资源的获取,进一步导致了禾本科牧草生物量占比逐渐减少。另外,禾本科植物多数植株较高,对富含木质素结构成分的氮投资更多,其枯落物质量较差,分解速率慢,导致相对较高的地表遮蔽和较低的土壤养分,不利于植物生长[53]。
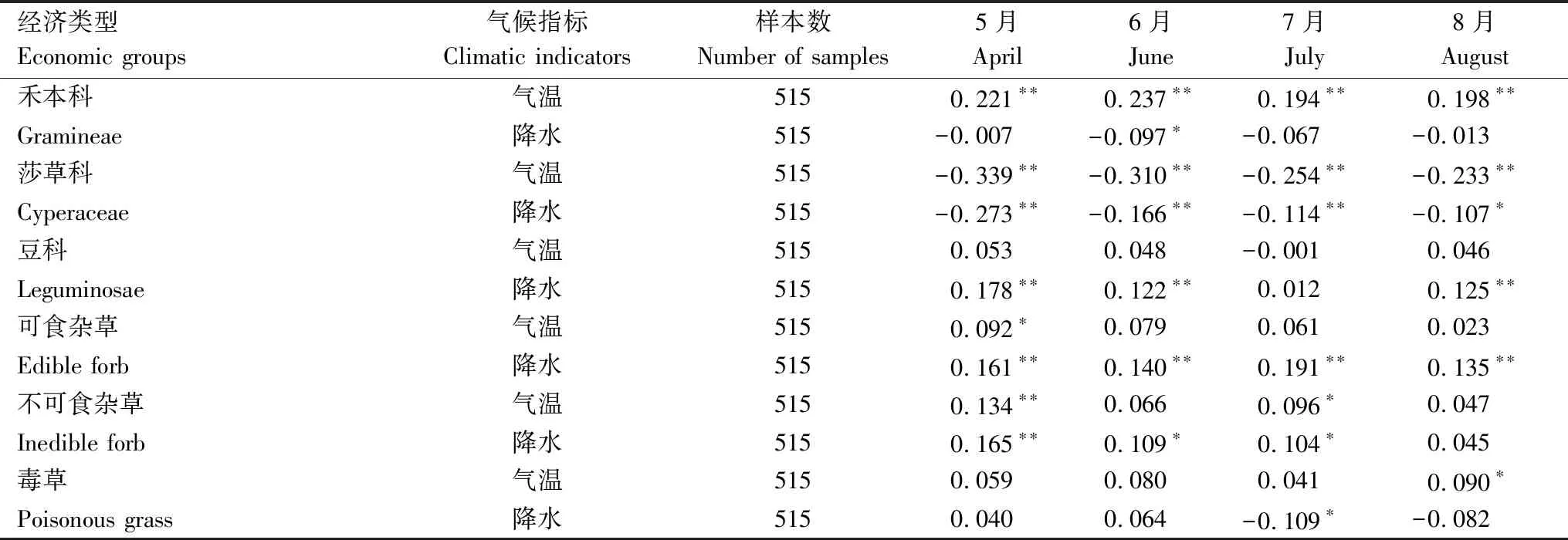
表4 生长季降水与温度与各功能群生物量比例相关系数Table 4 Relationship between temperature,precipitation and biomass proportion in growing season
另外,毒草对禾本科牧草存在化感作用,可通过自身的分泌物或挥发物抑制种子的萌发和胚根的生长来限制禾本科牧草的发育[54]。马瑞君等对黄帚橐吾(Ligulariavirgaurea)的研究发现其体内富含的挥发物会减少禾本科植物种子的萌发率、抑制种子的萌发速率,从而获得竞争优势[55]。唐燕研究表明白苞筋骨草(Ajugalupulina)和铁棒棰(Aconitumpendulum)的水浸提液对当地禾本科牧草具有显著的化感抑制作用[56]。
3.2 放牧区群落结构变化与放牧压力
国家在十二五(2011—2015)和十三五(2016—2020)开始实施“草原生态保护补助奖励政策”,其中对禁牧区域以外的草原根据承载能力核定合理载畜量,实施草畜平衡管理,并按照每年每公顷0.17元的测算标准给予草畜平衡奖励。从监测数据来看,尽管总生物量都没有表现出显著的趋势性变化,但放牧区草地群落结构已发生了明显变化,说明该政策对三江源高寒草甸区群落结构确实产生了深刻的影响。
从围栏封育区来看,禾本科、莎草科生物量占比呈显著减少趋势,毒草和可食杂草呈增加趋势;而在同时段放牧区的群落结构可以看出明显的阶段性特征,禾莎草生物量占比先减少后增加,2005—2011年快速减少,而2012—2017年又呈明显增加趋势(图6)。可食杂草生物量占比在2005—2011年维持较高的比例,而2012—2017则维持较低的比例。毒草生物量占比2005—2011年持续增加,而2012—2017则持续降低。
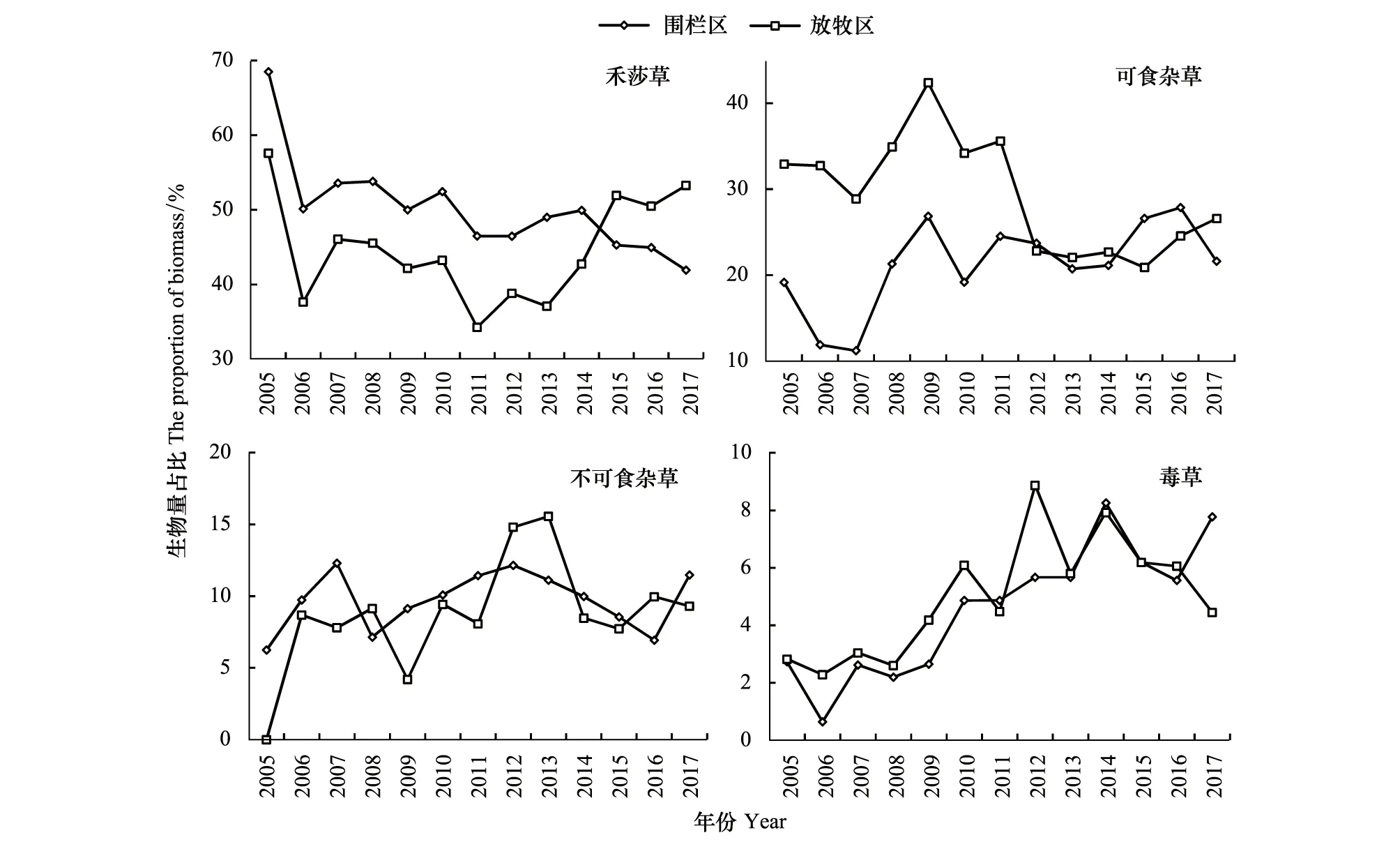
图6 各经济类群生物量占比年际变化Fig.6 Interannual variation of biomass proportion of each functional group
各经济类群生物量占比变化的时间转折点与国家天然草原保护政策实施节点非常吻合。《青海省草原生态保护补助奖励机制实施意见(试行)》青政办[2011]229号明确要求,在2011—2015年对全省1.6千万公顷中度以上退化天然草原实施禁牧,《新一轮草原生态保护补助奖励政策实施方案(2016—2020年)》青政办〔2016〕195号文要求在2016—2020年开始的新一轮草原生态保护补助奖励政策中,继续对这1.6千万公顷天然草原实施禁牧。可见,实施草畜平衡明显改善了草地群落的物种结构。在草畜平衡政策开始的第二年(2012年),草畜平衡在非禁牧区开始全面实施,在减少了牲畜压力后,禾本科与莎草科等优良牧草得到一定程度的修养生息,生物量占比由未实施前的持续减少而改变为持续增加,相应地可食杂草则占比迅速减少,基本稳定在20—30%之间,不可食杂草生物量占比改变了持续增加的趋势,从2014年开始生物量占比也基本稳定在10%以下。
需要注意的是,毒草生物量占比在实施草畜平衡政策后,迅速增加而后逐渐减少。这反映了在实施草畜平衡政策前,在牲畜压力减轻后,这些具有较大叶面积的毒杂草受牲畜踩踏的机会减少,而退化草地又为其扩张提供了生长空间。然而,这些阔叶毒草毕竟耐牧性差,在后续牲畜不断踩踏下植株受损,影响了毒草的生长与繁殖,其生物量占比逐渐减小,可见牲畜的踩踏对控制毒杂草扩张就有重要意义。
3.3 持续围栏封育负面效应与对策
三江源地区目前采取围栏封育的区域并不能显著提高草地的生产力,反而造成了草地群落结构变差等负面效应。未退化状态下作为建群种的禾本科与莎草科牲畜喜食的优良牧草生物量占比持续下降,毒杂草尤其是毒草增加迅速,经过10余年的封育草地群落结构变差且没有出现好转的迹象。草地群落结构变差首先影响的是草地经济价值,禾莎草生物量占比在研究时段内降低了31.2%,即使可食杂草增加了42.0%,但可食草总量总体上降低了13.5%,而且减少的主要是优良牧草。
现有的围栏封育措施还可能带来生态风险。禾本科和莎草科植物一般具有较发达的须根,尤其是莎草科植物根冠比明显高于杂类草[57-58]。禾本科和莎草科生物量占比下降,尤其是莎草科生物量占比下降,意味着高寒草甸区地下根系量减少[59],从而降低了草地生态系统的水源涵养和水土保持功能。另外,禾本科和莎草科凋落物分解速率明显低于杂类草[60],杂类草占比增加也会导致地表凋落物存量减少,进一步削弱了草地水源涵养和水土保持功能。
毒杂草增加可能加速围栏封育区的土壤风蚀。毒杂草植株多高大且比叶面积大,在枯黄期枯落物会快速被秋冬季大风所侵蚀,地表很难保留较多的枯落物,在植被枯萎季节大大加速了土壤风蚀。禾本科与莎草科牧草披针形和针形叶片及以紧伏地表的流线型垫状体,可有效地减少植物体的受风面积[61],从而保留更多的枯落物,大大减少土壤风蚀影响。
围栏封育后植被群落结构改变,可能进一步加剧鼠害,进而加剧草地的退化。在三江源地区主要野生鼠类为喜马拉雅旱獭、高原鼢鼠和高原鼠兔,其食谱狭窄且对鹅绒委陵菜(Potentillaanserina)、甘肃棘豆(Oxytropiskansuensis)、蓝花棘豆(Oxytropiscaerulea)、兰石草(Lanceatibetica)等杂类草有偏好性[62-63]。毒杂草增加为这三类动物都提供了更为丰富的食物来源。而植被生殖枝和营养枝变得低矮,盖度降低,也会为啮齿类动物提供了更好的生存环境[64]。
草地禁牧是青海省草地恢复的主要手段之一。然而,从10余年多样点持续地面观测结果来看,在三江源地区中重度退化草地中长期持续禁牧并不能显著提高草地的生产力,反而会导致禾本科与莎草科牲畜喜食的优良牧草生物量占比持续下降,毒杂草尤其是毒草显著增加,降低草地的经济价值,也会损害水源涵养与水土保持等生态功能。目前,放牧在草地生态恢复中的作用容易被忽视,牲畜践踏与采食可以增加牧草分蘖,加速牧草叶片更新,进而可以提高光合能力。牧草种子在牲畜践踏后更易进入土壤,且牲畜践踏对阔叶杂草的控制,牲畜粪便对土壤氮、磷平衡的维持以及牲畜活动对鼠类的控制等,对草地恢复都具有正面意义。因此,草畜平衡政策可以在不同退化程度草地上实施,在草畜平衡的前提下对各类退化草地减畜可能是目前草地恢复需要特别重视的途径。实际操作中可以基于牲畜可以采食的实际产草量以及生态保护目标,确定更为合理的载畜量,对各类退化草地实施更为精准的草畜平衡管理。
4 结论
围栏封育是三江源生态保护与修复主要措施之一,本文基于2005—2017年多样地草地各经济类群地上生物量的监测数据,对草地群落结构变化进行了分析。研究结果表明研究时段内三江源地区围栏封育显著提高了群落地上生物量,在围栏封育期间地上生物量稳定,但群落结构出现恶化趋势。研究时段内禾本科和莎草科优良牧草生物量占比分别减少了48.2%和23.9%,毒草增加了230.2%,可食杂草增加了42.0%。而放牧区草地群落结构变化具有明显的阶段性特征,禾莎草生物量占比先减少后增加,毒草杂草生物量占比先增加后减少,其变化可能与国家推行的草原生态保护补助奖励政策中草畜平衡措施有关。从现有研究结果来看,大部分研究中围栏区与放牧区的毒杂草比例和生物量无显著性差异,可能与围栏封育的观测年限较短有关。现有的持续围栏封育措施不但降低了草地的经济价值,也带来了一定的生态风险。基于草地实际产草量以及生态保护要求,确定更为合理的载畜量,实施更为精准的草畜平衡管理,应该比切断牲畜与草地关系的长期禁牧封育更为有效。
猜你喜欢
科普童话·百科探秘(2022年9期)2023-01-31 23:54:27
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
今日农业(2021年2期)2021-11-27 19:19:53
绿色科技(2021年10期)2021-06-23 03:13:06
好孩子画报(2019年8期)2019-09-19 12:57:27
草业学报(2019年2期)2019-02-25 01:57:16
电子测试(2017年12期)2017-12-18 06:36:07
湖南城市学院学报(自然科学版)(2016年4期)2016-02-27 14:02:41
农产品市场周刊(2015年39期)2015-10-31 19:12:27
