
吉林省蛟河市针阔混交林林层结构与地形的关系
2021-10-09 01:52鲁君悦张春雨赵秀海
生态学报 2021年16期
鲁君悦,张春雨,赵秀海
北京林业大学国家林业和草原局森林经营工程技术研究中心,北京 100083
森林结构是森林生态学研究中的基础内容,对揭示群落演替动态和物种多样性维持机制具有重要意义[1]。物种组成、密度结构等森林结构指标是森林可持续经营管理中必须测定和调控的内容[2- 3]。森林群落的结构特征不仅表现在水平方向上,也表现在垂直方向[4]。对森林群落垂直结构进行优化有利于其充分利用环境资源,提高资源的高效配置,从而提高整个森林群落的生产力[5- 6]。相比林下层,林冠层是森林同外部环境相互作用较活跃和直接的界面。林冠层对光照和土壤等一些资源的主导作用会对林下层林木的生存条件以及物种组成和生产力产生重要影响[7]。在天然林中,林冠层对森林生物量的贡献远超于林下层,而林下层对生物多样性的贡献较大[8]。
近年来,特别是巴拿马巴洛科罗拉多岛(BCI)在热带雨林建立首个50 hm2森林动态监测大样地后[9],许多学者借助森林动态监测样地开展了大量关于森林群落结构的研究[10-12],对群落动态的长期研究、物种多样性维持机制和物种空间分布格局等作出了巨大贡献[13- 14],推动了大样地在全球范围内建立[15]。21世纪初,在中国科学院森林生物多样性委员会的组织下多家单位建立了长期固定监测样地,推动了中国森林生物多样性监测网络的形成[16],进而为探索我国森林群落结构、物种共存和物种多样性维持机制等生态学机理提供数据支撑[17]。
森林群落的结构特征受海拔、坡向和坡度等地形因子的影响[18],地形异质性在一定程度上会改变环境条件,如土壤厚度和土壤养分分布,同时也会影响光照和降水的空间再分配。近年来,群落水平上的植物分布受地形因子的影响已受到广泛关注,但目前大部分研究多集中在群落水平上的结构特征与地形因子的关系[19-20],而忽略了地形因子对不同林层群落结构动态变化的影响。
东北林区在改善气候、防风固沙和生物多样性保护方面都发挥着重要作用[21-22],针阔混交林是东北地区重要的森林类型之一[23]。近年来,大部分研究只关注了群落水平上的森林结构特征及动态变化[24-25]。因此,深入了解温带针阔混交林不同林层结构特征及其动态变化具有重要意义。本文以吉林蛟河30 hm2的针阔混交林固定监测样地为基础,利用2010和2015年的野外调查数据,探讨了不同林层群落结构动态变化及其与地形因子之间的关系,旨在进一步阐明温带针阔混交林的演替规律,以期为针阔混交林的结构优化和可持续经营管理提供理论支持。
1 研究区概况与研究方法
1.1 研究区概况
研究地点在我国吉林省蛟河市林业实验区管理局林场内,属于长白山系张广才岭山脉。该区域为受季风影响的温带大陆性气候,年均降水量在700—800 mm之间,多集中于6—8月,年平均气温3.8℃,最冷月为1月,平均气温为-18.6℃,最热月为7月,平均气温为21.7℃。研究样地地势起伏较大,地面凹凸不平,海拔在577.84—780.67 m之间。土壤类型为暗棕色森林土,富含有机质,土层厚为20—100 cm。研究区植被类型属于长白山植物区系,主要的乔木树种包括红松(Pinuskoraiensis)、裂叶榆(Ulmuslaciniata)、紫椴(Tiliaamurensis)、白牛槭(Acermandshuricum)、枫桦(Betulacostata)、色木槭(AcermonoMaxim)、胡桃楸(Juglansmandshurica)。主要灌木树种包括毛榛(Corylusmandshurica)、东北鼠李(Rhamnusdavurica)、簇毛槭(Acerbarbinerve)、暴马丁香(Syringareticulata)等。
1.2 样地设置与数据获取
参照CTFS(Center for Tropical Forest Science)样地建设标准和操作规范,于2010年夏季,在吉林省蛟河市林业实验区管理局的林场内建立了500 m×600 m的固定样地。利用全站仪将样地划分为750个20 m×20 m的连续样方,在20 m处结点位置用水泥桩固定,水泥桩用喷漆喷上样方编号。同年对样地内所有胸径大于等于1cm的木本植物进行初测,除一些倒木和枯立木无法定名外,记录个体的种名、胸径、树高、冠幅(东西、南北)、枝下高及相对位置等信息,同时挂牌标记。2015年对样地进行复测调查。
地形因子选择海拔、坡向、坡度和凹凸度这4个变量,测量海拔因子时,使用GPS仪器测量20m×20m小样方内4个角位置上的海拔,该样方的海拔高度为这4个角上的海拔平均值。该样方的坡向值为垂直平面与样方内各顶角组成平面的夹角度数的平均值。坡度的计算方法与坡向的大致相同,在小样方内通过四个顶角的其中任意三个角的顶点组成的平面与水平面所产生的夹角的平均值为该样方的坡度。凹凸度的计算方法分为两种,第一种处于样地边缘的样方计算方法为样方中心的海拔高度值减去四个角位置的海拔平均值,第二种样地内的样方计算方法为样方中心的海拔高度值减去与这个小样方相邻的所有样方的海拔平均值。
1.3 林层划分和生产力计算
林层分层研究包括定性、定量和遥感等研究方法,本文依据林层定性划分方法将森林划分为林冠层和林下层两部分[26],树高占总体前30%归为林冠层,其余个体归于林下层。本文以样地内胸径大于等于1 cm的活立木为研究对象,在分析数据之前对数据进行处理,剔除存在异常数据的树木。根据吉林省不同区域各物种的胸径树高公式和立木材积公式[27],求得不同林层5年内的年均蓄积增长量作为各林层的蓄积生产力。
1.4 森林结构指标计算
1.4.1物种组成及特征
物种多样性采用物种香农指数(HS)作为代表,物种香农指数可以同时反应群落的物种分布的均匀程度和丰富度。

式中,NS是样方内物种总数;N是样方内总个体数;ni是第i个物种的个体数。
重要值(importance value,IV)是以综合数值衡量植物物种在群落中的相对重要值,用重要值来表征树种优势度。计算公式如下:

1.4.2林分密度结构
用树木个体胸高断面积之和与数量表示林分密度和株树密度。采用胸径变异系数(VarD)和树高变异系数(VarH)这2个指标代表林分结构多样性。
式中,N是样方内总个体数;DBHk是样方内第k个树木个体的胸径值;μ是样方中所有树木个体的胸径平均值。Hk是样方内第k个树木个体的树高值;μh是样方中所有树木个体的树高平均值。
1.5 数据处理
常见的相关分析方法包括简单相关分析、偏相关分析和距离分析。本文使用的相关分析方法为偏相关分析法,偏相关分析法也称为净相关分析法,当想要分析两个变量之间的关系时,通过排除其他相关联的变量的影响后,再对这两个变量的相关性进行分析。与简单的相关分析相比,偏相关分析能更准确的反映变量间最本质的联系。
所有数据在R 3.5.1软件和SPSS软件中进行统计与分析。
2 结果与分析
2.1 不同林层群落结构动态变化
基于2015年750个小样方的计算结果表明(表1),林冠层胸径范围较大,林下层胸径范围较小,林冠层的胸径平均值为林下层的6倍。林下层物种丰富度略微高于林冠层。林冠层是整个森林群落蓄积生产力的主要贡献者,林下层蓄积生产力约为林冠层的1/10。林冠层林分密度远远高于林下层,而林下层的株树密度约为林冠层株树密度的2倍。2010—2015年林冠层胸径变异系数、树高变异系数和物种香农指数变化很小。相比林冠层,林下层的三个指标稍有变化,呈增大趋势(图1)。
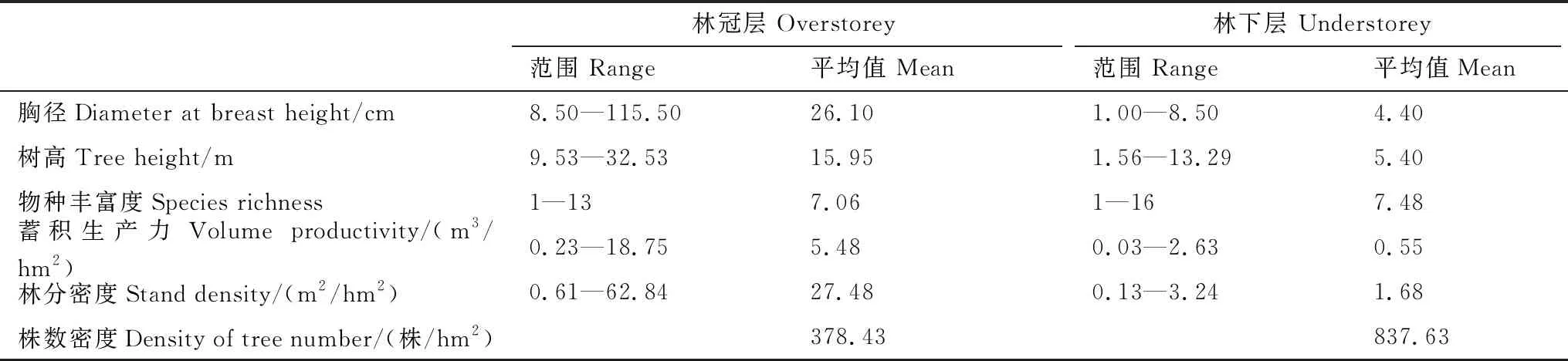
表1 样地内基础变量计算结果
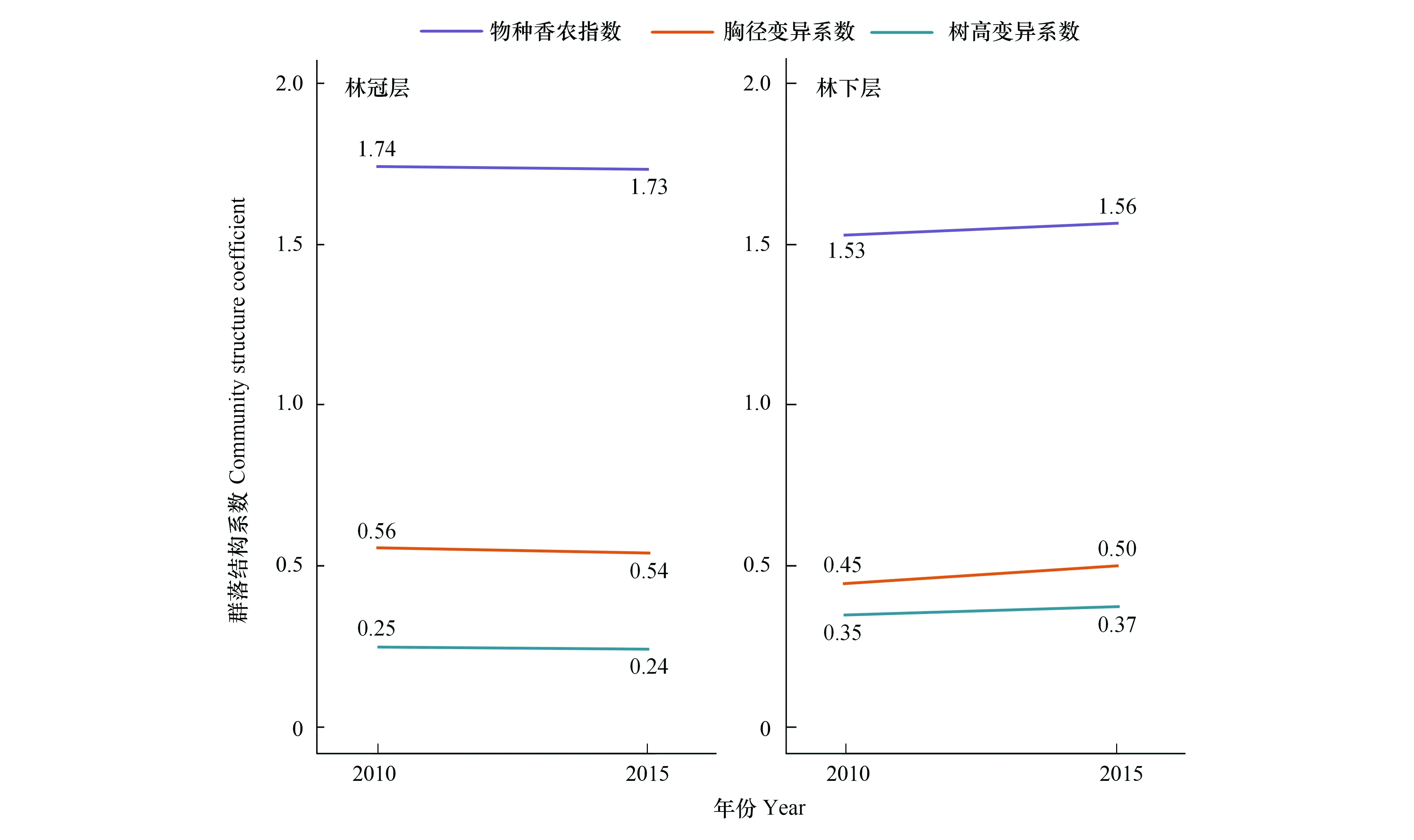
图1 不同林层物种多样性和林分结构多样性动态Fig.1 Dynamics of species diversity and stand structure diversity across tree strata
2010和2015年的群落调查发现林冠层树木依然保持为17科24属37种,林下层树木由18科27属42种增至18科27属43种。在这两次调查期间,仅有1个偶见种(总株数≤3)出现变动,其中臭松(Abiesnephrolepis)进入群落。2010—2015年,林冠层重要值排在前10位的优势物种组成和树种排列次序都没有发生变化,优势物种在林冠层的优势度稍微扩大,累计重要值由80.72增至81.02(表2)。林下层优势树种的累计重要值由83.66降至83.53(表3)。在林冠层重要值排在前10位的优势物种的胸高断面积值均提高,其中紫椴、裂叶榆、胡桃楸、色木槭、枫桦和红松这6个树种胸高断面积增长量之和占总断面积增长量的66.3%,但其多度和频度均下降(表2)。林下层优势物种的胸高断面积值大部分略微增加。林冠层树木个体数量总体上减少175株,林下层树木个体数量总体上增加3503株。
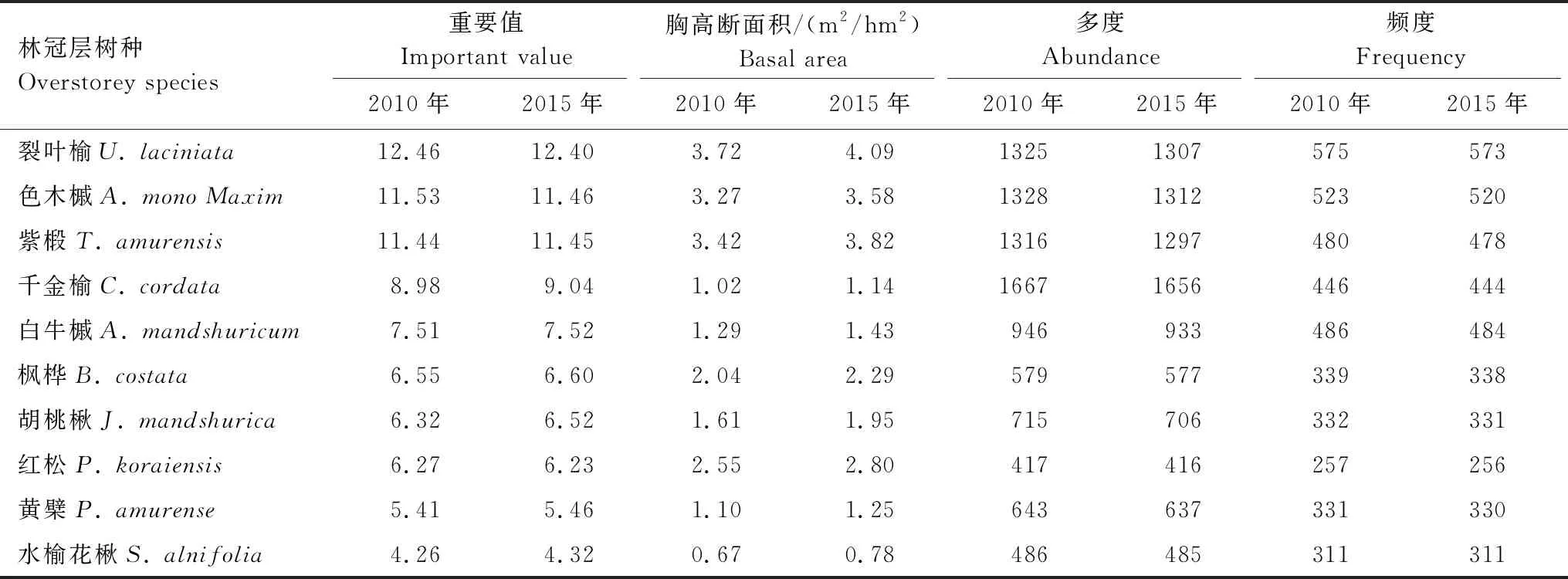
表2 2010年和2015年林冠层优势树种主要指标统计

表3 2010年和2015年林下层优势树种主要指标统计
2.2 不同林层群落结构变化与地形因子关系
林冠层物种多样性变化与海拔和坡向这两个地形因子的相关性较大(图2),与海拔的偏相关系数为0.082(P<0.05),与坡向的偏相关系数为0.093(P<0.05),与凹凸度和坡度无显著相关性。林下层物种多样性变化与海拔、坡向、凹凸度和坡度均无显著相关性。
不同林层林分结构多样性变化均与海拔呈负相关关系。林冠层胸径变异系数变化与海拔和坡向这两个地形因子呈显著负相关关系(-0.118和-0.093)。林冠层树高变异系数变化与海拔和凹凸度这两个地形因子呈相关关系(-0.190和0.083)。林下层胸径变异系数变化和树高变异系数变化与海拔的偏相关系数分别为-0.111(P<0.001)和-0.147(P<0.001)。不同林层林分结构多样性变化和物种多样性变化均为显著正相关。

图2 不同林层物种群落结构及地形因子的偏相关系数图Fig.2 Partial correlation among species diversity, structure diversity and terrain factor across tree strata SSI:物种香农指数;cvDBH:胸径变异系数;cvH:树高变异系数;Ele:海拔;Asp:坡向;Con:凹凸度;Slo:坡度
3 讨论
3.1 不同林层结构动态变化
森林群落结构是了解森林生态系统功能形成和维持机制的重要基础[28],以往的研究大都只关注整个群落水平上群落结构特征及其变化[10,24-25],而忽略了不同林层之间的差异,本研究重点关注了不同林层群落结构特征及其动态。林下层的物种丰富度高于林冠层,这是因为除了少数仅出现在林冠层的物种如朝鲜柳、拧筋槭、山杨和香杨外,林下层还有林冠层树木的幼树和幼苗存在。此外,群落内还有一些仅存在于林下层的树种,如东北山梅花、金银忍冬、东北鼠李、卫矛和山葡萄等。这与大部分人的研究一致,随着林层向上,物种丰富度指数下降[8,29]。簇毛槭、稠李、接骨木和长白忍冬等非林冠层优势种在林冠层消失或少量存在,仅紫椴、枫华和胡桃楸等少数物种成为林冠层的优势种,这可能是由于环境资源的异质性和物种本身的特性所造成的[30]。大部分研究发现,在天然的森林群落中生产力或生物量主要是由林冠层树种所贡献,而林下层树种对物种丰富大贡献较大[8,31],这与本文的研究结果一致。
林冠层林分结构多样性的指标减小,造成这一现象的原因可能是因为在林冠层树木株数减少较多,从而使林冠层产生林隙,因此林下层群落能进行更多的更新和演替,产生更多优势树种的幼树[32]。在林下层林分结构的分化程度增加,可能是因为林分结构越复杂,群落就会产生更多异质性的光环境,使得耐阴树种的存活率变高[33]。不同林层物种丰富度变化不大,这可能是因为本研究样地处于老龄林发展阶段,群落稳定性较高。
2010年到2015年,物种组成增加1种,偶见种引起了物种丰富度的变化,体现了偶见种在维持物种多样性中的重要性[34]。林冠层优势树种的重要值在群落演替过程中逐渐扩大,以此来确定在群落中的优势地位,这将会导致一些竞争力弱的物种被淘汰[35]。在这5年内,林冠层的优势树种的相对重要性没有发生改变。其他林冠层树种例如香杨、山杨和白桦等先锋物种的重要值降低,且香杨和山杨仅存活于林冠层,这可能是由于树种的竞争和庇护作用导致林下层已经没有其更新的幼树和幼苗[36],林冠层的紫椴、红松和色木槭等阴性树种在林下层更新良好。在长白山对阔叶红松林的研究发现群落演替实质是紫椴、红松和色木槭等阴性树种取代黄檗、山杨和白桦等阳性或半阴性树种的过程[37],这与本研究的结果基本一致。
3.2 地形因子对不同林层群落结构变化的影响
地形是影响物种变化和物种丰富度的主要环境因子之一,地形因子中的海拔、坡向、坡度等因子的改变将会引起土壤养分、土壤水分、光照、湿度和温度等环境因子的变化[38]。以往的研究大多只考虑了整个群落水平上的森林结构特征和地形因子之间的关系[19-20],本研究利用偏相关分析法考虑了不同林层群落结构动态变化与地形因子之间的关系,结果表明在林冠层物种多样性变化与海拔和坡向呈正相关关系,在林下层物种多样性变化和四个地形因子无显著关系,在不同林层林分结构多样性变化和海拔均呈负相关关系。但是王志恒等人的研究发现物种丰富度随着海拔的升高呈现先升高后下降的趋势[39],出现不同结果的原因可能是研究地整体海拔不高,处于升高的前半部分,未到达下降的临界海拔,或是林分年龄存在差异。
在林冠层物种多样性变化和海拔呈正相关关系可能是因为研究样地在低海拔的山谷处有一条溪流,由于河流的冲洗作用使得土壤厚度变低,且河道旁的石块非常多,所以植株在低海拔地区难以生长,随着由低海拔山谷向高海拔山脊过渡时,土壤厚度增加使得物种多样性增强[40],且林冠层树种在山脊处可以获得更多光照,造成的生态位差异会使物种更有效的利用资源[5],进而增加物种多样性。这和大多数人的研究结果一致,随着海拔升高物种多样性增加[40-41]。在林下层,地形因子对物种多样性变化的影响不显著,可能是因为林冠层树种对资源的主导资源会削弱林下层树木的资源利用率[42-43],使得两者间不存在显著关系。
在林冠层和林下层林分结构多样性均随着海拔升高而减少,可能是因为随着海拔的升高植物可以接受的光照资源变多,林冠层的优势树种相比劣势树种能更有效的利用这些资源,造成林冠层的劣势树种获取的资源有限而生长缓慢[44-45],使得林冠层大部分树木的胸径和树高变化较小,从而随着海拔升高林冠层林分结构多样性变化减少。而由于林下层树木随着海拔升高死亡个体增加,林分结构多样性减少。
4 结论
本研究以吉林省蛟河市针阔混交林为研究对象,分析了不同林层群落结构动态变化及其与地形因子的关系。不同林层的群落结构特征存在较大差异,这主要是由于环境资源的异质性和物种本身的生理特性所引起的。林冠层优势树种重要值的扩大将会导致一些竞争力弱的物种被淘汰。地形因子中的海拔是影响群落结构动态变化最重要的因子。现阶段的研究只分析了不同林层结构动态变化及其与地形因子的关系,并未深入阐释其它环境因子如土壤营养状况、水分、光照与不同林层结构动态变化的关系,这需要进行更深入的研究,以便更好的理解森林的结构特征和变化规律,为保护利用天然林资源提供更加科学的理论依据。
猜你喜欢
贵州畜牧兽医(2022年3期)2022-06-28
现代园艺(2021年23期)2021-12-01
新农业(2020年18期)2021-01-07
花卉(2020年24期)2021-01-04
环保科技(2020年2期)2020-05-21
浙江农林大学学报(2019年6期)2019-11-13
装备环境工程(2019年9期)2019-10-24
读者·校园版(2019年3期)2019-01-28
少年文艺·开心阅读作文(2017年1期)2017-02-24
小小说月刊·下半月(2015年11期)2015-05-14
