
不同光质对黄芪抗氧化酶活性及产量和品质的影响
2021-10-12 08:11:06胡瑜辉杨振宇宋诗娟王宝慧石志勇牛景萍梁建萍
山西农业科学 2021年10期
胡瑜辉,杨振宇,宋诗娟,王宝慧,石志勇,牛景萍,梁建萍,柴 智
(1.山西农业大学生命科学学院,山西太谷030801;2.山西中医药大学基础医学院,山西晋中030619)
黄芪是山西省主要道地药材,《中华人民共和国药典》2010版收录药用黄芪为蒙古黄芪(Astragalus membranaceusBunge var.Mongholicus(Bge.)Hsiao)和膜荚黄芪(Astragalus membranaceus(Fisch.)Bge.)的干燥根[1]。黄芪中含有黄芪皂苷、黄酮、多糖、多种氨基酸以及硒、锌等14种人体所需要的微量元素[2],具有补气固表、脱毒生肌之功效,在临床、制药和保健等方面广泛应用。但是近年来,由于环境污染和滥挖滥采的原因,使野生黄芪的产量和分布面积急剧减少[3],人工栽培成为商品黄芪的主要来源。如何提高人工栽培黄芪的品质和产量来代替野生黄芪资源,已经成为学者们研究的重要目标以及任务。药用植物特别是道地药材的质量与环境存在着密切的相关性,其次生代谢产物的积累受到很多环境因素如海拔、光照和水分等的影响,光质对植物的形态建成、生理生化特性及物质代谢都具有一定的影响,光能过量或不足对植物都可能造成胁迫[1]。药用植物栽培地的选择和生态条件的调控已经成为影响栽培药材产量及品质的关键因子[4-5]。
在植物体内光作为能量来源和信号物质,其受体能接收不同波长的光信号,通过信号转导至细胞内,影响次生代谢通路中关键酶基因的表达,进而影响植物的生理过程,最终影响生物量、抗氧化酶活性和药用有效成分的积累[6-8]。随着科学技术研究的不断进步,通过人工调节不同光质以提高经济作物的产量和品质已经成为现实[9]。覆盖不同光质色膜是作为光质调节的一种重要方法,其已应用于植物的设施栽培中,不同波长的光质被植物体内分子吸收后可产生一系列的化学反应,进而调节植株的形态建成、生长发育及生理代谢等[10]。利用不同颜色的滤光膜遮光探明药用植物的生长及其药用有效成分积累的变化,不仅可对道地药材形成的机制和次生代谢途径做出科学分析,而且对生产中提高次生代谢产物含量具有重要的指导意义。
本试验通过在人工栽培黄芪幼苗中设置不同光质色膜,研究光质对黄芪抗氧化系统和有效成分含量的影响,探讨光适应策略,以期找到适于黄芪生长的光生态条件,为定向生产优质药材提供科学依据。
1 材料和方法
1.1 试验地概况
试验在山西农业大学生命科学学院药用植物实践教学与科研基地进行。该地位于山西农业大学校园内,属暖温带季风气候,地处东经112°32′,北纬37°26′,海拔870 m。一年四季分明,年平均气温5~10℃,年均降水量456 mm,无霜期约170 d。
1.2 试验材料
供试材料为1年生蒙古黄芪实生苗,种子采自山西省浑源县官儿乡;光质采用与太阳光透光率相近的不同颜色的滤光膜进行处理,光膜购于上海伟康有色塑料厂。
1.3 试验方法
试验采用单因素4水平随机区组设计,每个处理重复3次。当黄芪生长至60 d后,于2018年8月1日进行黄光(Y)、蓝光(B)、红光(R)3种光质处理,同时以自然光作为对照(CK)。其中,红色光膜波长为620~780 nm,波峰为662 nm;黄色光膜波长为560~590 nm,波峰为570 nm;蓝色光膜波长为400~450 nm,波峰为420 nm。3种光膜透光率均为85%,每组处理30株黄芪。光质处理采用木杆搭建遮光棚进行,搭建高度约100 cm,顶部及四周均用滤光膜覆盖,下部距地面20 cm,以便通风,防止遮光棚内温度过高。光质处理后,于1、30、60、90、120 d(成熟期)采集黄芪叶片,测定其生理指标。待地上部分凋亡后挖出黄芪根,烘干,测定总多糖、总皂苷和总黄酮的含量,并计算产量。
1.4 测定指标及方法
生理指标参考高俊凤[11]的方法进行测定,其中,超氧化物歧化酶(SOD)活性采用氮蓝四唑法测定,过氧化物酶(POD)活性采用愈创木酚法测定,过氧化氢酶(CAT)活性采用紫外吸收法测定,丙二醛(MDA)含量采用硫代巴比妥酸法测定,细胞质膜透性采用电导仪法测定,脯氨酸含量采用茚三酮法测定。
次生代谢物(总多糖、总皂苷和总黄酮)含量测定参照梁建萍等[12]的方法进行,其中,总多糖含量采用蒽酮-硫酸比色法测定,总皂苷含量采用香草醛-硫酸比色法测定,总黄酮含量采用硝酸铝-亚硝酸钠比色法测定。
1.5 数据分析
数据结果采用平均值±标准差表示,采用Microsoft Excel 2010及SPSS 23.0对试验数据进行方差分析及作图。
2 结果与分析
2.1 不同光质处理对黄芪抗氧化酶活性的影响
由图1可知,随着黄芪的生长,自然光(CK)、黄光、蓝光和红光4种光质下SOD活性均呈逐渐下降的趋势;从5个时期来看,黄光下,SOD活性始终显著高于其他3种光质。处理1 d时,黄光、蓝光和红光处理下的SOD活性分别比自然光(CK)条件下高出28.6%(P<0.05)、5.7%和5.9%;处理30、60、90 d后,黄光和红光处理下的SOD活性均显著高于自然光(CK)(P<0.05),黄光处理下的变幅最大,蓝光处理下的变幅最小;处理60 d时,黄光、蓝光和红光下SOD活性分别较自然光(CK)高出42.2%(P<0.05)、8.6%和18.0%(P<0.05);处理120 d时,4种光质对SOD活性的影响同处理1 d时的趋势一致。

从图1可以看出,与自然光(CK)相比,黄光处理可提高POD活性,蓝光处理降低了POD活性,红光处理对POD活性无明显影响。处理30 d时,黄光处理下POD活性高出自然光(CK)18.0%(P<0.05),蓝光处理下低于自然光(CK)22.7%(P<0.05);处理90 d时,黄光处理下POD活性高于自然光(CK)19.1%(P<0.05),蓝光处理下POD活性低于自然光(CK)24.6%(P<0.05)。
从图1可以看出,CAT活性变化对光质的响应与SOD、POD活性都不同,CAT活性受光质影响程度较小,随着黄芪的生长,CAT活性无明显增加或降低的趋势,活性变化趋于平稳。处理1 d时,黄光、蓝光和红光处理下均显著低于自然光(CK)(P<0.05),分别下降了30.3%、20.3%和27.2%;处理30~120 d时,不同光质处理间的CAT活性均无显著差异。
2.2 不同光质处理对黄芪MDA含量、电导率和脯氨酸含量的影响
从图2可以看出,随着黄芪的生长,自然光(CK)、黄光、蓝光和红光4种光质处理下的MDA含量总体呈逐渐上升的趋势,与自然光(CK)相比,黄光处理下的MDA含量略有上升,但二者间差异不显著;蓝光和红光处理下的MDA含量显著降低(P<0.05);处理60 d时,与自然光(CK)相比,蓝光和红光处理下的MDA含量分别下降了66.1%和77.3%,且差异达显著水平(P<0.05),黄光处理下MDA含量升高5.6%,但与CK间差异不显著。
从图2可以看出,自然光(CK)、黄光、蓝光和红光4种光质下黄芪叶片电导率随黄芪生长总体呈缓慢上升的趋势;处理30 d时,与自然光(CK)相比,黄光、蓝光和红光处理下黄芪叶片的电导率分别显著提高了15.8%、12.9%和8.6%(P<0.05);处理30~120 d,电导率受4种光质影响的结果从大到小排序均为黄光>蓝光>红光>自然光。
由图2可知,黄芪叶片脯氨酸含量随时间增加而逐步上升,在5个不同时期,蓝光、黄光和红光处理下的脯氨酸含量均低于自然光(CK),且除处理90 d外,其余处理差异均达到了显著水平(P<0.05);处理60 d时,黄光、蓝光和红光分别较自然光(CK)显著降低10.4%、9.1%和26.5%(P<0.05);处理120 d时,黄光、蓝光和红光处理下的脯氨酸含量分别较自然光(CK)降低9.8%、8.6%和16.2%,且蓝光和红光间差异达显著水平(P<0.05)。由此可见,MDA含量、电导率受短波长光质影响较大,脯氨酸含量受光质胁迫程度较低。

2.3 不同光质对黄芪生长及药效成分积累量的影响
由表1可知,与自然光(CK)相比,株高、根长、茎叶和根的干质量在黄光、蓝红和红光处理下均有所下降。与自然光(CK)相比,株高在黄光、蓝光和红光处理下分别显著下降了17.4%、39.8%和12.1%(P<0.05);根长在黄光、蓝光和红光处理下分别显著下降了16.9%、35.8%和16.9%(P<0.05);茎叶干质量在黄光、蓝光和红光处理下分别显著下降了1.8%、22.2%和10.8%(P<0.05);根干质量在黄光、蓝光和红光处理下分别显著下降了2.0%、12.9%和11.6%(P<0.05)。可见,3种光质均明显降低了黄芪的纵向生长和干物质的积累。

表1 不同光质对黄芪单株生物量的影响
从表2可以看出,与自然光(CK)相比,不同光质处理均提高了多糖、黄酮和皂苷这3种药效成分的含量和单株产量,并且蓝光处理高于红光和黄光处理。与自然光(CK)相比,多糖含量在蓝光、黄光和红光处理下分别显著增加了78.4%、52.8%和27.5%(P<0.05);多糖单株产量在蓝光、黄光和红光处理下分别提高了55.4%、49.8%和12.7%,且蓝光、黄光处理均显著高于红光处理(P<0.05)。与自然光(CK)相比,皂苷含量在蓝光、黄光和红光处理下分别增加了133.3%、80.0%和13.3%,且蓝光、黄光处理与红光处理间差异达显著水平(P<0.05);皂苷单株产量在蓝光、黄光处理下分别显著提高了97.2%、70.8%(P<0.05),而红光处理下降了3.8%,但与CK间差异不显著。与自然光(CK)相比,黄酮含量在蓝光和黄光处理下分别显著增加了80.5%和51.2%(P<0.05),而红光处理下降低了9.8%,但与CK间差异不显著;黄酮单株产量在蓝光和黄光处理下分别显著提高了57.6%和48.4%(P<0.05),红光处理下显著降低了20.2%(P<0.05)。
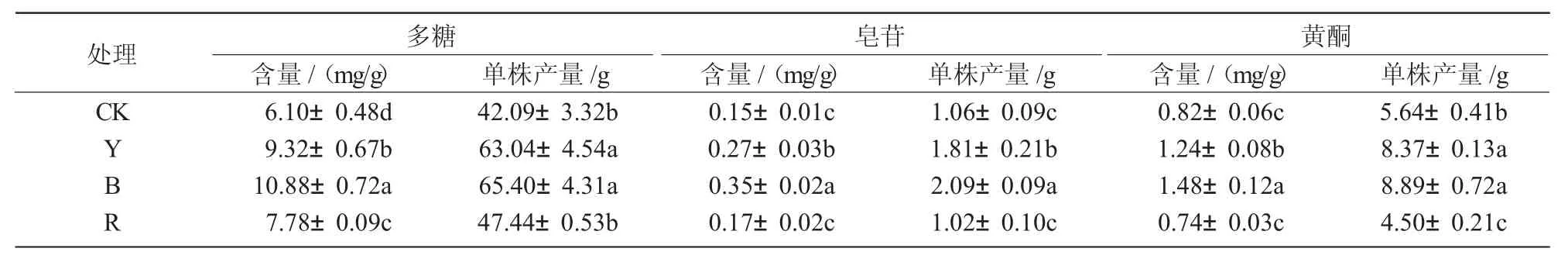
表2 不同光质对黄芪有效成分含量和单株产量的影响
3 讨论
3.1 光质处理对抗氧化酶系统的影响
植物从种子萌发到生长发育都受到光照的调控,而光质可以调控植物碳水化合物的代谢,并与作物的产量和品质相关[13]。近年来,国内外学者以光质作为环境调控因子,探讨其对植物抗氧化系统及生长的影响[14]。SOD是植物抗氧化系统中免受活性氧伤害的第一道防线,与POD、CAT协调配合清除过氧化物自由基,以保证细胞的正常功能[15]。研究表明,蓝光会使不同植物抗氧化酶的活性升高,以保持较低水平的超氧阴离子、过氧化氢和MDA[16-17]。红光处理的SOD、CAT活性以及MDA含量和H2O2含量在先锋橙和红桔幼苗中均高于蓝光处理[18]。本试验研究发现,与自然光相比,蓝光下SOD、POD活性无显著变化,而黄光下SOD、POD活性均上升,CAT受不同光质的影响较小,这与李倩[16]的研究结果一致。说明蓝光对黄芪幼苗的氧化胁迫伤害程度较轻,可以通过体内SOD、POD、CAT这3种抗氧化酶的协调作用使活性氧自由基达到平衡。
细胞膜透性的变化可以反映植物细胞对外部环境的敏感程度。MDA是膜脂过氧化的重要产物之一,用来表示细胞膜透性大小,从而间接衡量膜系统受损程度以及植物的环境适应性。本研究发现,与自然光相比,蓝光和红光处理下MDA含量下降,而黄光处理下MDA含量上升。电导率是反映细胞膜受伤害的另一个常用指标。本研究中,在黄芪生长过程中,4种不同光质对电导率影响大小均表现为黄光>蓝光>红光>自然光,且都存在显著差异。以上结果表明,蓝光处理对黄芪没有造成膜脂过氧化的发生,有利于黄芪的正常生长。
在正常环境条件下生长的植物,体内游离脯氨酸含量较低,但在逆境(如干旱、寒害和热害等)胁迫条件下,植物体内游离脯氨酸含量会显著增加。本研究中,与自然光(CK)相比,蓝光处理下的脯氨酸含量升高,这与王丽伟等[19]研究发现的“番茄幼苗在蓝光下脯氨酸含量较高”的结论相一致。表明在蓝光处理下,黄芪更易合成较多的脯氨酸含量来调节细胞的渗透压,从而使细胞质膜保持完整性,有助于黄芪的生长发育。
3.2 光质处理对黄芪生物量和有效成分积累的影响
光对植物生长发育和品质的影响已逐渐成为目前研究的热点[20],光质对植物生长发育的影响首先表现在植物光形态建成方面,株高、根长、叶面积、干物质积累等生物量是反映植物长势状况的重要指标,不同光质对生物量有着不同程度的影响。本研究中,在黄光、蓝光和红光处理下,黄芪的株高、根长和干质量受到不同程度的抑制,这与黄枝等[21]、阎秀峰等[22]和陈骊君等[23]研究得出“不同光质处理下植株的生长及鲜质量、干质量有不同程度下降”的结果相一致。
近年,植物次生代谢合成受光质的调控也受到学者的广泛关注,特别是药用植物研究领域。光质通过调控植物基因表达来促进或抑制一些功能性蛋白或酶的合成,最终影响植物的次生代谢及形态发生[24]。本试验研究表明,4种光质对黄芪根部有效成分积累的影响从大到小排序均表现为蓝光>黄光>红光>自然光,光质都不同程度地促进了黄芪根部有效成分的积累。有研究表明,蓝光也能显著增加植物内花青素[25]的含量,对植物体内其他营养物质(如槲皮素、异鼠李素)的积累也有一定的影响[26]。李小明等[13]研究发现,黄光、蓝光和红光3种光质下淫羊藿苷类黄酮含量均高于白光,而以黄光最高,这与本试验研究结果有所差异。这可能是由于不同植物对光质的耐受性不同造成的。
4 结论
本研究结果表明,蓝光能有效维持黄芪体内抗氧化系统平衡,有效调控黄芪次生代谢物合成,提高总黄酮、总多糖和总皂苷的含量,从而改善黄芪药材的品质,提高药材产量。这一结果可为黄芪人工定向栽培中光质的选择提供基础理论依据和技术支撑。
猜你喜欢
学与玩(2022年6期)2022-10-28 09:18:52
四川林业科技(2022年1期)2022-03-22 07:48:32
上海农业学报(2022年1期)2022-03-08 00:09:12
Plasma Science and Technology(2021年8期)2021-08-05 08:29:44
——记嘉荫县红光乡燎原村党支部
奋斗(2020年24期)2021-01-16 06:23:36
Plasma Science and Technology(2020年8期)2020-08-26 04:58:04
浙江农业学报(2017年1期)2017-05-17 06:13:36
作文与考试·小学低年级版(2016年10期)2016-09-10 07:22:44
中国果业信息(2015年12期)2015-01-24 07:31:03
无机化学学报(2014年1期)2014-02-28 17:30:11
